











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Durante el proceso del desarrollo que conduce a la floración en las angiospermas, un ápice vegetativo sufre la transición hacia reproductivo (Salazar et al., 1999). Este cambio ocurre en respuesta a mensajes bioquímicos (Rocha et al., 2010) y moleculares (Pin y Nilsson, 2012) procedentes de hojas maduras. Al inicio del proceso de transición, algunas condiciones ambientales y/o prácticas de manejo del huerto, como una lluvia o riego abundante, temperatura cálida, poda, fertilización nitrogenada o biorreguladores vegetales pueden inhibir el proceso (Salazar et al., 2013). Una vez percibidos y procesados los estímulos ambientales para la transformación de la yema vegetativa en reproductiva, el meristemo apical de la yema del aguacate cambia su forma de cónica a aplanada (Salazar et al., 1998).
En cierto punto, después de la transición de la fase vegetativa a reproductiva el destino, tanto del ápice como de la yema no es reversible y no puede ser modificado con tratamientos que normalmente inhiben la floración. A este proceso se le conoce como determinación irreversible a la floración (DIF) (Salazar et al., 1999). El conocer cuándo ocurre la DIF en aguacate es importante para programar las diversas prácticas de manejo del huerto ya sea para incrementar la intensidad de la floración o para promover el crecimiento vegetativo y disminuir la floración (Salazar et al., 2006).
La defoliación y el anillado de brotes son técnicas de campo empleadas para determinar cuándo ocurre la DIF (Salazar et al., 2006). Ambas promueven el crecimiento de las yemas en los brotes tratados, independientemente si se encuentran en estado vegetativo o reproductivo al momento del tratamiento. Las hojas mantienen a las yemas en una condición de paradormancia. La defoliación remueve este factor de inhibición del crecimiento forzando al brote a un rápido desarrollo, de ahí su utilidad como herramienta con tratamientos donde varían las fechas (Bernier et al., 1981; Bernier, 1988).
Por su parte, el anillado aísla al brote del resto del árbol y evita que las yemas reciban mensajes bioquímicos de otras partes de la planta que pudieran estimular la floración. Si las yemas florecen, significa que al momento del tratamiento ya habían alcanzado la DIF y continuaron hacia la diferenciación floral (Salazar et al., 2006). Esta técnica ha sido empleada en los aguacates ‘Hass’ (Salazar et al., 2006; Dixon et al., 2006; Rocha et al., 2010), ‘Choquette’ y ‘Booth-8’ (Salazar et al., 2008).
En los últimos años, el aguacate ‘Méndez No. 1’ (Plant Patent 11,173 USA 2002), referido aquí como ‘Méndez’ ha adquirido importancia en el sur del estado de Jalisco en donde existen más de 11 000 ha. La importancia económica de ‘Méndez’ se debe a que una proporción importante de su cosecha anual madura más temprano (verano) que la de ‘Hass’ (otoño- invierno). El fruto cosechado en verano usualmente alcanza el precio más alto del año (Salazar et al., 2015).
En el sur de Jalisco el cv. Méndez comúnmente presenta dos flujos de crecimiento vegetativo (invierno y verano) y dos épocas de floración. La primera (principal) en otoño (septiembre) y la segunda en invierno (febrero). El conocer cuándo ocurre la DIF es importante para entender el desarrollo floral del aguacate y mejorar las prácticas de manejo de los huertos. El objetivo de esta investigación fue determinar la fecha, estado de desarrollo de las yemas y la temperatura asociada a la determinación irreversible a la floración en los dos principales flujos vegetativos del aguacate ‘Méndez’ en el sur de Jalisco, México.
Materiales y métodos
Características de los huertos
La investigación inició en febrero 2014 en dos huertos comerciales de aguacate ‘Méndez’ manejados con fertirriego, suelo Feozem háplico y clima Semicálido subhúmedo [AC(w)] (García, 1998) del sur del estado de Jalisco. Los huertos “Colorín 1” y “Ocote cuate 2” se encuentran a distanciamientos de 7×3.5 y 5×5 m, tienen cuatro y seis años de edad y se ubican a 1 556 y 1 428 m de altitud, respectivamente. En cada huerto se seleccionaron 10 árboles y en cada uno de ellos se marcaron 30 brotes de cada flujo vegetativo (invierno o verano) ocurridos en 2014 y 2015.
Anillado y defoliación de brotes
Los tratamientos se realizaron en 2014 y 2015 a intervalos mensuales e iniciaron en marzo y octubre para brotes de los flujos vegetativos de invierno y verano, respectivamente. Por cada año y huerto, en cada fecha de tratamiento (cinco y seis fechas para los flujos de verano e invierno, respectivamente) se anilló y defolió un brote por árbol, de un total de 10 árboles por huerto. La defoliación consistió en la eliminación manual de todas las hojas de los dos últimos flujos de crecimiento vegetativo. Para el anillado se removió una franja completa de corteza de 2 cm de ancho en la parte basal del penúltimo crecimiento. Los tratamientos se suspendieron cuando las yemas apicales de los brotes alcanzaron el estado de desarrollo E- 6 según la escala visual de Salazar et al. (1998). Por cada año, huerto y flujo vegetativo fueron considerados 25 brotes testigos los cuales no fueron alterados.
Estado de desarrollo floral
En cada fecha de tratamiento se colectó una yema apical por árbol de cada uno de los flujos vegetativos. Las yemas fueron fijadas en FAA (formaldehído:ácido acético:etanol, 5:5:90, v:v:v) y después introducidas a una campana de vacío (Nalgene 8040317, Nalgen Company) a 30 KPa por 5 h. Posteriormente fueron clasificadas bajo un microscopio estereoscópico (Zeiss Stereomikroskop Mod. Stemi 2000-C, Carl Zeiss, Göttingen, Germany), con los estados (E) de desarrollo de la escala visual de Salazar-García et al. (1998) la cual comprende de E-1 (yema vegetativa) a E-11 (antesis). En total, para cada año, por todas las fechas de tratamiento fueron colectadas 60 y 50 yemas de los flujos de invierno y verano, respectivamente.
Una vez identificada la fecha en que ocurrió la DIF se realizó la caracterización anatómica de las yemas que alcanzaron este estado de desarrollo. Las yemas se extrajeron del FAA y se lavaron con agua corriente y luego se deshidrataron con alcohol y xileno. Posteriormente, fueron incluidas en parafina (Ruzin, 1999). Se obtuvieron secciones anatómicas longitudinales con un micrótomo rotatorio (HM 350S, Walldorf, Germany) y grosor de 8 µm, se montaron en portaobjetos y tiñeron con safranina y verde fijo (Ruzin, 1999). Al final se les aplicó bálsamo de Canadá y fueron protegidas con cubreobjetos. Las secciones fueron fotografiadas con una cámara digital (Canon modelo Power Shot G11. NY. USA) montada en un microscopio óptico (Zeiss HBO 50/AC, Carl Zeiss, Göttingen, Germany).
Tipo de crecimiento producido
El tipo de crecimiento (brotes florales, vegetativos o inactivos) producido por las yemas apicales tanto de los brotes tratados como del testigo se cuantificó al término de cada periodo de floración. Para los flujos vegetativos de invierno y verano 2014 las evaluaciones fueron en octubre 2014 y marzo 2015, respectivamente. En el caso de brotes de los flujos de invierno y verano 2015, la evaluación se realizó en noviembre 2015 y abril 2016, respectivamente.
Cálculo de horas frío (HF)
En cada huerto fue registrada cada hora la temperatura del aire. Se utilizaron registradores automatizados HOBO H8 (Onset Computer, Witzprod, Englewood Cliffs, NJ, USA) operados con baterías. De forma independiente para brotes de los flujos vegetativos de invierno y verano, se cuantificó la ocurrencia de temperaturas mínimas, desde 8 a 20 °C, en incrementos de 1 °C que pudieran asociarse con la fecha de la DIF. Se empleó la fórmula: HF = (Tmín T,1,0); donde: Tmín= temperatura mínima registrada cada hora; T= temperatura crítica, de 8 a 20 °C. Si se cumple la condición de temperatura, entonces el valor de HF es 1, en caso contrario es 0. Mediante el gestor de bases de datos Microsoft Access Versión 14 se sumaron los registros horarios de temperatura organizados desde 8 a 20 °C, en incrementos de 1 °C. Los valores de HF8 hasta HF20, se acumularon para cada periodo de defoliación y anillado.
A estos valores se les denominó horas frío acumuladas (HFA). Para los flujos vegetativos de invierno y verano se consideró como día cero cuando las yemas apicales estaban en E-1 (etapa vegetativa), lo que ocurrió en febrero y agosto, respectivamente.
Identificación de la temperatura asociada con la DIF
Las HFA se usaron como variables independientes y el desarrollo floral de yemas apicales en brotes de cada flujo como variable dependiente. Posteriormente, se usó el procedimiento Stepwise SAS/STAT (SAS Institute Inc., 9.2.) para seleccionar el mejor modelo por orden de respuesta (segundo hasta quinto orden) para cada temperatura crítica (a partir de ≤ 8 °C hasta ≤ 20 °C, en intervalos de 1 °C). Los criterios para elegir los mejores modelos fueron: 1) valor de R2; 2) el menor cuadrado medio del error (CME); y 3) el valor de Cp, sugerido por Mellows (Draper y Smith, 1981).
Obtención de valores de los coeficientes matemáticos.
Una vez identificados los mejores modelos, se calcularon sus coeficientes matemáticos (B0 ,......., Bn ) por el procedimiento REG utilizando las HFA, a partir de ≤ 8 °C hasta ≤ 20 °C, en intervalos de 1 °C.
Validación de los modelos de predicción
Para los brotes de cada flujo vegetativo se evaluó la capacidad para predecir la DIF de los mejores modelos de predicción obtenidos en el año 1 contra el mismo flujo vegetativo en el año 2, y viceversa. Los valores de desarrollo floral predichos fueron analizados mediante una regresión contra los valores observados del desarrollo floral del año y flujo vegetativo correspondiente, utilizando el programa Excel (2010). Los criterios para determinar si los valores de los dos años pertenecían a una sola población fueron: 1) que la ordenada al origen de la regresión fuera lo más cercana a uno (B0= 1); 2), que la pendiente fuera lo más aproximada a uno (B1= 1); y 3) el mayor valor del coeficiente del modelo ajustado (R2). Este procedimiento sirvió para depurar modelos y encontrar el mejor.
Generación de un modelo único para predecir la DIF en cada flujo vegetativo
Posterior a la verificación de la no diferencia entre años, se obtuvo un modelo de regresión integrando la información de los dos años en un solo conjunto de datos, para cada flujo vegetativo.
Análisis de la información
Para el tipo de crecimiento producido por los brotes se utilizó un diseño experimental completamente al azar, con diferente número de tratamientos (fechas de defoliación y anillado) y 10 repeticiones (brotes). Para los brotes de invierno, el análisis estadístico fue hecho como factorial de la forma 2×2×7 (años × huertos × tratamientos) y para los brotes de verano se utilizó un factorial 2×2×6 (años × huertos × tratamientos). Previo a su análisis, los valores expresados en porcentaje fueron transformados mediante el arcoseno X+0.5 (Steel y Torrie, 1984), aunque en los resultados se presentan los valores reales. La comparación de medias fue hecha con la prueba de comparación múltiple de Waller-Duncan, p≤ 0.05.
Resultados y discusión
La información disponible sobre defoliación y anillado (solos o combinados) de brotes para realizar estudios de desarrollo floral inicial en frutales leñosos de clima cálido es escasa. Los trabajos relacionados con este tema son de Erner (1988) en cítricos, Pérez et al. (2006) en mango y Salazar et al. (2006): Dixon et al. (2016) en aguacate ‘Hass’. Todos ellos han sido efectivos en determinar el tiempo en que ocurre la DIF.
Tipo de crecimiento producido en respuesta a los tratamientos
Brotes de invierno
No se detectaron diferencias en la producción de inflorescencias entre los dos años de estudio o entre los dos huertos experimentales, aunque sí se detectaron para las fechas de tratamiento. Tampoco hubo diferencias en la producción de brotes vegetativos entre los años 2014 y 2015, aunque sí para huertos y fechas de tratamiento (Cuadro 1).
Cuadro 1 Análisis de la varianza para el tipo de crecimiento producido por brotes del flujo de invierno 2014 y 2015 que fueron defoliados y anillados en aguacate ‘Méndez’. Conjunto de datos de los huertos Colorín 1 y Ocote cuate 2.
| Variable | Fuente | GL | SC | CM | F | Pr> F |
| Inflorescencias | Años | 1 | 5.46 | 5.46 | 0.92 | 0.3396 |
| Huertos | 1 | 21.47 | 21.47 | 3.63 | 0.0605 | |
| Tratamientos | 6 | 468.6 | 78.1 | 13.2 | 0.0001 | |
| Huertos* años | 1 | 0.016 | 0.016 | 0 | 0.9584 | |
| Año*tratamiento | 6 | 18.75 | 3.12 | 0.53 | 0.7854 | |
| Vegetativos | Años | 1 | 18.57 | 18.57 | 3.29 | 0.0734 |
| Huertos | 1 | 26.87 | 26.87 | 4.77 | 0.032 | |
| Tratamientos | 6 | 139.27 | 23.21 | 4.12 | 0.0012 | |
| Años* huertos | 1 | 5.49 | 5.49 | 0.97 | 0.3265 | |
| Año* tratamiento | 6 | 40.33 | 6.72 | 1.19 | 0.3195 | |
| Inactivos | Años | 1 | 0.35 | 0.35 | 0.05 | 0.8322 |
| Huertos | 1 | 0.28 | 0.28 | 0.04 | 0.8489 | |
| Tratamientos | 6 | 143.52 | 23.92 | 3.02 | 0.0105 | |
| Años* huertos | 1 | 47.41 | 47.46 | 5.99 | 0.0166 | |
| Año* tratamiento | 6 | 7.93 | 1.32 | 0.17 | 0.9848 | |
Fechas de evaluación: 28 de octubre de 2014 y 26 de noviembre de 2015.
El tipo de crecimiento producido por los brotes de invierno (datos de ambos huertos) no fue afectado por el año de estudio (Cuadro 1). Al comparar entre huertos, sólo hubo diferencias en la producción de brotes vegetativos y el huerto Ocote cuate 2 presentó mayor proporción (66.1%) que el Colorín 1 (53%) (Cuadro 2).
Cuadro 2 Efecto de huerto y año sobre el tipo de crecimiento producido por brotes del flujo de invierno 2014 y 2015 en aguacate ‘Méndez’. Huertos Colorín 1 y Ocote cuate 2.
| Años | Nz | Tipo de crecimiento producido por brotes de invierno (%) | ||
| Inflorescencia de verano | Vegetativo | Inactivo | ||
| 2014 | 170 | 18.8 ay | 65 a | 16.2 a |
| 2015 | 170 | 28.1 a | 53.9 a | 18 a |
| Pr>F | 0.3396 | 0.0734 | 0.8322 | |
| Huertos: | ||||
| Colorín 1 | 170 | 29.8 a | 53 b | 17.2 a |
| Ocote cuate 2 | 170 | 17.3 a | 66.1 a | 16.6 a |
| Pr>F | 0.0605 | 0.032 | 0.8489 | |
Fechas de evaluación: 28 de octubre de 2014 y 26 de noviembre de 2015; z= brotes evaluados, 10 brotes tratados por fecha × 6 fechas × 2 huertos, incluye 25 brotes del testigo por huerto (50 total); y= comparación de medias en las columnas para años o huertos por Waller-Duncan, p≤ 0.05.
La respuesta a las fechas en que se hizo la defoliación y anillado indicó que a partir del 28 de mayo 5.6% de yemas alcanzaron la DIF ya que produjeron inflorescencias. Sin embargo, para el 26 de junio, esta proporción se incrementó a 41%, superando al testigo (22.2%) (Cuadro 3).
Cuadro 3 Efecto de la fecha de defoliación y anillado sobre el tipo de crecimiento producido por brotes del flujo de invierno 2014 y 2015 en aguacate ‘Méndez’. Conjunto de datos de los huertos Colorín 1 y Ocote cuate 2.
| Fecha de tratamiento (2014 y 2015) |
Nz | DYy | Tipo de crecimiento producido por brotes de invierno (%) | |||
| Inflorescencias de verano | Vegetativo | Inactivo | ||||
| 28 marzo | 40 | 2.2 | 0 cx | 90.3 a | 9.72 ab | |
| 28 abril | 40 | 3.3 | 0 c | 76.4 a | 23.6 ab | |
| 28 mayo | 40 | 3.5 | 5.6 bc | 73.6 ab | 20.8 ab | |
| 26 junio | 40 | 3.7 | 41 a | 53.5 bc | 5.5 b | |
| 28 julio | 40 | 5.5 | 49.4 a | 39.6 c | 11.1 ab | |
| 28 agosto | 40 | 6.4 | 39.6 a | 52.1 bc | 8.3 ab | |
| Testigo | 100 | 22.2 b | 47.8 c | 30 a | ||
| Pr> F | 0.0001 | 0.0012 | 0.0105 | |||
Fechas de evaluación: 28 de octubre de 2014 y 26 de noviembre de 2015; z= número de brotes evaluados, 10 brotes tratados en cada fecha de tratamiento × 2 años × 2 huertos. Incluye 25 brotes del testigo × 2 huertos × 2 años; y= mediana de las etapas de desarrollo floral según la escala visual de Salazar-García et al. (1998); x= comparación de medias en las columnas por Waller-Duncan, p≤ 0.05.
Para el tratamiento aplicado el 26 de junio la mediana del estado de desarrollo floral fue E- 3.7. Esto significa que la mayor población de yemas de los brotes de invierno alcanzó la DIF para el 26 de junio con un estado de desarrollo entre E-3 y E-4, según la escala de Salazar et al. (1998). Las yemas en E-3 presentan las siguientes características externas: yema apical cerrada y senescencia parcial de la escama de la yema. En el caso del E-4, sus características son: las escamas de las yemas se han separado y las brácteas de la inflorescencia se expanden hacia todos los lados de la yema. El E-3 ha sido asociado con la transición de la fase vegetativa a la reproductiva en aguacate ‘Hass’ en California (Salazar et al., 1998).
Brotes de verano
En brotes de verano el análisis factorial para la producción de inflorescencias mostró diferencias entre huertos, entre tratamientos y en la interacción huertos*años (Cuadro 4). Algo similar ocurrió para la producción de brotes vegetativos, aunque en este caso sólo hubo diferencias significativas entre huertos y entre tratamientos. La presencia de brotes inactivos resultó significativa para tratamientos y para la interacción año*tratamiento.
Cuadro 4 Análisis de varianza para el tipo de crecimiento producido por brotes del flujo de verano 2014 y 2015 que fueron defoliados y anillados en árboles de aguacate ‘Méndez’. Conjunto de datos de los huertos Colorín 1 y Ocote cuate 2.
| Variable analizada | Fuente | GL | SC | CM | F | Pr>F |
| Inflorescencias | Huertos | 1 | 33.43 | 33.43 | 7.53 | 0.0074 |
| Tratamientos | 5 | 525.88 | 105.18 | 23.68 | 0.0001 | |
| Huertos*años | 1 | 36.81 | 36.81 | 8.29 | 0.0052 | |
| Año*tratamiento | 5 | 3.29 | 0.66 | 0.15 | 0.98 | |
| Vegetativos | Años | 1 | 7.92 | 7.92 | 1.34 | 0.2505 |
| Huertos | 1 | 23.93 | 23.93 | 4.05 | 0.0478 | |
| Tratamientos | 5 | 142.71 | 28.54 | 4.83 | 0.0007 | |
| Años*huertos | 1 | 0.75 | 0.75 | 0.13 | 0.7227 | |
| Año*tratamiento | 5 | 46.65 | 9.33 | 1.58 | 0.1765 | |
| Inactivos | Años | 1 | 18.04 | 18.04 | 2.81 | 0.0978 |
| Huertos | 1 | 0.02 | 0.02 | 0 | 0.9532 | |
| Tratamientos | 5 | 80.32 | 16.06 | 2.5 | 0.0378 | |
| Años*huertos | 1 | 13.58 | 13.58 | 2.12 | 0.15 | |
| Año*tratamiento | 5 | 99.15 | 19.83 | 3.09 | 0.0138 | |
Fechas de evaluación: 25 de marzo de 2015 y 13 de abril de 2016.
En el Colorín 1 se produjo mayor cantidad de inflorescencias (35%) y menor de brotes vegetativos (43.7%) que en Ocote cuate 2 (24.4 y 54%, respectivamente) (Cuadro 5). Respecto al efecto de los tratamientos para identificar la fecha de la DIF, los brotes del flujo vegetativo de verano 2014 y 2015 tratados en septiembre y octubre, no produjeron inflorescencias (Cuadro 6) y más de 68% de brotes vegetativos. La producción de inflorescencias (25.7%) por los brotes tratados se presentó a cuando la defoliación y anillado se hicieron a partir del 26 noviembre. Para esta fecha, la mediana del estado de desarrollo de las yemas fue E-3.2.
Cuadro 5 Efecto de huerto y años sobre el tipo de crecimiento producido por brotes del flujo de verano 2014 y 2015 en aguacate ‘Méndez’. Huertos Colorín 1 y Ocote cuate 2.
| Años | Nz | Tipo de crecimiento producido por brotes de verano (%) | ||
| Inflorescencias de invierno | Vegetativo | Inactivo | ||
| 014 | 150 | 29 | 46.8 | 24.1 |
| 015 | 150 | 29.9 | 51 | 19 |
| r>F | 0.6274 | 0.2505 | 0.0978 | |
| Huertos: | 43.7 b | 21.3 | ||
| Colorín 1 | 150 | 35.0 ay | ||
| Ocote cuate 2 | 150 | 24.4 b | 54 a | 21.6 |
| r>F | 0.0074 | 0.0478 | 0.9532 | |
z= brotes evaluados, 10 brotes tratados por fecha × 5 fechas × 2 huertos, incluye 25 brotes del testigo por huerto (50 total); y= comparación de medias en las columnas para años o huertos por Waller-Duncan, p≤ 0.05.
Cuadro 6 Efecto de la fecha de defoliación y anillado sobre el tipo de crecimiento producido por brotes del flujo de verano 2014 y 2015 en aguacate ‘Méndez’. Conjunto de datos de los huertos Colorín 1 y Ocote cuate 2.
| Fecha de tratamiento (2015 y 2016) |
Nz | DYy | Tipo de crecimiento producido por brotes de verano (%) | ||
| Inflorescencias de invierno | Vegetativo | Inactivo | |||
| 26 septiembre | 40 | 2.1 | 0 dx | 68.1 ab | 31.9 a |
| 26 octubre | 40 | 2.8 | 0 d | 76.4 a | 23.6 ab |
| 26 noviembre | 40 | 3.2 | 25.7 c | 43.8 cd | 30.6 a |
| 26 diciembre | 40 | 5.6 | 31.9 bc | 56.3 bc | 11.8 b |
| 26 enero | 40 | 6.2 | 54.9 a | 35.4 d | 9.7 b |
| Testigo | 100 | 45.1 b | 33.5 d | 21.4 ab | |
| Pr>F | 0.0001 | 0.0007 | 0.0378 | ||
Fecha de evaluación: 25 de marzo de 2015 y 13 de abril 2016; z= número de brotes evaluados, 10 brotes tratados en cada fecha de tratamiento × 2 años × 2 huertos; incluye 25 brotes del testigo × 2 huertos × 2 años; y= mediana de las etapas de desarrollo floral según la escala visual de Salazar-García et al. (1998); x= comparación de medias en las columnas por Waller-Duncan, p≤ 0.05.
Como ya se mencionó, en el cv. Méndez la DIF ocurrió en dos épocas, según el tipo de brote. En brotes del flujo vegetativo de invierno ocurrió a partir del 26 de junio y en brotes de verano a partir del 26 de noviembre. Lo anterior difiere de lo encontrado en el clima cálido del estado de Nayarit, México en donde los brotes de primavera e invierno del cv. Choquette alcanzaron la DIF en la misma fecha (15 noviembre) (Salazar et al., 2008); en esta misma región, tanto los brotes de invierno como de verano del cv. Hass alcanzaron la DIF el 15 de octubre (Salazar et al., 2006) y en el cv. Booth 8 la DIF en brotes de verano ocurrió para el 30 de noviembre. En Nueva Zelanda los brotes de ‘Hass’ emergidos en primavera (otoño, para el hemisferio norte) alcanzaron la DIF en otoño (primavera para el hemisferio norte) (Dixon et al., 2006). Lo anterior, evidencia la influencia de las condiciones ambientales propias de cada región productora sobre la fenología de cada cultivar de aguacate, en este caso, la época en que ocurre la DIF.
Características microscópicas de las yemas apicales determinadas irreversiblemente a floración
Las yemas colectadas de ‘Méndez’ para ambos flujos vegetativos macroscópicamente lucieron más desarrolladas que las de ‘Hass’ descritas para California (Salazar et al., 1998) y Nayarit (Salazar et al., 2007), de tal forma que parecían E-4. Sin embargo, microscópicamente se confirmó que correspondían a E-3, cuyas características son: meristemo del eje primario en forma convexa, presencia de meristemos de ejes secundarios de la inflorescencia y de las escamas que cubren la yema (Figura 1). De acuerdo a esto, las yemas clasificadas inicialmente como E-4 fueron reclasificadas a E-3. El estado de desarrollo E-3 ya ha sido asociado con la DIF en el aguacate ‘Hass’ (Salazar et al., 1999).

Efecto de la temperatura ambiental sobre la DIF
De todas las temperaturas base evaluadas (≤8 °C hasta ≤20 °C), las temperaturas ≤18 e ≤20 ºC fueron las que mejor se asociaron con la DIF de yemas en brotes de los flujos vegetativos de verano e invierno, respectivamente (Cuadro 7). El resultado encontrado para yemas del flujo de invierno (≤20 ºC) difiere de lo reportado por Salazar et al. (1999) en donde los árboles jóvenes de ‘Hass’ mantenidos a temperatura constante de 25/20°C (día/noche) en cámaras de crecimiento no florecieron. Las temperaturas ambientales 25/20 °C no ocurren de forma constante a nivel comercial ya que hay fluctuaciones en ambos sentidos que permiten el cultivo exitoso de ‘Hass’ en diferentes regiones productoras (Salazar et al., 2013).
Cuadro 7 Coeficientes matemáticos y criterios de selección los mejores modelos de predicción de la DIF, basados en temperatura, en brotes de invierno 2014 y 2015 (que florecieron en verano 2014 y 2015) y verano 2014 y 2015 (que florecieron en invierno 2015 y 2016) en aguacate ‘Méndez’.
| Temperaturas (HFA) | Parámetros | Criterios | ||||||||
| Bo | B1 | B2 | B3 | B4 | B5 | B6 | R2 | Cp | CME | |
| Brotes de invierno 2014 | ||||||||||
| ≤19 °C | 1.1 | -9.06E-04 | 1.22E-05 | -1.25E-08 | 3.84E-12 | 0.99 | 3.99 | 0.01 | ||
| ≤20 °C | 1.1 | -5.16E-04 | 9.85E-06 | -9.47E-09 | 2.70E-12 | 0.99 | 4.02 | 0.03 | ||
| ≤21 °C | 1.1 | -0.0002956 | 0.00000606 | -7.34E-12 | 4.95E-15 | -9.42E-19 | 0.99 | 4.03 | 0.03 | |
| Brotes de invierno 2015 | ||||||||||
| ≤19 °C | 1.31 | 6.55E-04 | 2.11E-08 | -3.86E-11 | 2.40E-14 | -4.93E-18 | 0.96 | 3.95 | 0.11 | |
| ≤20 °C | 1.3 | -8.95E-04 | 8.82E-06 | -1.43E-11 | 1.08E-14 | -2.29E-18 | 0.97 | 4.17 | 0.08 | |
| ≤21 °C | 1.30451 | -0.00054172 | 0.00000689 | -9.49E-12 | 6.56E-15 | -1.27E-18 | 0.98 | 4.1 | 0.05 | |
| Brotes de verano 2014 | ||||||||||
| ≤17°C | 1.04489 | 0.02457 | -0.00011901 | 1.76E-07 | -1.38E-13 | 6.08E-17 | 0.7 | 6 | 1.58 | |
| ≤18 °C | 1.18247 | 0.00191 | 2.92E-08 | -7.60E-11 | 6.37E-14 | -1.72E-17 | 0.84 | 4.02 | 0.69 | |
| ≤19 °C | 1.11505 | 0.0006 | 0.00001031 | -1.42E-08 | 8.08E-15 | -2.97E-18 | 0.9 | 4.09 | 0.41 | |
| Brotes de verano 2015 | ||||||||||
| ≤17°C | 1.29 | 7.32E-03 | -1.49E-05 | 3.80E-11 | -3.93E-14 | 1.16E-17 | 0.87 | 4.31 | 1.05 | |
| ≤18 °C | 1.3 | 5.70E-04 | 2.92E-08 | -5.96E-11 | 4.16E-14 | -9.63E-18 | 0.76 | 4.08 | 1.6 | |
| ≤19 °C | 1.22 | 3.36E-03 | -4.95E-12 | 4.86E-15 | -1.23E-18 | 0.85 | 4.05 | 0.99 | ||
Como la DIF de la población de brotes muestreados en los árboles experimentales se alcanzó cuando las yemas estaban entre E-3 y E-4, con fines estadísticos se consideró E-4 como el estado donde la DIF había ocurrido. En brotes del flujo de invierno desde E-1 (febrero) hasta E-4 (26 junio) se acumularon (promedio de dos años) 1480 HF con temperaturas ≤20 °C. Para brotes de verano desde E-1 (agosto) a E-4 (26 noviembre) se acumularon (promedio de dos años) 1 266 HF con temperaturas ≤18 °C.
La diferencia en la temperatura crítica y los requerimientos de HF para alcanzar la DIF entre los brotes de ambos flujos vegetativos puede deberse a que los brotes de invierno desarrollaron bajo las temperaturas cálidas de primavera; lo contrario ocurrió para brotes de verano, los que experimentaron el descenso de la temperatura ambiental típica del verano- otoño en la región donde se realizó el estudio. Los resultados confirman que el descenso de las temperaturas, más que un valor específico de la misma, es la que estimuló la transición de las yemas hacia la fase reproductiva a expensas del crecimiento vegetativo.
En un estudio en cámaras de crecimiento se sometieron árboles jóvenes de aguacate ‘Hass’ a cuatro semanas a 10/7 °C (día/noche) más cuatro semanas a 20/15 °C. La DIF que resultó en intensa floración se alcanzó con 1344 HF (Salazar et al., 1999). Estas HF son muy cercanas a lo registrado en condiciones de campo en el presente estudio, 1 480 HF y 1 239.5 HF en brotes de invierno y verano, respectivamente.
Verificación de la no diferencia entre datos del año 1 (2014) vs. año 2 (2015)
Para brotes de invierno el modelo HFA ≤20 °C del año 2 mostró una alta capacidad de predicción de la DIF para el año 1 (R2= 0.98; Figura 2A). Lo mismo sucedió al usar el modelo del año 1 vs. el año 2 (R2= 0.98; Figura 2B). Esta prueba también fue aplicada en brotes de verano con el modelo HFA ≤18 °C y resultó en una R2= 0.96 (año 2 vs. año 1) y R2= 0.98 (año 2 vs. año 1) (Figuras 2C y 2D).

Nuevos modelos generados utilizando datos de dos años
Ante la no diferencia entre años de la capacidad predictiva de los modelos para brotes de los flujos de invierno o verano se fusionaron los datos de los dos años y se obtuvo un nuevo modelo de predicción para cada tipo de brote (Cuadro 8). Los modelos únicos para brotes de invierno HFA ≤20 °C y verano HFA ≤18 °C fueron probados contra los datos de desarrollo floral reales obtenidos en 2014-2015 y su R2 fue superior a 0.98 (Figura 3)
Cuadro 8 Coeficientes matemáticos de los mejores modelos de predicción de la DIF para brotes de invierno o verano en aguacate ‘Méndez’ empleando el conjunto de datos de 2014 y 2015.
| Temperaturas | Brotes de invierno | Criterios | |||||||||
| Bo | B1 | B2 | B3 | B4 | B5 | B6 | R2 | Cp | CME | ||
| ≤20 °C | 1.20360 | 0.0007 | 1.68E-08 | -2.83E-11 | 1.63E-14 | -3.12E-18 | 0.962 | 5.23 | 0.102 | ||
| Brotes de verano | |||||||||||
| ≤18 °C | 1.10634 | 0.0001 | 0.00001376 | -1.83E-08 | 9.71E-15 | -3.43E-18 | 0.851 | 4.14 | 0.516 | ||
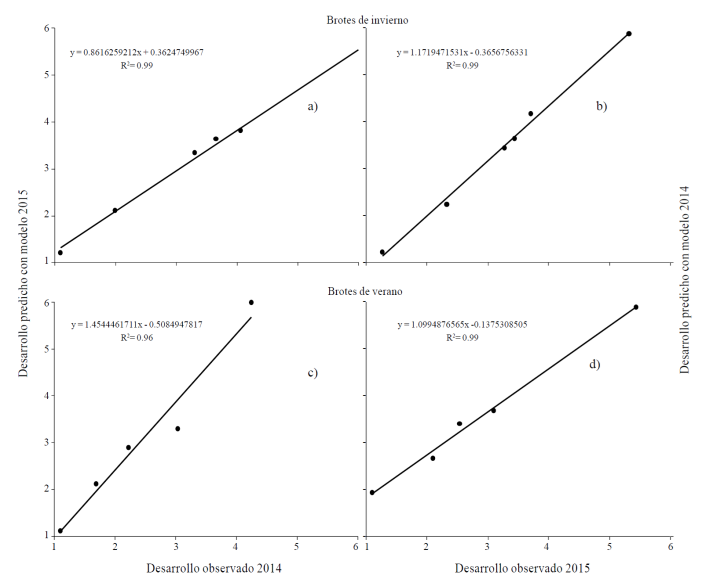
Figura 3 Ajuste de valores entre el estado de desarrollo floral observado en el año 1 (2014) y año 2 (2015) vs. el desarrollo floral predicho obtenido con el conjunto de datos de los años 2014-2015 en brotes del flujo de invierno con los modelos de HFA ≤20 (a y b) y de verano ≤18 °C (c y d).
Estos nuevos modelos generados podrán ser empleados para desarrollar una aplicación en Internet que muestre cuando los brotes de cada flujo vegetativo alcanzarán la DIF y programar de una manera segura algunas actividades en el huerto que usualmente inhiben o disminuyen la intensidad de floración, como la poda, la fertilización nitrogenada y de la aplicación de biorreguladores vegetales, entre otras, las cuales han mostrado ser efectivas para controlar el tamaño del árbol, para reducir la caída de fruto, para acelerar la madurez fisiológica e incrementar la producción y tamaño de fruto.
Conclusiones
En esta investigación se confirmó que el descenso de las temperaturas, más que un valor específico de la misma, es la que estimula la transición de las yemas hacia la fase reproductiva. Las diferencias en las épocas en que ocurre la determinación hacia la floración en los brotes originados por distintos flujos vegetativos son debidas a las variaciones estacionales de la temperatura. También, se demostró que en los aguacates ‘Hass’ y ‘Méndez’ las yemas determinadas irreversiblemente hacia floración presentan un estado de desarrollo anatómico similar, independiente de las diferencias geográficas y climáticas.

