










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 no.3 Texcoco abr./may. 2018
https://doi.org/10.29312/remexca.v9i3.1225
Investigation note
Nutritional growth and extraction of poinsettia in response to the ratio nitrate:calcium and phenological stage
1Facultad de Ciencias Agropecuarias-Universidad Autónoma del Estado de Morelos. Av. Universidad 1001, CP. 62209. Col. Chamilpa, Cuernavaca, Morelos. (torres27-tov@yahoo.com.mx; ijac96@yahoo.com.mx; mjsainz63@yahoo.es).
2Departamento de Horticultura-Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1923, Buenavista, Saltillo, Coahuila, México. (luisalonso.valdez@uaaan.mx; hernandez865@hotmail.com).
The nutrient requirements of the plants are different in their different phenological stages, so in the present experiment the Steiner solution was modified to adapt it to each stage of poinsettia (root growth, vegetative development and pigmentation). The Prestige cultivar was used the substrate was a mixture of leaf soil, coconut fiber and red tezontle in proportion 60:20:20 (%, v/v) and black polyethylene container of 15.24 cm in diameter. The experiment was carried out in the Faculty of Agricultural Sciences of the Autonomous University of the State of Morelos, Cuernavaca, Morelos. Morphological variables of the poinsettia plants were evaluated in response to the relative concentration between NO3 - and Ca2+ of the nutrient solution. The supply of nutrients according to the phenological stages of poinsettia affects the growth, extraction and concentration of nutrients, but depends on the nutritional regime. The regimen with the concentration (in meq L-1) NO3 -:Ca2+ 10:7 in radical growth, 10:9 in vegetative development and 12:9 in the pigmentation stage induced the poinsettias to present greater extraction of N, P, K, Ca and Mg, will increase the concentration of K and the morphological characteristics and the production of dry matter were outstanding.
Keywords: electrical conductivity; nutrients; nutritive solution; phenology
Los requerimientos nutrimentales de las plantas son distintos en sus diferentes etapas fenológicas por lo que en el presente experimento se modificó la solución de Steiner para adaptarla a cada etapa de nochebuena (crecimiento radical, desarrollo vegetativo y pigmentación). Se utilizó el cultivar Prestige, el sustrato fue una mezcla de tierra de hoja, fibra de coco y tezontle rojo en proporción 60:20:20 (%, v/v) y contenedor de polietileno negro de 15.24 cm de diámetro. El experimento se llevó a cabo en la Facultad de Ciencias Agropecuarias de la Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos. Se evaluaron variables morfológicas de las plantas de nochebuena como respuesta a la concentración relativa entre NO3 - y Ca2+ de la solución nutritiva. El suministro de nutrimentos de acuerdo con las etapas fenológicas de la nochebuena afecta el crecimiento, extracción y concentración de nutrimentos, pero depende del régimen nutrimental. El régimen con la concentración (meq L-1) NO3 -:Ca2+ 10:7 en crecimiento radical, 10:9 en desarrollo vegetativo y 12:9 en la etapa de pigmentación indujeron que las plantas de nochebuena presentaran mayor extracción de N, P, K, Ca y Mg, se incrementara la concentración de K y las características morfológicas y la producción de materia seca fueran sobresalientes.
Palabras clave: conductividad eléctrica; fenología; nutrimentos; solución nutritiva
The detailed studies of the growth of the plants allow quantifying different aspects such as the duration of the cycle, definition of the stages of development and distribution of photoassimilates between organs. Growth analyzes are basic to better understand the physiological processes that determine plant production, and thus rationally underpin crop management practices such as nutrition, irrigation, pruning, protection strategies, among others (Azofeifa and Moreira, 2004).
The ions of the nutritious solution (SN) establish mutual relationships that are important to consider to cover the nutritional needs of each stage of crop development (Gómez and Montoya, 2001). For this reason, several investigations are directed to the fractional application of nutrients (Klocke et al., 1999) according to the requirements of the crop (Andraski et al., 2000) in order to reduce the risk of contamination and obtain yields competitive
The poinsettia has high demand for nitrogen (N) and potassium (K) (Martínez, 1995; Ayala-Arreola et al., 2008), as well as an unusually high requirement for calcium (Ca), magnesium (Mg) and molybdenum (Dole and Wilkins, 2005; Ayala-Arreola et al., 2008). The excess of N induces exuberant and succulent foliage, while the deficiency causes weak stems. Vázquez and Salome (2004) recommend making complementary applications of Ca through irrigation or foliage between the second and third stage in order to obtain stems more resistant to tearing (avoid fragile stems) and bracts of higher quality.
With the foundation that plants have a different requirement of nutrients in their different phenological stages, the present study aimed to determine the morphological response and nutritional extraction of poinsettia to the modification of the SN in its three phenological stages.
The experiment was carried out in a greenhouse with plastic cover of the Faculty of Agricultural Sciences of the Autonomous University of the State of Morelos, Chamilpa campus (18º 58’ 52.87” north latitude and 99º 13’ 57.92” west longitude, 1 875 meters above sea level) in Cuernavaca, Morelos, with minimum and maximum average temperature of 13.7 and 34.7 °C respectively, relative humidity between 55% and 73%. The photosynthetically active diurnal radiation was on average 203 μmol m-2 s-1. It used Prestige cultivar poinsettias transplanted on July 6 in black polyethylene containers of 15.24 cm in diameter, in a substrate made with sifted leaf soil, coconut fiber and red tezontle (granulometry between 0.1 to 0.5 cm in diameter) in proportion of 60:20:20 (%, v/v). The irrigation was performed with SN of electrical conductivity (CE) of 2 dS m-1 and pH 5.5.
The treatments were generated by the combination of the NO3 - and Ca2+ concentrations of the nutrient solution according to the phenological stage of poinsettia (Table 1), obtained by Torres-Olivar et al. (2015). The modification between NO3 -:Ca2+ was made based on the universal nutrient solution (Steiner, 1984), keeping constant the total concentration of anions and cations in 20 meq L-1 each, as well as the mutual relations between SO4 2- : H2PO4 - of 7 and K+: Mg2+ of 1.75. The relative concentration of the ions involved remained within the limits of a true solution because it is known that the interaction between them influences the absorption, distribution or function of some other nutrient in the plant, which reduces the probability to induce deficiencies or toxicities (Schwarz, 1995; Villegas et al., 2005).
The control treatments were two: 1) universal nutritive solution (Steiner, 1984, with 12 of NO3 - and 9 of Ca2+, in meq L-1) supplied in the three phenological stages; and 2) the one recommended by Martinez (2011) which consists in applying the universal nutrient solution (Steiner, 1984) in 80% (NO3 -, 9.6; Ca2+, 7.2, in meq L-1) of the original concentration in radical growth, 120% (NO3 -, 14.4; Ca2+, 10.8, in meq L-1) in vegetative development and 80% (NO3 -, 9.6; Ca2+, 7.2, in meq L-1) in the pigmentation stage (Table 1).
Table 1 Nutritional regimens evaluated on poinsettia.
| Regimen | Radical growth | Vegetative development | Pigmentation |
| SM/SM/SM | 10:7 | 10:9 | 10:9 |
| SM/SM/ST | 10:7 | 10:9 | Steiner |
| SM/ST/SM | 10:7 | Steiner | 10:9 |
| SM/ST/ST | 10:7 | Steiner | Steiner |
| ST/SM/SM | Steiner | 10:9 | 10:9 |
| ST/SM/ST | Steiner | 10:9 | Steiner |
| ST/ST/SM | Steiner | Steiner | 10:9 |
| STEINER | Steiner | Steiner | Steiner |
| 80/120/80 | 80% | 120% | 80% |
10:7= 10 meq L-1 of NO3 - and 7 meq L-1 of Ca2+; 10:9= 10 meq L-1 of NO3 - and 9 meq L-1 of Ca2+; SM= modified solution. ST= universal nutrient solution (Steiner, 1984) containing 12 meq L-1 of NO3 - and 9 meq L-1 of Ca2+.
The root growth was considered from the transplant to the pruning of the plants, which was done when the root was visible in the periphery of the root ball. The vegetative development, from the pruning to the appearance of the transition bracts, and the pigmentation ranged from the appearance of the transition bracts to the presence of pollen.
The evaluated growth variables included plant height measured from the neck to the apex of the highest shoot, the diameter of the plant was determined with a digital vernier, the leaf area and the bracts (LI-3100C, LI-COR, Inc. Lincoln, Nebraska, USA) and the total dry matter weight, which was recorded after drying the tissues in an oven with forced air circulation at a temperature of 70 °C for 72 h. To determine the extraction of nutrients and relate it to the phenological stage, at the end of the experiment the destructive sampling of the plants of four selected treatments was carried out (SM/SM/SM, SM/SM/ST, 80/120/80 and Steiner).
The total N concentration was determined by the micro-Kjeldahl method (Chapman and Pratt, 1973, Brearen and Mulvaney, 1982) and that of K with a Corning 400 Flamemeter (Alcantar and Sandoval, 1999). Phosphorus (P), Ca and Mg were determined by inductively coupled plasma emission spectrometry (ICP-AES VARIAN, Liberty model). The data obtained were analyzed in SAS 8.1 (SAS Institute, Cary, North Carolina, USA) in a randomized complete block design with four repetitions; each repetition was a container with a plant. The variables with significant difference were subjected to Duncan’s multiple comparison test of means (p≤ 0.05).
The modification of the SN in each of the phenological stages affected the growth of the poinsettia plant in a different way. The supply of nutrients through the SM/SM/ST regime significantly favored the height, the diameter of the plant, foliar area and dry matter (Table 2). The ST/ST/SM regimen induced plants with greater leaf area than with any other nutrition management. In the SM/SM/ST regimen, the concentration of NO3 - is maintained in 10 meq L-1 in the stages of radical growth and vegetative development, while it increases to 12 meq L-1 during the pigmentation of poinsettia. In the same regimen, the concentration of Ca2+ was 7 meq L-1 in the stage of radical growth, while it increased to 9 meq L-1 in the development of vegetation and pigmentation.
Table 2 Poinsettia growth due to the nutritional regime.
| Regime | Height (cm) | Plant diameter (cm) | Foliar area (cm2) | Bract area (cm2) | Plant dry matter (g plant-1) |
| SM/SM/SM | 23.2 ab | 40.7 cb | 1385 abc | 1751 ab | 24.2 ab |
| SM/SM/ST | 23.7 a | 44.5 a | 1546 a | 1727 ab | 26.1 a |
| SM/ST/SM | 22.5 abc | 40.9 cb | 1331 abc | 1683 b | 22.2 b |
| SM/ST/ST | 20.4 d | 39.8 c | 1527 ab | 1724 ab | 22.7 b |
| ST/SM/SM | 22.5 abc | 42.1 b | 1306 bc | 1725 ab | 22.6 b |
| ST/SM/ST | 22.1 bc | 39.6 c | 1268 c | 1555 bc | 19.3 cd |
| ST/ST/SM | 22.5 abc | 44.4 a | 1339 abc | 1976 a | 22.6 b |
| ST | 21.6 c | 40.7 cb | 1259 c | 1726 ab | 19.5 c |
| 80/120/80 | 18.6 e | 35.2 d | 1041 d | 1339 c | 17.3 d |
| CV (%) | 3.6 | 2.7 | 10.3 | 10.4 | 6.4 |
Means with the same literal in the column are statistically equal according to the Duncan test (p≤ 0.05). CV= coefficient of variation; SM= modified solution; ST= Steiner solution.
Otherwise it happened with the ST/ST/SM regime. In the stages of radical growth and vegetative development the NO3 - was maintained at 12 meq L-1 and was reduced to 10 meq L-1 during the pigmentation, while the Ca2+ remained constant at 9 meq L-1 during the whole cycle of plant. Some authors mention that in the initial stages concentrations of 17.87 to 21.43 meq L-1 of N (Berghage et al., 1987; Ecke et al., 2004) are recommended, which apparently are excessive, since it has been reported that 7.14 meq L-1 of N is sufficient to maintain adequate growth conditions (Whipker and Hammer, 1997). In this experiment, with 10 meq L-1 of NO3 - during the stages of root growth and vegetative development it was adequate to obtain poinsettia plants with outstanding morphological characteristics, but this ion must be increased to 12 meq L-1 during pigmentation.
The analysis of the nutrient extraction indicates that the SM/SM/SM and SM/SM/ST regimes caused the plants to present a statistically similar demand in N (Figure 1A), independently of the increase in the concentration NO3 - (meq L-1) in the pigmentation stage (SM/SM/SM: 10.0/10.0/10.0; SM/SM/ST: 10.0/10.0/12.0); however, plants fed with the SM/SM/ST regimen presented the best morphological characteristics (Table 2).
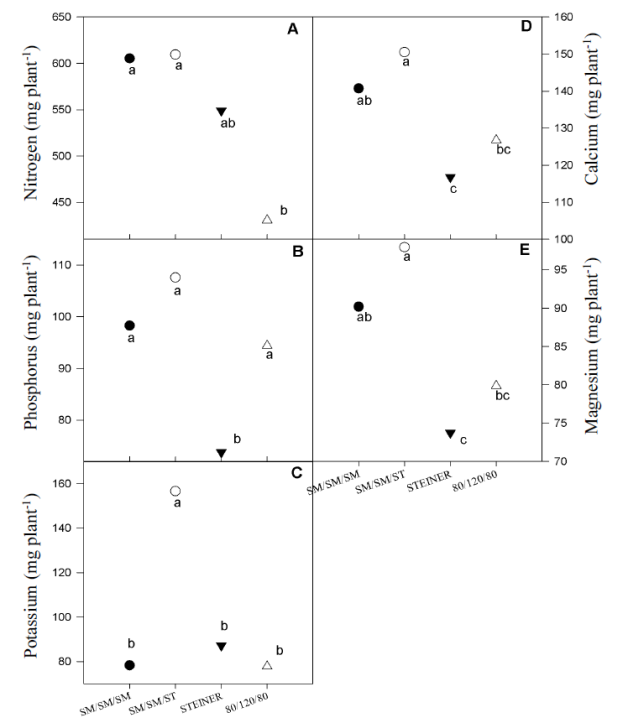
Figure 1 Nutritional extraction of the poinsettia plant due to the change of the nutritive solution.
The supply of 12 meq L-1 of NO3 - during the three phenological stages of poinsettia (ST treatment, eg. ST/ST/ST: 12/12/12), induced a significant increase in the concentration of this element (Table 3), however, their extraction decreased (Figure 1) as did the growth of the plants (Table 2). This suggests a luxury consumption; that is, poinsettia absorbed N in quantity greater than that required for optimal growth.
Table 3 Concentration of nutrients in poinsettia due to the effect of the nutrimental regime.
| Regime | N (%) | P (%) | K (%) | Ca (%) | Mg (%) |
| SM/SM/SM | 2.5 ab | 0.406 b | 0.324 b | 0.582 b | 0.373 b |
| SM/SM/ST | 2.36 b | 0.418 b | 0.606 a | 0.582 b | 0.379 b |
| ST | 2.85 a | 0.384 b | 0.455 b | 0.609 b | 0.384 b |
| 80/120/80 | 2.39 b | 0.525 a | 0.431 b | 0.706 a | 0.444 a |
| CV | 8.05 | 4.75 | 15.64 | 6.34 | 3.97 |
Means with the same literal in the column are statistically equal according to the Duncan test (p≤ 0.05). CV= coefficient of variation; SM= modified solution; ST= Steiner solution.
In this sense, Torres-Olivar et al. (2015) reported higher production of aerial and root biomass in poinsettia plants nourished with 10 meq L-1 in the stages of radical and vegetative growth, compared with plants fed with 12 and 14 meq L-1 of NO3 -. According to Steiner (1984), the plants respond to the mutual relationships between anions (NO3 - + H2PO4 - + SO4 2-), cations (K+ + Ca2++ Mg2+), total ionic concentration and pH of the nutrient solution. When the third and fourth physical-chemical characteristics are kept constant, the relative concentration of some anions can be modified with respect to the total anions, or of some cation in relation to the total of cations.
Under these conditions, the effect observed in the plants can be attributed to the modification of the anion or cation, as long as the mutual relations between the other two ions are kept constant. In this investigation, the relations between SO4 2-:H2PO4 - of 7 and K+:Mg2+ of 1.75 were maintained constant, which the effect of the nutritive solutions in the growth of the plants can be attributed to the relative concentration between NO3 - and Ca2+; however, the nutritional regime should be considered during the development of poinsettia because a better effect was observed in the morphological characteristics and dry matter production of the SM/SM/ST supply compared to ST (ST/ST/ST).
The extraction of P was greater in the plants fed with solutions modified in their phenological stages (Figure 1B). It is possible that this phenomenon is due to the different amount of P supplied during its development, according to the nutritional regime. Concentration of H2PO4 - (meq L-1) in SM/SM/SM: 1.25/ 1.25/1.25; SM/SM/ST: 1.25/1.25/1.00; ST/ST/ST: 1.0/1.0/1.0; 80/120/80: 0.8/1.2/0.8. However, with the 80/120/80 regime, the plants showed the highest concentration of P (Table 3), which can be attributed to a concentration effect as a consequence of the lower biomass production.
The extraction of K (Figure 1C) and its concentration (Table 3) was higher in the plants fed with the SM/SM/ST regimen. The concentration of this ion (meq L-1) in each of the regimens were as follows. SM/SM/ST, 8.27/7.00/7.00; SM/SM/ST, 8.27/7.00/7.00, ST/ST/ST, 7.00/7.00/7.00; 80/120/80, 5.60/8.40/5.6. It has been shown that K+ is a nutrient absorbed in greater quantity by poinsettia (Oliveira et al., 2004; Torres-Olivar et al., 2015); however, the data of the present experiment indicate that it depends on relative concentration with respect to NO3 - and H2PO4 - in the pigmentation stage.
The extraction of Ca2+ and Mg2+ was stimulated by the SM/SM/ST regimen (Figure 1D and 1E), even though the respective concentration was statistically lower than that presented by the plants nourished with the 80/120/80 regime (Table 3) and similar to the SM/SM/SM y ST/ST/ST regimes (Table 3). The concentration of Ca2+(meq L-1) according to the nutritional regimen was as follows. SM/SM/SM: 7.00/9.00/9.00; SM/SM/ST: 7/9.00/9.00; ST/ST/ST: 9.00/9.00/9.00; 80/120/80: 7.2/10.8/7.2 and to Mg2+ (meq L-1), SM/SM/SM: 4.73/4.00/4.00; SM/SM/ST: 4.73/4.00/4.00; ST/ST/ST: 4.00/4.00/4.00; 80/120/80: 3.2/4.8/3.2. The highest concentration of Ca and Mg in the control (80-120-80) is possible due to a concentration effect due to the lower production of dry matter.
The above coincides with what was reported by Reyes-Santamaría et al. (2000) who indicate that an increase in leaf area correlates positively with dry matter production; This phenomenon is related to the role of Ca in the regulation of the growth and development of cells (Tuteja and Mahajan, 2007; Valdéz-Aguilar et al., 2015) and of Mg to be a constituent of the chlorophyll molecule (Mengel and Kirkby, 1987), pigment involved in the process responsible for the generation of dry matter, photosynthesis.
Conclusions
The supply of nutrients according to the phenological stages of poinsettia affects the growth, extraction and concentration of nutrients, but depends on the nutritional regime. The regimen with the concentration (meq L-1) NO3 -:Ca2+ 10:7 in radical growth, 10: 9 in vegetative development and 12:9 in the pigmentation stage induced the poinsettias to present greater extraction of N, P, K, Ca and Mg, the concentration of K will increase, and the morphological characteristics and dry matter production will be outstanding
Literatura citada
Alcántar, G. G. y Sandoval, V. M. 1999. Manual de análisis químico de tejido vegetal. Publicación especial 10. Sociedad Mexicana de la Ciencia del Suelo. AC. Chapingo, México. 156 p. [ Links ]
Andraski T. W.; Bundy, L. G. and Brye, K. R. 2000. Crop management and corn nitrogen rate effects on nitrate leaching. J. Environ. Qual. 29(4):1095-1103. [ Links ]
Ayala-Arreola, J.; Castillo-González, A. M.; Valdez-Aguilar, L. A.; Colinas-León, M. T.; Pineda-Pineda, J. and Avitia-García, E. 2008. Effect of calcium, boron and molybdenum on plant growth and bract pigmentation in poinsettia. Rev. Fitotec. Mex. 31(2):165-172. [ Links ]
Azofeifa, A. y Moreira, M. A. 2004. Análisis de crecimiento del chile jalapeño (Capsicum annuum L. cv. Hot) en Alajuela, Costa Rica. Agron. Costarricense. 28(1):57-67. [ Links ]
Berghage, R. D.; Heins, R. D.; Carlson, W. H. and Biernbaum, J. 1987. Poinsettia production. Michigan State Coop. Ext. Serv. Bul. E-1381. 9 p. [ Links ]
Brearen, L. M. and Mulvaney, S. C. 1982. Nitrogen-total. In: Page, A. L.; Miller, R. H. and Keeney, D. R.(Eds.) Methods of soil analysis. Part 2. Chemical and microbiological properties (Agronomy 9). 2nd (Ed.). American Society of Agronomy, Soil Science Society of America. Madison, WI. 595-634 pp. [ Links ]
Chapman, H. D. y Pratt, F. P. 1973. Manual de análisis para suelos, aguas y plantas. Ed. Trillas, México. 195 p. [ Links ]
Dole, J. M. and Wilkins, H. F. 2005. Floriculture. Principles and species. 2nd (Ed.). Pearson Prentice Hall. New Jersey, USA. 1023 p. [ Links ]
Ecke, Paul III.; James, E.; Faust, P. H. D.; Higgins, A. and Williams, J. 2004. The Ecke poinsetia manual. Edited by Rick Blanchette. 268 p. [ Links ]
Gómez, R. J. y Montoya M. 2001. Soluciones nutritivas en la producción de hortalizas, frutas y flores. Editorial Año Dos Mil. Distrito Federal, México. 20-29 pp. [ Links ]
Klocke, N. L.; Watts, D. G.; Schneekloth, J. P.; Davison, D. R.; Todd, R. W. and Parkhurst, A. M. 1999. Nitrate leaching in irrigated corn and soybean in a semiarid climate. Transactions of the ASAE. 42:(6):1621-1630. [ Links ]
Martínez, M. F. 1995. Manual práctico de nochebuena. Consultoría OASIS. Morelos, México. 87 p. [ Links ]
Martínez, T. J. 2011. Sustratos, reguladores de crecimiento y fertirriego en doce variedades de nochebuena (Euphorbia pulcherrima Willd. ex Klotzsch). Tesis de licenciatura. Universidad Autónoma del Estado de Morelos. 103 p. [ Links ]
Mengel, K. y Kirkby, E. 1987. Principios de nutrición vegetal. Trad. 4ª (Ed.) por Melgar, R. J. and Ruiz, M. Internacional Potash Institute. Basilea, Suiza. 692 p. [ Links ]
Reyes, S. M. I.; Villegas, M. A.; Colinas, L. M. T. y Calderón, Z. G. 2000. Peso específico, contenido de proteína y de clorofila, en hojas de naranjo y tangerino. Agrociencia. 34(1):49-55. [ Links ]
Schwarz, M. 1995. Soilless culture management. Advanced series in Agricultural Sciences 24. Springer-Verlag. Berlin, Germany. 197 P. [ Links ]
Steiner, A. A. 1984. The universal nutrient solution. In: proceedings of sixth international congress on soilless culture. International Society for Soilless Culture. Lunteren, The Netherlands. 633-649 pp. [ Links ]
Oliveira, R. H.; Rosolem, C. A. and Trigueiro, R. M. 2004. Importance of mass flow and diffusion on the potassium supply to cotton plants as affected by soil water and potassium. Rev. Bras. Cienc. Solo. 28(3):439-445. [ Links ]
Torres, O. V.; Villegas, T. O. G.; Valdez, A. L. A.; I. Alia, T.; López, M. V. y Trejo, T. L. I. 2015. Respuesta de la nochebuena (Euphorbia pulcherrima Willd. ex Klotzsch) a la relación nitrato:calcio en tres etapas fenológicas. Rev. Mex. Cienc. Agríc. Pub. Esp. Núm. 12:2345-2357. [ Links ]
Tuteja, N. and Mahajan, S. 2007. Calcium signaling network in plants. Inglaterra. Plant Signal Behav. 2(2):79-85. [ Links ]
Whipker, B. E. and Hammer, P. A. 1997. Nutrient uptake in poinsettia during different stages of physiological development. J. Am. Soc. Hortic. Sci. 122(4):565-573. [ Links ]
Valdez, A. L. A.; Hernández, P. A.; Alvarado, C. D. and Cruz, A. A. 2015. Design of a fertilization program for chrysanthemum based on extraction of macronutrients. Rev. Mex. Cienc. Agríc. Pub. Esp. Núm. 12:2263-2276. [ Links ]
Villegas, T. O. G.; Sánchez, G. P.; Baca, C. G. A.; Rodríguez, M. M. N.; Trejo C.; Sandoval, V M. y E. Cárdenas S. 2005. Crecimiento y estado nutrimental de plántulas de tomate en soluciones nutritivas con diferente concentración de calcio y potencial osmótico. Terra Latinoam. 23(1): 49-56. [ Links ]
Vásquez, G. L. M. y Salomé, E. C. 2004. Nochebuena (Euphorbia pulcherrima) Cuetlaxóchitl. Universidad Autónoma del Estado de México (UAEM). Ed. Cigome. Toluca, Estado de México. 72-73 pp. [ Links ]
Received: April 00, 2018; Accepted: May 00, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
