











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El control de poblaciones plaga mediante poblaciones de enemigos naturales (depredadores parasitoides y entomopatógenos) es lo que denominamos control biológico, y de este, tres enfoques principales han sido desarrollados: clásico, por incremento y por conservación (Van Driesche et al., 2007). Existe consenso de que el tercero es el más importante de los tres tipos de control biológico; sin embargo, también se reconoce ha sido el menos estudiado (Ehler, 1998; Ferro y McNeil, 1998), principalmente en países en vías de desarrollo (Wlyckhuys et al., 2013).
No obstante, lo anterior el paradigma actual de control de plagas, enfermedades y malezas es mediante productos químicos órgano-sintéticos, método que por sí solo ha demostrado ineficacia en el control de plagas, púes eventualmente propicia el desarrollo de resistencia a plaguicidas y eliminación de sus enemigos naturales. Lo anterior, favorece el incremento de dosis y frecuencia de aplicación de plaguicidas, finalmente, existe un mayor riesgo de contaminación, deterioro ambiental y daños a la salud (Pimentel y Edwards, 1982).
Es urgente coadyuvar con la restauración de la biodiversidad de los agro-ecosistemas, principalmente en monocultivos extensivos. Para incrementar y conservar las poblaciones de enemigos naturales debe propiciarse primero la diversificación vegetal, donde su principal aportación es como alimento alterno en forma de néctar y polen; pero también, mayores poblaciones de herbívoros especialistas o plagas secundarias. Estos últimos funcionan como hospederos alternos de enemigos naturales que eventualmente regularán las poblaciones de insectos en los agro-ecosistemas, incluidos aquellos considerados plaga. El conocimiento de que los enemigos naturales de plagas pueden actuar también sobre insectos fitófagos no plaga (hospederos alternos) es una de las bases del control biológico por conservación (Wyckhuys et al., 2013).
Si consideramos que del total de insectos, alrededor de 5 a 6 000 especies son plagas agrícolas, mientras que cientos de miles son fitófagos no plaga (Van Lenteren, 2007), el potencial del empleo de hospederos alternos en el manejo de plagas es enorme. No obstante, son pocos los estudios desarrollados al respecto (Jacas y Urbaneja, 2010; Wyckhuys et al., 2013).
En México, la planta Asclepias curassavica L, 1753, es mejor conocida por hospedar a la mariposa monarca Danaus plexippus L, 1758 (Lepidoptera: Nymphalidae), pero también hospeda otros fitófagos importantes; entre ellos, Aphis nerii B. de F., 1841(Hemiptera: Aphididae) (Peña-Martínez et al., 2001; Cervantes-Peredo y Elizalde-Amelco, 2007). También, diversos enemigos naturales han sido identificados para A. nerii en A. curassavica, por lo que ésta planta ha sido sugerida como importante reservorio de enemigos naturales (Peña-Martínez et al., 2001). Estudios previos documentaron nueve registros de fitófagos y 21 de enemigos naturales asociados a la planta (Cortez-Madrigal et al., 2016).
Los cítricos, por otra parte, son extensamente cultivados en México y su producción se ha incrementado, actualmente en el país se ubica en el cuarto lugar a nivel mundial (Faostat, 2016). Los cítricos en México presentan una amplia diversidad de plagas, varias de origen exótico. Por ello, en México los cítricos son tal vez el cultivo donde más programas de control biológico se han implementado; como resultado, actualmente varias plagas exóticas de los cítricos están reguladas biológicamente (Arredondo y Rodríguez, 2008). Sin embargo, plagas recientemente introducidas como el pulgón café Toxoptera citricida (Kirkaldy) y el psílido asiático de los cítricos Diaphorina citri Kuwayama, amenazan con el desequilibrio del agro-ecosistema al requerir eventualmente del uso de insecticidas químicos para su control.
Entonces, los cítricos pudieran ser un buen modelo para evaluar el impacto de plantas “acompañantes” (como A. curassavica) como reservorio de hospederos alternos de enemigos naturales y su impacto en las plagas del cultivo. Una hipótesis válida sería que la inclusión de A. curassavica en plantaciones de limón persa favorecerá el incrementó de poblaciones de enemigos naturales y el control biológico de las principales plagas del cultivo. El objetivo de este estudio fue conocer el potencial de la planta A. curassavica en el manejo de plagas agrícolas, utilizando como modelo un cultivo de cítricos.
Materiales y métodos
Establecimiento del estudio
El trabajo se desarrolló del 01 de enero al 28 de diciembre de 2012 en una plantación (1 ha) de limón persa de tres años de edad, en Emiliano Zapata, Villamar, Michoacán, México. En la huerta se seleccionaron dos secciones de 20 árboles cada una, con una separación de seis hileras de árboles entre ellas; en una de las secciones se adicionó el componente A. curassavica (tratamiento) y la otra permaneció sin esa planta (testigo). A. curassavica fue colectada de canales y riachuelos de la región, posteriormente fueron trasplantadas individualmente a macetas de 17 × 21 cm con una mezcla de suelo + Peat most®+ gel (silos de agua®). Fuera de periodos de lluvias se realizaron riegos manuales por cada maceta. Un total de 50 plantas fueron distribuidas equitativamente alrededor de cada uno de los 20 árboles de limón.
Con excepción de la aplicación de plaguicidas, el manejo de la plantación se desarrolló conforme a lo programado por el productor, que incluyó riego, fertilización y una poda a principios de junio. Las aplicaciones de plaguicidas fueron reducidas al mínimo, se aplicaron compuestos de cobre para el manejo de enfermedades y dos aplicaciones de insecticidas de bajo impacto, una incluyó al organofosforado Malatión y otra al detergente Roma®. El impacto de los plaguicidas puede ser un factor de confusión para identificar el impacto de A. curassavica en la entomofauna; sin embargo, dado el reducido número de aplicaciones y que éstas fueron hechas en ambas secciones de la plantación (con y sin asclepias), se esperaría que el impacto en la densidad y fluctuación poblacional total de la entomofauna en cítricos y en asclepias sea mínimo.
Muestreos
Semanalmente se implementaron muestreos de insectos y ácaros. Para asclepias se revisó visualmente el follaje de 20 plantas al azar, las variables fueron: especies de insectos y ácaros fitófagos, especies de depredadores, parasitoides y porcentaje de áfidos parasitados. Paralelamente, árboles de cítricos con (tratamiento) y sin asclepias (testigo) fueron también revisados. Se revisaron tres brotes/cada uno de los cuatro puntos cardinales del árbol. Las variables consideradas fueron: número y especies de insectos plaga, depredadores (en sus diferentes estados) y parasitoides en 20 árboles por cada tratamiento. En ocasiones se requirió del apoyo de una lupa (5- 20X) y un contador manual.
Muestras de especímenes fueron transferidas a frascos con alcohol (75%) para su posterior identificación. Algunos insectos se mantuvieron en laboratorio hasta la emergencia de adultos o parasitoides. En campo fueron también registradas las colonias de áfidos parasitados, y se llevaron muestras de ellos al laboratorio para la obtención e identificación de parasitoides. Las especies se identificaron mediante claves dicotómicas y literatura especializada (Blackman y Eastop, 1984; Peña-Martínez et al., 2001; Bautista, 2006; Marshall, 2012). Aunque se registraron especies de parasitoides en psílidos, áfidos y minadores, los niveles de parasitismo solo fueron registrados esporádicamente, por lo que ese aspecto no se incluye en el análisis gráfico ni cuantitativo de la entomofauna de los cítricos
Análisis y procesamiento de datos
Se utilizaron estadísticas descriptivas y el conocimiento biológico de las especies. Complementariamente se practicó un análisis de varianza entre las poblaciones de insectos en cítricos con y sin A. curassavica, el cual fue procesado mediante el paquete estadístico SAS (SAS Institute Inc. 1999). Análisis de correlación fueron practicados entre las poblaciones de las principales especies de depredadores y las de las plagas clave registradas en cítricos. También, bases de datos de temperatura y precipitación fueron obtenidas de la estación climática “Presa Guaracha” de la Comisión Nacional del Agua (CNA) para su análisis de la temperatura media y la precipitación en relación con las poblaciones de insectos, lo que se hizo solo mediante exploración gráfica.
Resultados
Principales especies plaga
En bajas densidades poblacionales se registraron los áfidos T. aurantii Boyer de Fonscolombe y A. spiraecola (Patch), además el psilido asiático de los cítricos D. citri y el minador Phylocnistis citrella Stainton. Las mayores poblaciones correspondieron a los áfidos (1.86 ±4.59 insectos/3 brotes), seguidos del psílido (0.37 ±0.55 insectos/3 brotes), mientras que el minador mostró las más bajas (0.14 ±0.67) poblaciones.
Otros fitófagos registrados esporádicamente fueron Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae) y otras morfoespecies de la misma familia. También, se registraron diversas especies de escamas (Hemiptera: Coccidae); entre ellas, la escama algodonosa Icerya purchasi (Maskell). Asimismo, diversas especies de chicharritas (Hemiptera: Cicadellidae), el “gusano perro” del naranjo Papilio cresphontes Cramer (Lepidoptera: Papilionidae) y la hormiga arriera Atta sp.
Principales enemigos naturales
Los principales depredadores fueron miembros de las familias Chrysopidae y Coccinellidae. Las principales especies de coccinélidos fueron Cicloneda sanguinea L., Hippodamia convergens Guérin-Méneville, Scymnus sp. y Stethorus sp. en menor número se registraron Olla v-nigrum (Mulsant), Brachiacantha sp. también, se identificó la catarinita “Vedalia” Rodolia cardinalis (Mulsant), depredador específico de la escama I. purchasi. eventualmente, ejemplares de larvas de Syrphus sp. (Diptera: Syrphidae). Otros registros esporádicos fueron las familias Mantidae (Mantodea), Reduviidae (Hemiptera), Anthocoridae (Hemiptera), y una amplia diversidad de arañas (Araneae). Los miembros de las familias Chrysopidae (incluyendo huevos, larvas y adultos) fue el grupo de mayor abundancia, mientras que los coccinélidos mostraron la menor abundancia.
Uno de los parasitoides más abundantes en áfidos de cítricos fue la especie Lysiphlebus testaceipes Cresson (Hymenoptera: Aphidiidae), mientras que las ninfas del psílido fueron parasitadas por la especie exótica Tamarixia radiata Waterston (Hymenoptera: Eulophidae). Para los minadores, se registró el parasitoide Ageiniaspis citricola Logvinovskaya (Hymenoptera: Encyrtidae). De acuerdo con observaciones y algunos registros cuantitativos durante el estudio, el parasitismo fue más consistente en áfidos y minadores que en D. citri.
Dinámica poblacional de las principales plagas
En general, tres periodos de incidencia de plagas fueron registrados en cítricos: el primero de mayo-junio, el segundo de agosto-octubre y uno más de noviembre-diciembre. Sin embargo, durante la primera mitad del año (enero-junio) las poblaciones de insectos y plagas prácticamente estuvieron ausentes del cultivo. Solo durante los picos de mayo-junio se detectaron poblaciones de D. citri en bajas densidades. Contrariamente, durante los picos de agosto-octubre y noviembre-diciembre, el áfido T. aurantii fue la especie dominante, con sus mayores picos poblaciones durante agosto-octubre. El minador fue la plaga registrada en más bajas densidades, con picos detectables solo en el segundo semestre. El inició de los dos picos poblacionales de D. citri coincidió con las fechas de mayor temperatura (principios de mayo y principios de agosto), lo que coincidió con el desarrollo de brotes vegetativos del cultivo. La intensa brotación en agosto fue también favorecida por una previa poda de los árboles (Figura 1a y 1c).
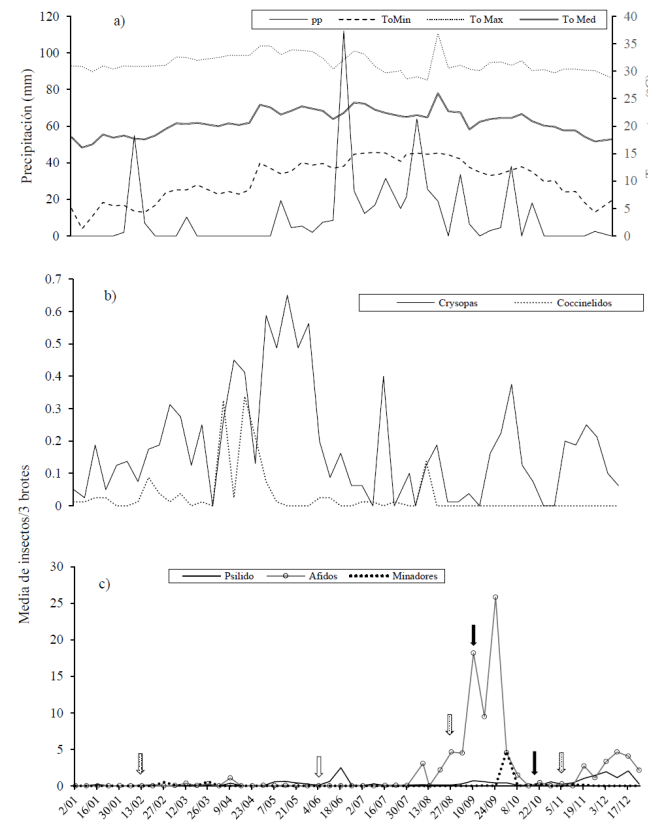
Figura 1 Relación de temperatura y precipitación (a) con los enemigos naturales (b) y las principales plagas (c) del cultivo del limón persa (testigo) en la región de Emiliano Zapata, Villamar, Michoacán 2012. Brotes vegetativos ( ), poda (
), poda ( ), insecticidas (
), insecticidas ( ).
).
De los depredadores, las crisopas fueron registradas desde el inicio de los muestreos (02 de enero) y aunque mostraron frecuentes variaciones, se observó una tendencia a incrementar sus poblaciones hasta alcanzar sus máximos picos durante la primavera (mayo-junio) posteriormente, junto con la temperatura, las poblaciones tendieron a disminuir hacia el mes de julio, con incrementos intermitentes (Figura 1b).
Respecto a los coccinelidos, sus mayores poblaciones ocurrieron en el primer semestre del año, con sus principales picos en abril-mayo. Otros picos menores ocurrieron en febrero-marzo y agosto. De finales de agosto hasta la conclusión de los muestreos, los coccinélidos estuvieron ausentes de la plantación de cítricos (Figura 1b).
Una exploración gráfica permite inferir que las poblaciones de plagas en cítricos estuvieron estrechamente relacionadas con los factores bióticos y abióticos de regulación considerados. Así, las mayores poblaciones de coccinélidos y crisopidos coincidieron con las más bajas poblaciones de psílidos y áfidos (primavera-verano), desfavorecidas también por una baja brotación del cultivo en cambio, durante el segundo semestre, cuando las poblaciones de áfidos, psílidos y minadores mostraron su mayor incremento, las poblaciones de enemigos naturales fueron bajas; esto coincidió con algunos picos de incremento de la temperatura y una intensa brotación del cultivo. No obstante, durante ese periodo se registraron reducciones intermitentes en las poblaciones de plagas, coincidentes con incrementos poblacionales de enemigos naturales, particularmente crisopas. Durante el segundo semestre existió un descenso paulatino de temperatura y precipitación, lo que coincidió con la desaparición de la familia Coccinellidae como factor de regulación natural de plagas en cítricos (Figura 1).
Efecto de Asclepias curassavica en la entomofauna de cítricos
Los principales enemigos naturales registrados en A. curassavica estuvieron estrechamente asociados con el áfido amarillo A. nerii. Se identificaron depredadores de las familias Coccinellidae, Chrysopidae, y un complejo de dípteros de las familias Syrphidae y Chamaemyiidae (por conveniencia, denominados “Syrphidae”). También se registró durante el año de estudio el parasitoide de áfidos L. testaceipes (Figura 2a). Nunca se registraron en cítricos las especies fitófagos de A. curassavica.
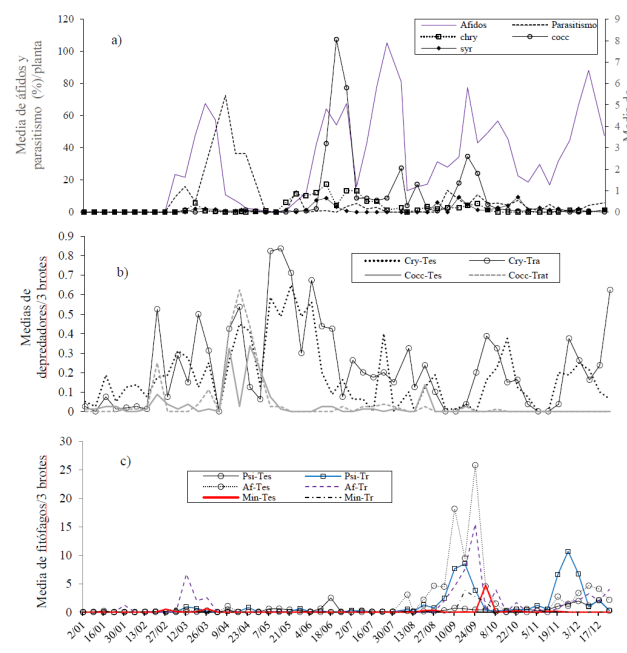
Figura 2 Distribución temporal de las principales plagas del cultivo del limón persa con (tratamiento) y sin (testigo) presencia de Asclepias curassavica en la región de Emiliano Zapata, Villamar, Michoacán 2012. Psi= psílido; Af= afidos; Min= minador; Tr= tratamiento; Tes= testigo.
La tendencia poblacional de las principales plagas fue similar entre tratamiento y testigo. Dadas las bajas poblaciones durante el primer semestre del año, no fue posible identificar algún efecto de A. curassavica en la incidencia de plagas en cítricos; no obstante, las altas poblaciones de enemigos naturales registradas en A. curassavica sugieren algún efecto benéfico de la planta (Figura 2a). La tendencia gráfica de mayores poblaciones de coccinélidos y crisopidos en los cítricos donde se ubicó A. curassavica así lo sugieren. Uno de los depredadores más abundantes en A. curassavica fue la especie Cycloneda sanguinea, también registrada frecuentemente en cítricos. A partir de que se colocaron las macetas con asclepias (03 de marzo) las poblaciones de depredadores incrementaron respecto a la plantación donde no se tuvo la planta (Figura 2b).
Fue durante el segundo semestre cuando se identificó con mayor claridad el efecto de A. curassavica en áfidos y psílidos. Una exploración gráfica permite inferir que la planta influyó en la reducción de poblaciones de áfidos, comparativamente con la huerta donde no se colocaron plantas de A. curassavica.
Contrariamente, una tendencia de mayores niveles poblaciones de psílidos fueron registrados en cítricos con A. curassavica, respecto a donde no se tuvo la planta. Dadas las bajas densidades del minador P. citrella, gráficamente no se detectó algún efecto de asclepias en las poblaciones de la plaga (Figura 2c), lo que no significa que no exista.
El análisis estadístico de las poblaciones de insectos, anuales y por semestre, mostró diferencias significativas (p≤ 0.05) entre ambas plantaciones. La población media anual de D. citri fue más abundante en el tratamiento (asclepias) que en el testigo, mientras que para áfidos y minadores solo se registraron tendencias de menores poblaciones donde se adicionó la planta asclepias (trat). El análisis estadístico de las poblaciones del minador confirmó la tendencia gráfica de tratamiento y testigo, con niveles poblacionales similares en ambas plantaciones. De los depredadores, las crisopas fueron estadísticamente más abundantes en cítricos con Asclepias que en el testigo sin la planta; mientras los coccinelidos solo mostraron tendencia de mayor abundancia donde se adicionó la planta (Figura 3a).
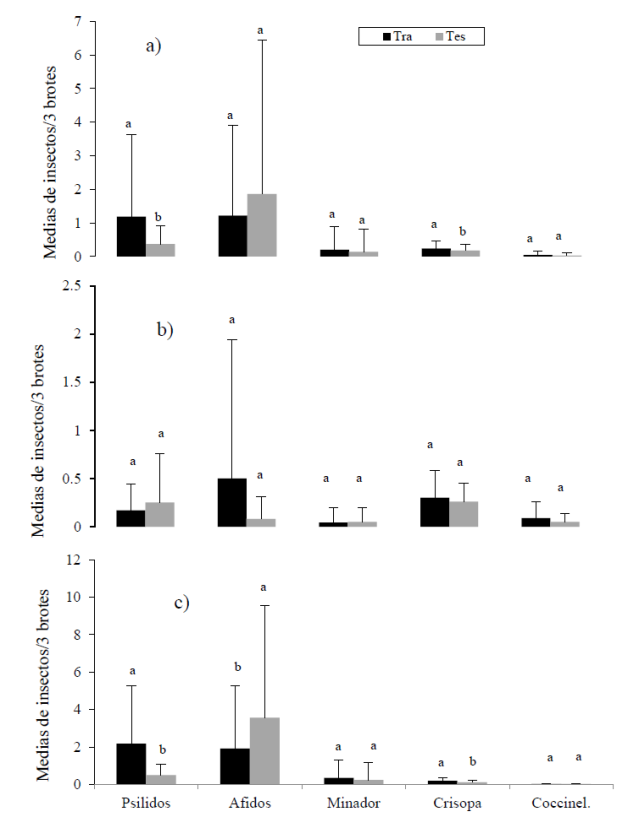
Figura 3 Medias de las principales plagas y sus enemigos naturales en cítricos con (tra) y sin (tes) la presencia de la panta Asclepias curassavica; a) Año 2012; b) primer periodo (enero-junio); c) segundo periodo (julio-diciembre). Emiliano Zapata, Villamar, Michoacán 2012. Pareja de barras con la misma letra no difieren estadísticamente (Tukey, p≤ 0.05).
El análisis semestral permitió identificar diferencias que fueron enmascaradas en el análisis anual. Durante el primer semestre del año (enero-junio), cuando ocurrieron las más bajas poblaciones de plagas, las diferencias entre tratamientos no fueron significativas (p≤ 0.05), solo se identificaron tendencias. Así, los psilidos mostraron más bajos niveles poblacionales en el tratamiento que en el testigo, coincidiendo con las altas poblaciones de coccinélidos presentes en A. curassavica. Los áfidos fueron más abundantes en el tratamiento con asclepias. En minadores, solo se registró una tendencia de menores poblaciones en el tratamiento. La tendencia en enemigos naturales fue similar a la del análisis anual (Figura 3b).
En el segundo semestre (julio-diciembre) fue cuando se registraron las más claras diferencias estadísticas entre las plantaciones con y sin asclepias. La más baja población de áfidos ocurrió en la plantación con asclepias y lo contrario ocurrió para los psílidos. En tanto que los minadores solo mostraron una tendencia de mayores poblaciones en la plantación con asclepias. Las poblaciones de crisopas fueron mayores en la plantación con Asclepias, mientras los coccinelidos prácticamente desaparecieron durante ese periodo (Figura 3 c).
Discusión
No obstante, los bajos niveles poblacionales de plagas y el reducido número de plantas de A. curassavica establecidas (3/árbol), los resultados muestran evidencias de que la planta influyó en los niveles poblacionales de dos de las principales plagas de los cítricos: T. aurantii y D. citri. Ello se atribuye a la aportación de enemigos naturales incrementados en asclepias entre ellos, diversas especies de Coccinelidos y el parasitoide L. testaceipes. La reducción significativa (46.3%) de áfidos durante el segundo semestre en la plantación con A. curassavica así lo sugiere. Sin embargo, también se registró un incremento (77%) en las poblaciones de D. citri en la misma plantación. Por el contrario, las tendencias poblacionales durante el primer semestre sugieren un efecto contrario; más bajas poblaciones del psílido y mayores de áfidos en los cítricos con Asclepias.
Las causas de los resultados pueden ser múltiples, pero la principal pudiera ser que, al abatirse las poblaciones de alguno de los dos insectos chupadores por efecto de sus enemigos naturales (coccinélidos para psílidos, y L. testaceipes para áfidos) favorecidos por la presencia de A. curassavica, la disponibilidad de brotes vegetativos de cítricos fuera mayor. Esos nichos vacíos permitirían una mayor colonización del insecto sujeto a menor biocontrol (psílidos o áfidos), lo que dependió de la época del año (factores climáticos). Una situación similar ha sido registrada en plagas del mismo gremio (consumen el mismo recurso) por ejemplo, el áfido de las leguminosas A. craccivora colonizó brotes reproductivos de cacao en Tabasco, México. Sólo cuando el principal áfido del cultivo, T. aurantii, estuvo ausente por factores climáticos limitativos (Sánchez y Cortez, 1998). En el presente estudio, se registraron picos de temperatura cercanos a 25 oC, condiciones que han sido mencionadas como las más favorables para el desarrollo de ninfas del psílido.
Por otro lado, la presencia de fitófagos dominantes en los agroecosistemas puede significar diferentes cosas, dependiendo si son especies exóticas o nativas. En el primer caso tales especies llegan a ser abundantes simplemente por estar ausentes sus enemigos naturales originales (Valladares y Salvo, 1999). D. citri es una plaga recientemente introducida a México, y aunque diversas especies de enemigos naturales han sido mencionados para el psilido (incluido su parasitoide original T. radiata), existe consenso en que los coccinelidos son responsables de la mayor mortalidad de ninfas de D. citri (Ruiz-Cancino et al., 2008; Cortez-Mondaca et al., 2011; González et al., 2012; Grafton-Cardwell et al., 2013). La ausencia de coccinélidos en el segundo periodo del estudio pudo contribuir también con el incremento del psilido.
Existe consenso de que las crisopas y sirfidos son dos de los principales grupos de enemigos naturales de D. citri; sin embargo, sírfidos y crisópidos raramente han sido observados alimentándose del psílido (Grafton-Cardwell et al., 2013). Lo anterior coincide con lo observado en el presente estudio, aunque las crisopas estuvieron presentes durante todo el año en la plantación con Asclepias, dadas las abundantes poblaciones de áfidos (cítricos + asclepias), las crisopas pudieron preferir alimentarse de áfidos que de psílidos. Una mayor correlación entre poblaciones de crisopas y áfidos (r= -0.51, p≤ 0.05) que entre crisopas y D. citri (r= -0.13, p≤ 0.05) así lo sugiere. Adicionalmente, observaciones de campo y laboratorio evidenciaron que en el segundo semestre del año las poblaciones de T. radiata fueron tan escasas, que tuvieron un pobre impactó en las poblaciones del psílido, tal como ha sido registrado en otras regiones de México (Hernández-Guerra et al., 2011).
La conservación de hospederos alternos ha sido una de las estrategias ampliamente recomendada para la conservación de enemigos naturales de plagas (Landis et al., 2000; Salvo y Valladares, 2007; Wyckhuys et al., 2013). Al respecto, se sabe que en España es común establecer la planta Nerium oleander en linderos de huertas de naranja. N. oleander hospeda al pulgón A. nerii, especie inocua a los cítricos, pero importante hospedera de parasitoides que más tarde impactan en poblaciones de áfidos plaga del cultivo (Jacas y Urbaneja, 2010). Lo anterior es acorde con los resultados obtenidos en el presente estudio donde la presencia de A. nerii en A. curassavica favoreció el incremento de enemigos naturales que redujeron las poblaciones de plagas en cítricos. Sin embargo, dicha estrategia eventualmente pudiera propiciar el incremento de fitófagos no deseados, tal como ocurrió con el psílido de los cítricos D. citri durante el segundo semestre del año.
Por lo anterior, la estrategia del empleo de hospederos alternos para la conservación de enemigos naturales involucra relaciones tróficas complejas; por ejemplo, fitófagos-plantas, enemigos naturales-fitófagos, entre los mismos enemigos naturales, o bien, entre enemigo natural-fitófago-planta. Por lo tanto, un mayor entendimiento de las relaciones tróficas entre los organismos involucrados y su medio ambiente deben ser objetivos de estudios futuros. Por ejemplo, además de A. nerii, los otros fitófagos registrados en A. curassavica pueden ser también importantes reservorios de enemigos naturales así, los huevos de Danaus plexippus fueron importante hospederos de T. pretiossum (Cortez-Madrigal et al., 2014) y este parasitoide ha sido ampliamente utilizado en producción masiva para el control de lepidópteros plaga (Hoffmann y Frodsham, 1993). El hecho de que las especies de fitófagos registradas en A. curassavica (Cortez-Madrigal et al., 2016) no sean plaga de cultivos (Blackman y Eastop, 1984; Cervantes-Peredo y Elizalde-Amelco, 2007) indica el potencial de la planta para utilizarse como refugio e incremento de enemigos naturales para el manejo de plagas en los agroecosistemas.
Por lo anterior, el empleo de plantas “acompañantes” para la conservación de hospederos alternos de enemigos naturales, debe visualizarse como una estrategia más del manejo integrado de plagas, donde el objetivo central debe ser lograr la estabilidad del agroecosistema más que el control de plagas específicas. En el cultivo de los cítricos, por ejemplo, debe mantenerse vigilancia de las principales plagas mediante muestreos rutinarios; y el manejo, tanto de la planta acompañante como de los cítricos (podas). Lo anterior, indicará la necesidad de la utilización de otras estrategias para el control de plagas emergentes; por ejemplo, para D. citri sería la liberación de T. radiata o en último caso, el empleo de insecticidas de bajo impacto.
La inclusión de plantas como A. curassavica en los cultivos cubriría el importante vacío que existe en cuanto a diversidad biológica de los sistemas agrícolas altamente tecnificados, pero pobres en cuanto a diversidad de enemigos naturales. Otros estudios han documentado la importancia de otras especies de Asclepias, pero solo como fuente de néctar para enemigos naturales (Fielder y Landis, 2007). Con el presente estudio se fortalece la idea de que A. curassavica puede ser un importante reservorio de enemigos naturales (Peña-Martínez et al., 2001) y se muestran evidencias de su potencial en el manejo de plagas de los cítricos.
A. curassavica es una planta perenne, ampliamente distribuida (Juárez-Jaimes et al., 2007) y fácil de reproducir, lo que facilita su empleo como reservorio de hospederos alternos y fuente de néctar para enemigos naturales. Además de su presencia in situ, A. curassavica también pudiera aprovecharse para incrementar poblaciones de A. nerii en condiciones de invernadero, infestarlas con el parasitoide L. testaceipes y posteriormente transferir las macetas con las “momias” a los agroecosistemas donde existan problemas de áfidos.
Un aprovechamiento que no ha sido mencionado en las plantas acompañantes es su utilidad como plantas “centinela” en los agroecosistemas esta planta junto con sus fitófagos, indicaría la presencia o ausencia de una amplia gama de enemigos naturales de plagas, que eventualmente compartiría con los cultivos. Las bajas poblaciones o ausencia de enemigos naturales pudiera ser la señal para implementar otras estrategias de control; por ejemplo, la ausencia de coccinélidos en el segundo semestre del año sugeriría una mayor vigilancia de D. citri.
Conclusiones
La presencia de Asclepias curassavica en plantaciones de limón persa mostró un efecto significativo en la incidencia de plagas, su impacto dependió de la especie plaga y la época del año. El aprovechamiento de la planta debe visualizarse como parte de un manejo integrado de plagas, donde el objetivo central sea la estabilidad de la entomofauna del agroecosistema y no el control biológico de plagas específicas.

