










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 no.2 Texcoco Fev./Mar. 2018
https://doi.org/10.29312/remexca.v9i2.1073
Articles
Potential of Aclepias curassavica L. (Apocynaceae) in the biological control of pests
1Instituto Politécnico Nacional-CIIDIR-Unidad Michoacán. Justo Sierra Núm. 28, col. Centro, Jiquilpan, Michoacán. Tel. 35330218.
To know the potential of Asclepias curassavica L. in the management of pests, from January 1 to December 28, 2012, a study was established in a plantation of “Persian” lemon Citrus latifolia in Emiliano Zapata, Villamar, Michoacán, Mexico. One section (treatment) included the A. curassavica plant and the other remained without that species (control). Weekly samples documented species and number of predators, phytophages, parasitoids, and parasitism levels in aphids. Toxoptera aurantii, Aphis spiraecola, Diaphorina citri and Phyllocnistis citrella were the main pests. Its largest populations occurred in the second half of the year and coincided with the main periods of budding of the crop. The main natural enemies observed in A. curassavica were the families Coccinellidae, Syrphidae, Chamaemyiidae, Chrysopidae and the parasitoid Lysiphlebus testaceipes. The Coccinellids predominated during the first semester of the year, while the other groups were frequent throughout the year. The results show a 46.3% reduction in citrus aphid populations where Asclepias were established; however, D. citri populations increased 77%. Probably the greatest biocontrol of aphids favored the availability of empty niches (vegetative shoots) and consequently the populations of the psyllid. The wide diversity of natural enemies registered in A. curassavica suggests a high potential of the plant to be used in the regulation and stability of the entomowildlife of agroecosystems, rather than in the control of specific pests.
Keywords: Diaphorina citri; Toxoptera aurantii; Phyllocnistis citrella; Lysiphlebus testaceipes
Para conocer el potencial de Asclepias curassavica L. en el manejo de plagas, del 01 de enero al 28 de diciembre de 2012, se estableció un estudio en una plantación de limón “persa” Citrus latifolia en Emiliano Zapata, Villamar, Michoacán, México. Una sección (tratamiento) incluyó la planta A. curassavica y la otra se mantuvo sin esa especie (testigo). Muestreos semanales documentaron especies y número de depredadores, fitófagos, parasitoides, y niveles de parasitismo en áfidos. Toxoptera aurantii, Aphis spiraecola, Diaphorina citri y Phyllocnistis citrella, fueron las principales plagas. Sus mayores poblaciones ocurrieron en el segundo semestre del año y coincidieron con los principales periodos de brotación del cultivo. Los principales enemigos naturales observados en A. curassavica fueron las familias Coccinellidae, Syrphidae, Chamaemyiidae, Chrysopidae y el parasitoide Lysiphlebus testaceipes. Los Coccinellidos predominaron durante el primer semestre del año, mientras que los otros grupos fueron frecuentes durante todo el año. Los resultados muestran una reducción de 46.3% en las poblaciones de áfidos de los cítricos donde se establecieron Asclepias; sin embargo, las poblaciones de D. citri se incrementaron 77%. Probablemente el mayor biocontrol de áfidos favoreció la disponibilidad de nichos vacíos (brotes vegetativos) y consecuentemente las poblaciones del psílido. La amplia diversidad de enemigos naturales registrados en A. curassavica sugiere un alto potencial de la planta para utilizarse en la regulación y estabilidad de la entomofauna de los agroecosistemas, más que en el control de plagas específicas.
Palabras clave: Diaphorina citri; Lysiphlebus testaceipes; Phyllocnistis citrella; Toxoptera aurantii
Introduction
The control of plague populations through populations of natural enemies (parasitoid and entomopathogenic predators) is what we call biological control, and from this, three main approaches have been developed: classical, incremental and conservation (Van Driesche et al., 2007). There is a consensus that the third is the most important of the three types of biological control; however, it is also acknowledged to have been the least studied (Ehler, 1998, Ferro and McNeil, 1998), mainly in developing countries (Wlyckhuys et al., 2013).
However, the current paradigm of control of pests, diseases and weeds is through organophosphate chemicals, a method that by itself has shown ineffectiveness in the control of pests, which eventually promotes the development of resistance to pesticides and elimination of their natural enemies. The above, favors the increase of dose and frequency of application of pesticides, finally, there is a greater risk of contamination, environmental deterioration and damage to health (Pimentel and Edwards, 1982).
It is urgent to contribute to the restoration of the biodiversity of agro-ecosystems, mainly in extensive monocultures. In order to increase and conserve the populations of natural enemies, first the plant diversification must be encouraged, where its main contribution is as an alternative food in the form of nectar and pollen; but also, greater populations of specialist herbivores or secondary pests. The latter function as alternate hosts of natural enemies that will eventually regulate insect populations in agro-ecosystems, including those considered pests. The knowledge that natural enemies of pests can also act on non-pest phytophagous insects (alternate hosts) is one of the bases of biological control by conservation (Wyckhuys et al., 2013).
If we consider that from the total number of insects, around 5 to 6 000 species are agricultural pests, while hundreds of thousands are non-plague phytophagous (Van Lenteren, 2007), the potential of the use of alternate hosts in pest management is enormous. However, there are few studies developed in this regard (Jacas and Urbaneja, 2010; Wyckhuys et al., 2013).
In Mexico, the plant Asclepias curassavica L, 1753, is best known for hosting the monarch butterfly Danaus plexippus L, 1758 (Lepidoptera: Nymphalidae), but also hosts other important phytophages; among them, Aphis nerii B. of F., 1841 (Hemiptera: Aphididae) (Peña-Martínez et al., 2001; Cervantes-Peredo and Elizalde-Amelco, 2007). Also, several natural enemies have been identified for A. nerii in A. curassavica, so this plant has been suggested as an important reservoir of natural enemies (Peña-Martínez et al., 2001). Previous studies documented nine phytophagous records and 21 of natural enemies associated with the plant (Cortez-Madrigal et al., 2016).
Citrus fruits, on the other hand, are extensively cultivated in Mexico and their production has increased, currently in the country it is ranked fourth in the world (Faostat, 2016). Citrus fruits in Mexico have a wide variety of pests, several of exotic origin. Therefore, in Mexico citrus fruits are perhaps the crop where more biological control programs have been implemented; as a result, currently several exotic citrus pests are regulated biologically (Arredondo and Rodríguez, 2008). However, recently introduced pests such as the brown louse Toxoptera citricida (Kirkaldy) and the Asian citrus psyllid Diaphorina citri Kuwayama, threaten the imbalance of the agro-ecosystem by eventually requiring the use of chemical insecticides to control them.
Then, citrus fruits could be a good model to evaluate the impact of “companion” plants (such as A. curassavica) as a reservoir of alternate hosts of natural enemies and their impact on crop pests. A valid hypothesis would be that the inclusion of A. curassavica in Persian lemon plantations will favor the increase of populations of natural enemies and the biological control of the main pests of the crop. The objective of this study was to know the potential of the A. curassavica plant in the management of agricultural pests, using as a model a citrus crop.
Materials and methods
Establishment of the study
The work was carried out from January 1 to December 28, 2012 in a plantation (1 ha) of Persian lemon of three years of age, in Emiliano Zapata, Villamar, Michoacán, Mexico. In the garden two sections of 20 trees each were selected, with a separation of six rows of trees between them; in one of the sections component A. curassavica (treatment) was added and the other remained without that plant (control). A. curassavica was collected from channels and streams of the region, later they were transplanted individually to 17 × 21 cm pots with a soil mixture + Peat most®+ gel (silos de agua®). Outside of rainy periods, manual irrigations were carried out for each pot. A total of 50 plants were evenly distributed around each of the 20 lemon trees.
With the exception of the application of pesticides, the management of the plantation was developed according to the schedule by the producer, which included irrigation, fertilization and pruning at the beginning of June. Pesticide applications were reduced to a minimum, copper compound were applied for disease management and two applications of low impact insecticides, one included the organophosphate Malathion and another one the Roma® detergent. The impact of pesticides can be a confounding factor to identify the impact of A. curassavica on the entomowildlife; however, given the small number of applications and that these were made in both sections of the plantation (with and without Asclepias), it would be expected that the impact on the density and total population fluctuation of the entomowildlife in citrus and Asclepias would be minimal.
Sampling
Samplings of insects and mites were implemented weekly. For Asclepias the foliage of 20 plants was visually reviewed at random, the variables were: species of insects and phytophagous mites, predator species, parasitoids and percentage of parasitized aphids. In parallel, citrus trees with (treatment) and without Asclepias (control) were also reviewed. Three buds/each of the four cardinal points of the tree were reviewed. The variables considered were: number and species of insect pests, predators (in their different states) and parasitoids in 20 trees for each treatment. Sometimes the support of a magnifying glass (5- 20X) and a manual counter were required.
Samples of specimens were transferred to bottles with alcohol (75%) for later identification. Some insects were kept in the laboratory until the emergence of adults or parasitoids. The colonies of parasitized aphids were also recorded in the field, and samples were taken to the laboratory for obtaining and identifying parasitoids. The species were identified through dichotomous keys and specialized literature (Blackman and Eastop, 1984; Peña-Martínez et al., 2001; Bautista, 2006; Marshall, 2012). Although parasitoid species were recorded in psyllids, aphids and leaf miners, parasitism levels were only recorded sporadically, so this aspect is not included in the graphical or quantitative analysis of the citrus entomowildlife.
Analysis and data processing
Descriptive statistics and biological knowledge of the species were used. In addition, an analysis of variance was carried out among insect populations in citrus fruits with and without A. curassavica, which was processed using the SAS statistical package (SAS Institute Inc. 1999). Correlation analyzes were performed between the populations of the main predator species and those of the key citrus registered pests. Also, temperature and precipitation databases were obtained from the “Presa Guaracha” climate station of the National Water Commission (CNA) for its analysis of average temperature and precipitation in relation to insect populations, which was done only by graphic exploration.
Results
Main species plague
At low population densities, the aphids T. aurantii Boyer of Fonscolombe and A. spiraecola (Patch) were registered, as well as the Asian citrus psyllid D. citri and the Phylocnistis citrella Stainton miner. The largest populations corresponded to aphids (1.86 ±4.59 insects/3 shoots), followed by the psyllid (0.37 ±0.55 insects/3 shoots), while the leafminer showed the lowest (0.14 ±0.67) populations.
Other phytophagous recorded sporadically were Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae) and other morphospecies of the same family. Also, several species of scales were recorded (Hemiptera: Coccidae); among them, the cottony scale Icerya purchasi (Maskell). Also, different species of leafhoppers (Hemiptera: Cicadellidae), the “worm dog” of the orange tree Papilio cresphontes Cramer (Lepidoptera: Papilionidae) and the ant arriera Atta sp.
Main natural enemies
The main predators were members of the families Chrysopidae and Coccinellidae. The main coccinellid species were Cicloneda sanguinea L., Hippodamia convergens Guerin-Meneville, Scymnus sp. and Stethorus sp. in lesser number Olla v-nigrum (Mulsant), Brachiacantha sp. also, the catarinita “Vedalia” Rodolia cardinalis (Mulsant), specific predator of the scale I. purchasi was identified, eventually, larvae specimens of Syrphus sp. (Diptera: Syrphidae). Other sporadic records were the families Mantidae (Mantodea), Reduviidae (Hemiptera), Anthocoridae (Hemiptera), and a wide variety of spiders (Araneae). Members of the Chrysopidae families (including eggs, larvae and adults) were the most abundant group, while the coccinellids showed the lowest abundance.
One of the most abundant parasitoids in citrus aphids was the species Lysiphlebus testaceipes Cresson (Hymenoptera: Aphidiidae), while the nymphs of the psyllid were parasitized by the exotic species Tamarixia radiata Waterston (Hymenoptera: Eulophidae). For the miners, the parasitoid Ageiniaspis citricola Logvinovskaya (Hymenoptera: Encyrtidae) was recorded. According to observations and some quantitative records during the study, parasitism was more consistent in aphids and leaf miners than in D. citri.
Population dynamics of the main pests
In general, three periods of incidence of pests were recorded in citrus: the first of may-june, the second of August-October and one more of November December. However, during the first half of the year (January June) insect and pest populations were practically absent from the crop. Only during the peaks of May-June were D. citri populations detected at low densities. Contrarily, during the peaks of August-October and November December. The aphid T. aurantii was the dominant species, with its largest peaks populations during August-October. The quarry was the registered pest in lower densities, with detectable peaks only in the second semester. The start of the two population peaks of D. citri coincided with the higher temperature dates (beginning of May and beginning of August), which coincided with the development of vegetative shoots of the crop. The intense sprouting in august was also favored by a previous pruning of the trees (Figure 1a y 1c).

Figure 1. Relationship of temperature and precipitation (a) with the natural enemies (b) and the main pests (c) of the Persian lemon crop (control) in the region of Emiliano Zapata, Villamar, Michoacan 2012. Vegetative shoots ( ), pruning (
), pruning ( ), insecticides (
), insecticides ( ).
).
Of the predators, the lacewings were recorded from the beginning of the samplings (January 2) and although they showed frequent variations, a tendency was observed to increase their populations until reaching their maximum peaks during the spring (May-June) later, together with the temperature, the populations tended to decrease towards the month of July, with intermittent increases (Figure 1b).
Regarding the coccinellids, their largest populations occurred in the first half of the year, with its main peaks in April-May. Other minor peaks occurred in February-March and August. From the end of august until the conclusion of the samplings, the coccinellids were absent from the citrus plantation (Figure 1b).
A graphical exploration allows us to infer that citrus plague populations were closely related to the biotic and abiotic regulation factors considered. Thus, the largest populations of coccinellids and chrysopids coincided with the lower populations of psyllids and aphids (Spring-Summer), also disadvantaged by a low budding of the crop in contrast, during the second half, when populations of aphids, psyllids and leaf miners showed their greatest increase, populations of natural enemies were low; this coincided with some peaks of temperature increase and intense budding of the crop. However, during this period intermittent reductions were recorded in the populations of pests, coinciding with population increases of natural enemies, particularly lacewings. During the second semester, there was a gradual decrease in temperature and precipitation, which coincided with the disappearance of the Coccinellidae family as a natural regulation factor for citrus pests (Figure 1).
Effect of Asclepias curassavica in the citrus entomowildlife
The main natural enemies recorded in A. curassavica were closely associated with the yellow aphid A. nerii. Predators of the families Coccinellidae, Chrysopidae, and a dipterous complex of the families Syrphidae and Chamaemyiidae (for convenience, called “Syrphidae”) were identified. The aphid parasitoid L. testaceipes was also recorded during the study year (Figure 2a). The phytophagous species of A. curassavica were never recorded in citrus.
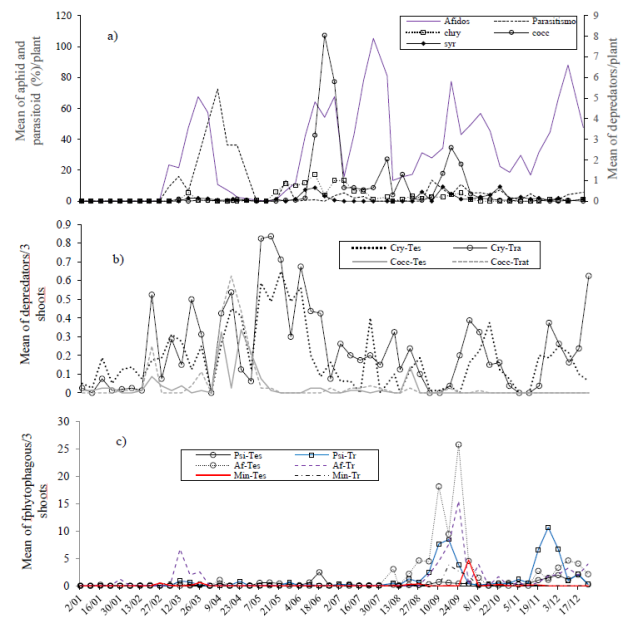
Figure 2 Temporal distribution of the main pests of the Persian lemon crop with (treatment) and without (control) presence of Asclepias curassavica in the region of Emiliano Zapata, Villamar, Michoacan 2012. Psi= psyllid; Af= aphids; Min= miner; Tr= treatment; Tes= witness.
The population trend of the main pests was similar between treatment and control. Given the low populations during the first semester of the year, it was not possible to identify any effect of A. curassavica on the incidence of citrus pests; however, the high populations of natural enemies recorded in A. curassavica suggest some beneficial effect of the plant (Figure 2a). The graphic trend of larger populations of coccinellids and chrysophobia in citrus fruits where A. curassavica was located, suggest this. One of the most abundant predators in A. curassavica was Cycloneda sanguinea, also frequently recorded in citrus fruits. After the pots were placed with Asclepias (March 3), the populations of predators increased with respect to the plantation where the plant was not present (Figure 2b).
It was during the second semester that the effect of A. curassavica on aphids and psyllids was more clearly identified. A graphic exploration allows us to infer that the plant influenced the reduction of aphid populations, comparatively with the orchard where A. curassavica plants were not placed.
Contrarily, a trend of higher levels of psyllid populations were registered in citrus fruits with A. curassavica, with respect to where the plant was not. Given the low densities of the P. citrella leafminer, no effect of Asclepias was detected graphically on the populations of the pest (Figure 2c), which does not mean that it does not exist.
The statistical analysis of insect populations, annual and by semester, showed significant differences (p≤ 0.05) between both plantations. The average annual population of D. citri was more abundant in the treatment (Asclepias) than in the control, while for aphids and miners only minor population trends were recorded where the plant Asclepias (trat) was added. The statistical analysis of the populations of the miner confirmed the graphical trend of treatment and control, with similar population levels in both plantations. Of the predators, lacewings were statistically more abundant in citrus with Asclepias than in the control without the plant; while coccinellids only showed a trend of greater abundance where the plant was added (Figure 3a).

Figure 3 Means of the main pests and their natural enemies in citrus fruits with (tra) and without (tes) the presence of the Asclepias curassavica panta. A) Year 2012; B) First period (January June); C) Second period (July December). Emiliano Zapata, Villamar, Michoacán 2012. Pair of bars with the same letter do not differ statistically (Tukey, p≤ 0.05).
The biannual analysis allowed to identify differences that were masked in the annual analysis. During the first semester of the year (january-june), when the lowest pest populations occurred, the differences between treatments were not significant (p≤ 0.05), only trends were identified. Thus, the psilids showed lower population levels in the treatment than in the control, coinciding with the high populations of coccinellids present in A. curassavica. Aphids were more abundant in the treatment with Asclepias. In miners, there was only a trend of lower populations in the treatment. The trend in natural enemies was similar to that in the annual analysis (Figure 3b).
In the second semester (July December) was when the clearest statistical differences between the plantations with and without Asclepias were registered. The lowest population of aphids occurred in the plantation with Asclepias and the opposite occurred for the psyllids. While the miners only showed a trend of higher populations in the plantation with Asclepias. The lacewing populations were higher in the Asclepias plantation, while the coccinellids practically disappeared during that period (Figure 3c).
Discussion
However, the low population levels of pests and the small number of plants of A. curassavica established (3/tree), the results show evidence that the plant influenced the population levels of two of the main pests of citrus: T. aurantii and D. citri. This is attributed to the contribution of natural enemies increased in Asclepias among them, several species of Coccinelidae and the parasitoid L. testaceipes. The significant reduction (46.3%) of aphids during the second semester in the plantation with A. curassavica suggests it. However, there was also a significant increase (77%) in D. citri populations in the same plantation. On the contrary, the population trends during the first semester suggest an opposite effect; lower populations of the psyllid and greater aphids in citrus fruits with Asclepias.
The causes of the results can be multiple, but the main one could be that, when the populations of one of the two sucking insects are killed by the effect of their natural enemies (coccinellids for psyllids, and L. testaceipes for aphids) favored by the presence of A. curassavica, the availability of vegetative shoots of citrus was greater. Those empty niches would allow a greater colonization of the insect subject to lower biocontrol (e.g. psyllids or aphids), which depended on the time of year (climatic factors). A similar situation has been registered in pests of the same guild (they consume the same resource) for example, the aphid of the legumes A. craccivora colonized reproductive buds of cocoa in Tabasco, Mexico. Only when the main aphid of the crop, T. aurantii, was absent due to limiting climatic factors (Sánchez and Cortez, 1998). In the present study, temperature peaks near 25 oC were recorded, conditions that have been mentioned as the most favorable for the development of psyllid nymphs.
On the other hand, the presence of dominant phytophages in agroecosystems can mean different things, depending on whether they are exotic or native species. In the first case, such species become abundant simply because their original natural enemies are absent (Valladares and Salvo, 1999). D. citri is a pest recently introduced to Mexico, and although several species of natural enemies have been mentioned for the psilid (including its original parasitoid T. radiata), there is a consensus that coccinellids are responsible for the higher mortality of D. citri nymphs (Ruiz-Cancino et al., 2008; Cortez-Mondaca et al., 2011; González et al., 2012; Grafton-Cardwell et al., 2013). The absence of coccinellids in the second period of the study could also contribute to the increase of the psyllid.
There is a consensus that lacewings and syrphids are two of the main groups of natural enemies of D. citri; however, syrphids and chrysopids have rarely been observed feeding on the psyllid (Grafton-Cardwell et al., 2013). The above coincides with what was observed in the present study, although the lacewings were present throughout the year in the plantation with Asclepias, given the abundant populations of aphids (citrus + asclepias), the lacewings could prefer to feed on aphids than on psyllids. A higher correlation between lacewings and aphids (r= -0.51, p≤ 0.05) than between lacewings and D. citri (r= -0.13, p≤ 0.05) suggests this. Additionally, field and laboratory observations showed that in the second semester of the year the populations of T. radiata were so scarce, that they had a poor impact on the populations of the psyllid, as has been registered in other regions of Mexico (Hernández-Guerra et al., 2011).
The conservation of alternate hosts has been one of the widely-recommended strategies for the conservation of natural enemies of pests (Landis et al., 2000; Salvo and Valladares, 2007; Wyckhuys et al., 2013). In this regard, it is known that in Spain it is common to establish the Nerium oleander plant in boundaries of orange orchards. N. oleander hosts the A. nerii aphid, a citrus-free species, but an important host of parasitoids that later impact on pest populations of the crop (Jacas and Urbaneja, 2010). The above is in agreement with the results obtained in the present study where the presence of A. nerii in A. curassavica favored the increase of natural enemies that reduced citrus plague populations. However, this strategy could eventually lead to the increase of unwanted phytophages, such as occurred with the citrus psyllid D. citri during the second half of the year.
Therefore, the strategy of using alternate hosts for the conservation of natural enemies involves complex trophic relationships; for example, phytophagous-plants, natural enemies-phytophagous, between the same natural enemies, or, between natural enemy-phytophagous-plant. Therefore, a greater understanding of the trophic relationships between the organisms involved and their environment should be the objectives of future studies. For example, in addition to A. nerii, the other phytophages recorded in A. curassavica may also be important reservoirs of natural enemies, and the eggs of Danaus plexippus were important hosts of T. pretiossum (Cortez-Madrigal et al., 2014) and this parasitoid has been widely used in mass production for the control of lepidoptera pests (Hoffmann and Frodsham, 1993). The fact that the phytophagous species recorded in A. curassavica (Cortez-Madrigal et al., 2016) is not a crop pest (Blackman and Eastop, 1984; Cervantes-Peredo and Elizalde-Amelco, 2007) indicates the potential of the plant to be used as a refuge and increase of natural enemies for the management of pests in agroecosystems.
Therefore, the use of “companion” plants for the conservation of alternate hosts of natural enemies should be seen as one more strategy of integrated pest management, where the central objective should be to achieve the stability of the agroecosystem rather than the control of pests specific. In the cultivation of citrus fruits, for example, surveillance of the main pests must be maintained through routine sampling; and the management, both of the accompanying plant and citrus fruits (e.g. pruning). The foregoing will indicate the need for the use of other strategies for the control of emerging pests; for example, for D. citri would be the release of T. radiata or, in the last case, the use of low impact insecticides.
The inclusion of plants such as A. curassavica in crops would cover the important gap that exists in terms of biological diversity of highly technified agricultural systems, but poor in terms of diversity of natural enemies. Other studies have documented the importance of other Asclepias species, but only as a source of nectar for natural enemies (Fielder and Landis, 2007). The present study strengthens the idea that A. curassavica can be an important reservoir of natural enemies (Peña-Martínez et al., 2001) and evidence of its potential in the management of citrus pests is shown.
A. curassavica is a perennial plant, widely distributed (Juarez-Jaimes et al., 2007) and easy to reproduce, which facilitates its use as a reservoir of alternative hosts and a source of nectar for natural enemies. In addition to its presence in situ, A. curassavica could also be used to increase populations of A. nerii in greenhouse conditions, infest them with the parasitoid L. testaceipes and later transfer the pots with the “mummies” to the agroecosystems where there are aphid problems.
An advantage that has not been mentioned in the companion plants is its usefulness as “sentinel” plants in the agroecosystems this plant along with its phytophages, would indicate the presence or absence of a wide range of natural enemies of pests, which would eventually share with crops. Low populations or absence of natural enemies could be the signal to implement other control strategies; For example, the absence of coccinellids in the second half of the year would suggest greater surveillance of D. citri.
Conclusions
The presence of Asclepias curassavica in Persian lemon plantations showed a significant effect on the incidence of pests, its impact depended on the pest species and the time of year. The use of the plant should be visualized as part of an integrated pest management, where the central objective is the stability of the agroecosystem entomowildlife and not the biological control of specific pests.
Gratefulness
To the Secretariat of Research and Postgraduate of the National Polytechnic Institute for the financing for the realization of the study. To the National Council of Science and Technology (CONACYT) for the grant-thesis awarded to the first author.
REFERENCES
Arredondo, B. H. C. y Rodríguez del B. L.A . 2008. Casos de control biológico en México. Mundiprensa. 1ra . Ed. México, D. F. 423 p. [ Links ]
Bautista, M. N. 2006. Insectos plaga: una guía ilustrada para su identificación. Colegio de Postgraduados. 1ra . Ed. Montecillo, Texcoco, Estado de México, México. 113 p. [ Links ]
Blackman, R. L. and Eastop, V. F. 1984. Aphids on the world’s crops. An identification guide. John Wiley & Sons LTD. London, UK. 466 p. [ Links ]
Cervantes, P. L. y Elizalde, A. E. 2007. Estados de desarrollo y biología de tres especies de Lygaeinae (Hemiptera-Heteroptera: Lygaeoidea: Lygaeidae). México. Rev. Mex. Biod. 78(2):339-350. [ Links ]
Cortez, M. H.; García, G. F. y Guzmán, L. A. 2014. Conservando la mariposa monarca (Danaus plexippus L.), conservando enemigos naturales de plagas. México. Rev. Chapingo Ser. Ciencias Forestales y del Ambiente. 30(3):247-253. [ Links ]
Cortez, M. H.; García, G.; F.; Guzmán, L. A.; Acuña, S. J. A. and Otero, C. G. 2016. Conserving phytophagous arthropods to conserve natural enemies: Asclepias curassavica as the model. Southwestern Entomol. 41(3):681-692. [ Links ]
Cortez, M. E.; Lugo, A. N. E.; Pérez, M. J. y Apodaca, S. M. A. 2011. Primer reporte de enemigos naturales y parasitismo sobre Diaphorina citri Kuwayama en Sinaloa, México. Revista Científica UDO Agrícola. 11(1):97-103. [ Links ]
Ehler, L. E. 1998. Conservation biological control. In: conservation biological control. Barbosa, P. (Ed.). Academic Press. San Diego, CA, USA. 1-8 pp. [ Links ]
Faostat. 2016. Agriculture Organization of the United Nations Statistics Division (2014).http://faostat3.fao.org/browse/Q/QC/S. [ Links ]
Ferro, D. N. and McNeil, J. N. 1998. Habitat enhancement and conservation of natural enemies of insects. In: conservation biological control. Barbosa, P. (editor). Academic Press. San Diego, CA, USA. 123-132 pp. [ Links ]
Fielder, A. K. and Landis, D. A. 2007. Attractiveness of Michigan native plants to arthropod natural enemies and herbivores. EE. UU. Environ. Entomol. 36(4):751-765. [ Links ]
González, C. J. C.; Castellanos, S. I. E.; Fucikovsky, Z. L. J.; López, H. M. y Sánchez, R. G. 2012. Coccinelidos como potenciales enemigos naturales de Diaphorina citri (Hemiptera: Psyllidae) en huertos de cítricos de Tuxpan, Veracruz, México. Revista Científica UDO Agrícola. 12(4): 855-860. [ Links ]
Grafton, C. E.; Stelinski, L. L. and Stansly, P. A. 2013. Biology and management of Asian citrus psyllid, vector of the Huanglongbing pathogens. EE.UU. Annul Rev. Entomol. 58:413-432. [ Links ]
Hernández, G. C.; Curti, D. S. A.; Sandoval, R. J. A.; Uribe, B. A.; Loredo, S. R. X. y López, A. J. I. 2011. Fluctuación poblacional de enemigos naturales de Diaphorina citri en cinco regiones agroecológicas de Veracruz y Puebla. In: Memorias del 2do. Simposio Nacional sobre investigación para el manejo del psílido asiático de los cítricos y el Huanglongbing en México. López-Arrollo, J. I. y Gónzalez-Lauck, V. W. (Comp.). Colegio de Postgraduados. Montecillo, Estado de México, México. 228-240 pp. [ Links ]
Hoffmann, M. P. and Frodsham, A. C. 1993. Naturals enemies of vegetable insect pests. Cornell Cooperative Extension. Ithaca, NY, USA. 63 p. [ Links ]
Jacas, J. A. and Urbaneja, A. 2010. Biological control in citrus in Spain: from clasical to conservation biological control. In: Integrated management of arthropod pests and insect borne diseases. Ciancio, A. and Mukerji, K. G. (eds.). Springer, London, UK. 61-70 pp. [ Links ]
Juárez, J. V.; Alvarado, C. L. O. y Villaseñor, J. L. 2007. La familia Apocynaceaesensu latoen México: diversidad y distribución. México. Rev. Mex. Biod. 78(2):459-482. [ Links ]
Landis R, D. A. 2000. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Rev. Entomol. 45:175-201. [ Links ]
Marshall, S. A. 2012. Flies: the natural history and diversity of Diptera. Firefly Books. Buffalo, New York, USA. 616 p. [ Links ]
Peña, M. M. R.; Lomeli, J. R.; Trejo G., L. A. y Villegas, N. J. 2001. Monitoreo de afídos y afidófagos de importancia agrícola. Escuela Nacional de Ciencias Biológicas-Instituto Politécnico Nacional. México, D. F. 89 p. [ Links ]
Pimentel, D. and Edwards, C. A. 1982. Pesticides and ecosystems. Inglaterra. Biosc. 32(7):595-600. [ Links ]
Ruiz, C. E.; Coronado, B. J. M. y Myartseva, S. N. 2008. Psílido asiático de los cítricos Diaphorina citri (Hemiptera: Psyllidae). In: casos de control biológico en México. Arredondeo, B. H. C. y Rodríguez del, B. L. A. (Eds.). Mundiprensa. México, D. F. 323-331 pp. [ Links ]
Salvo, A. y Valladares, G. R. 2007. Parasitoides de minadores de hojas y manejo de plagas. Ciencia e Investigación Agraria. 34(3):167-185. [ Links ]
Sánchez, S. S. y Cortez, M. H. 1998. Presencia de Aphis craccivora Koch. (Homoptera: Aphididae) en plantaciones de cacao en Tabasco, México. Brasil. Agrotrópica. 10(1):47-48. [ Links ]
SAS Institute Inc. 1999. SAS Sofware for Windows Version 4.10. SAS Institute. Cary N. C, USA. [ Links ]
Valladares, G. R. and Salvo, A. 1999. Insect-plant food webs could provide new clues for Pest Management. Environ. Entomol. 28:539-544. [ Links ]
Van Driesche, R.; Hoddle, M.; Center, T. D.; Ruíz, C. E.; Coronada, B. J. y Manuel, A. J. 2007. Control de plagas y malezas por enemigas naturales (No. 632.96 V33). US Department of Agriculture, US Forest Service, Forest Health Technology Enterprise Team. 751 p. [ Links ]
Van Lenteren, J. C. 2007. Internet book of biological control. IOBC. 4th Edition. Wageningen, The Netherlands. 182 p. [ Links ]
Wyckhuys, K. A. G.; Lu Y.; Morales, H.; Vázquez, L. L.; Legaspi, J. C.; Eliopoulos, P. A. and Hernández, L. M. 2013. Current status and potential of conservation biological control for agricultura in the developing world. EE.UU. Biological Control. 65(1):152-167. [ Links ]
Received: February 00, 2018; Accepted: April 00, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
