










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.8 no.4 Texcoco Jun./Jul. 2017
https://doi.org/10.29312/remexca.v8i4.10
Articles
Morphological characterization of manzano pepper hybrids
1Centro Universitario Tenancingo-Universidad Autónoma del Estado de México. Carretera Santa Ana-Villa Guerrero, km 1.5. Tenancingo, Estado de México. CP. 52400. (ayala-brenba@hotmail.com; pa-che-ca@hotmail.com; lmvazquezg@uaemex.mx).
2Facultad de Ciencias Agrícolas-Universidad Autónoma del Estado de México. El Cerrillo Piedras Blancas, San Cayetano Morelos, Toluca, Estado de México. CP. 50000. ( m-rubi65@yahoo.com.mx).
The manzano pepper (Capsicum pubescens R and P) cultivated in central México shows genetic variability that can be used in breeding programs. The purpose of this research was to evaluate two hybridization methods and to morphologically characterize F1 descendants of crosses between Manzano pepper morphotypes. The experiment was carried out from 2012 to 2013 in greenhouse with the parents identified as white flower (FB), purple flower-long internode (FML) and purple flower-short internode (FMC). They were hybridized by the “with corolla” and “without corolla” methods and the F1 was evaluated for fruit picking and morphological characterization. The results indicated that the protection of the hybridized flower affected fruit picking by more than 50%. There were no significant differences between direct or reciprocal crosses (t; p= 0.42), suggesting no maternal effects. The position of the style in both progenitors and hybrids was heteromorphic, with 70-90% at the anther level. The variants of the characters stem color, corolla color, filament color and style color were found in the FB progenitor and were recessive to the corresponding FML and FMC. The low plant size and smaller leaves present in the FB and FMC progenitors were dominant in the hybrids and negatively and significantly affected fruit yield and heterosis. The results suggest additional strategies for genetic improvement in hybrids such as selection of segregants and backcrosses that would improve fruit size.
Keywords: Capsicum pubescens; hybridization; genetic variation; morphotypes
El chile manzano (Capsicum pubescens R y P) cultivado en el centro de México presenta variabilidad genética aprovechable en programas de mejoramiento genético. El objetivo de la investigación fue evaluar dos métodos de hibridación y caracterizar morfológicamente los descendientes F1 de las cruzas entre morfotipos de chile manzano. El experimento se desarrolló de 2012 a 2013 en invernadero con los progenitores identificados como flor blanca (FB), Flor morada-entrenudo largo (FML) y flor morada-entrenudo corto (FMC). Se hibridaron por los métodos “sin corola y “con corola” y la F1 se evaluó a prendimiento de fruto y caracterización morfológica. Los resultados indicaron que la protección de la flor hibridada afectó el prendimiento de fruto en más del 50%. No hubo diferencias significativas entre cruzas directas ni recíprocas (t; p= 0.42), lo que sugiere ausencia de efectos maternos. La posición del estilo tanto en progenitores como en los híbridos fue heteromórfica, con 70-90% al nivel de las anteras. Los variantes de los caracteres color del tallo, color de la corola, color del filamento y color del estilo se encontraron en el progenitor FB y fueron recesivos a los correspondientes de FML y FMC. El porte bajo de la planta y hojas de menor tamaño presentes en los progenitores FB y FMC fueron dominantes en los híbridos y afectaron negativa y significativamente en el rendimiento de frutos y heterosis. Los resultados sugieren estrategias de mejoramiento genético adicionales en híbridos como selección de segregantes y retrocruzas que mejorar el tamaño del fruto.
Palabras claves: Capsicum pubescens; hibridización; morpotipos; variación genética
Introduction
The Manzano pepper (Capsicum pubescens R and P) with the potential to be a viable alternative for the economic development of regions like the south of México state, has not reached the importance of crops like the floricultural ones by factors like the absence of an adequate technological management of its crop and fruits of heterogeneous quality; the latter being mainly affected by the existing genetic variability of native cultivated materials, which, contrary to the commercial disadvantage, represent a large genetic reservoir that can be used to select different qualitative and quantitative attributes of agronomic importance in this species (Pickersgill, 1997).
In the genus Capsicum there is a wide diversity among species, as well as variability within them for foliage and fruit characters (Bosland and Vostava, 2000), which is not necessarily related to the origin centers (Harlan, 1975). Therefore, the observation and variability recording create the basis for the selection of materials and techniques for genetic improvement, such as hybridization, that allow its exploitation (Sahagun, 1999). Broadly, hybridization accurately specifies the crosses of individuals from populations distinguishable from each other in one or more heritable characters (Harrison, 1990) and is a suitable tool for the analysis of character segregation and includes effects such as intraspecific genetic variation, origin and transfer of genetic variations, the origin of new ecotypes or species and the reinforcement or rupture of reproductive barriers.
The wide diversity of micro climates in México favors the existence of variation within species such as in southern Mexico state, where different C. pubecens morphotypes are cultivated, which represent a valuable reservoir of germplasm that could be useful in the improvement of this species. The aim of this research was to develop hybrids of direct and reciprocal crosses of three manzano pepper morphotypes (Capsicum pubescens R and P) contrasting in different characters and to evaluate them for different qualitative and quantitative attributes.
Materials and methods
The experiment was carried out from 2012 to 2013 in greenhouses of the UAEM Tenancingo University Center located at 18º 58՚ 2” north latitude and 99º 36՚ 44” west longitude and at an altitude of 2 064 m, in Tenancingo, Mexico state. The biological material consisted of three collections from the southern region of México state contrasting with different morphological characters and identified as white flower short internodes (FB); purple flower long internodes (FML) and purple flower short internodes (FMC).
Seeds extracted from fruits at physiological maturity of each morphotype were planted in a sieved substrate based on oak leaves in 200-well polystyrene germinators. The substrate was adjusted to pH 5.5 with agricultural lime at the rate of 600 g m-3. As background fertilization, simple calcium superphosphate (19.5%) was used at the rate of 1 kg m-3. The trays were placed in greenhouse and under shade mesh of 50% with relative humidity between 60% and 90%. Initial transplantation to 250 ml polystyrene cups was performed when plants showed 10 true leaves (40 days after sowing).
The final transplant was done in 600-gauge plastic bags of 60 L capacity when the plants showed the first bifurcation (80 days after sowing). Planting density was 1.5 plants m-2. In both transplants the substrate was a mixture of sandy soil, tepojal and organic matter, in a ratio of 5: 4: 1 respectively. The pH was adjusted to 5.5 with agricultural lime at the rate of 3 kg m-3. Background fertilization with simple calcium superphosphate (19.5%) was added at 2 kg m-3 of substrate. The periodic fertilizations were done by irrigation according to the universal formula of Steiner (1984): 25% in seedling; 50% in the vegetative stage and 100% in the reproductive stage.
Three plants were randomly chosen as parents by morphotype and when flowering started the direct crosses were made according to p(p-1)/2 combinations and their reciprocals, as well as self-fertilization in the controls. Two hybridization methods were evaluated for the management of the female parent. The first consisted of the complete removal of the corolla in the emasculation, while in the second, only two petals of the flower were removed. In all hybridization cases, the emasculation was performed in flowers with immature anthers prior to anthesis, which happened when the flower buds prior to its opening exhibited in the upper part the coloring of the petals, a phenological stage in which style and stigma are at the anthers level and is an indicator of receptivity; whereas male parents were of flowers of the day that displayed anthesis with abundant pollen of creamy and hydrated color.
Pollinization was by anther detachment from the male parent and its friction in the stigma of the female parent. Hand-pollinated and self-fertilized flowers were labeled and isolated with organza bags for 72 h to avoid contamination with unwanted pollen (Sánchez et al., 2010). A total of 60 direct crosses and 60 reciprocals were made with equal numbers for each combination. The success of fertilization was considered with the development of the ovary within 10 days of pollination, although the cross success was considered with the fruit mooring and physiological maturity development.
The contrast of progenitors and hybrids was made according to 20 qualitative and quantitative morphological descriptors of the Capsicum descriptor guide of IPGRI (1995) measured from when 50% of the plants had at least one open flower. For each plant, stem shape and color, plant height and length and width of leaf were measured. For each flower, corolla color, color and position of the style, corolla diameter, number and length of petals and anther length; while for fruits, length of pedunce, fruit color, fruit length and width, pericarp thickness, number of locules and number of seeds per fruit were measured. Flower characters were measured in fully open flowers after anthesis, while fruit evaluation was at physiological maturity. The data of the evaluated variables were analyzed by descriptive and inferential statistics with the Info Stat program (Di Rienzo et al., 2011).
For the qualitative characteristics a frequency analysis was carried out according to the scales described in the guide. In order to obtain the statistical significance of the quantitative traits, a variance analysis was performed for each variable and where there were differences the Duncan mean comparison test (p≤ 0.05) was used. In order to estimate the linear association degree between evaluated variables, the Pearson correlation coefficients and their associated probability were calculated. Estimates of heterosis (H) were made based on the formula H= ((F1 - MP)/MP))*100, where: F1 was the average value of the cross and MP was the value of the best progenitor (Reyes, 1985).
Results and discussion
Hybridized flowers and self-pollinated controls with protection of organza bags showed on average 25% of fruit engraftment 10 days after pollination (Figure 1), a low percentage when compared to the stall observed in the absolute control, which was left to free and unprotected pollination which presented 90% mooring (Figure 1). Hybridization in buds at the opening stage to which only two lateral petals were detached showed more than twice as many engraftment tan those who had whole corolla removed. The latter method was the least convenient with only 12% mooring; however, for both hybridization methods, the fruit mooring was below 25%.
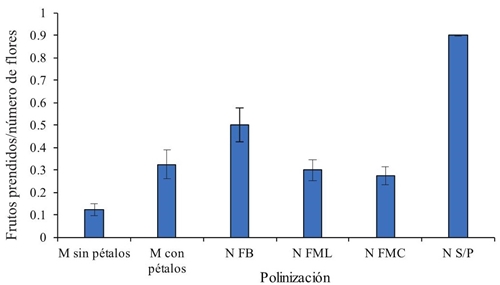
Figure 1 Proportion of engraftment fruits by number of flowers with two methods of manual hybridization and free pollination. Bars are standard error.
In the hybridization method in which the corolla was completely removed, floral structures probably collapsed as they were more exposed to adverse environmental factors. Another factor could be the mechanical damage to the pistil during manipulation for emasculation and pollination. However, when caring for self-pollinated controls with protection, they were on average affected by more than 50% of fruit engraftment compared to the total free-pollination control, suggesting that the cover used for flowers insulation is the main factor affecting the fruits engraftment. Similar results on hybridization with successful fruit engraftment until seed production of between 40 and 50% are reported in cotton by Santhy et al. (2008).
The flowers selected as female progenitors and candidates for emasculation had closed buds close to open, because in most cases in open flowers (90%), the anthers even without complete dehiscence had already released pollen, giving them a cleistogamy character. However, George (1999) points out that particularly in the Capsicum genus, although when many of its species are self-pollinate, it is difficult to consider as such due to differences observed among or even within its species, often influenced by the environment.
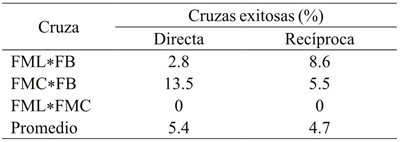
Of the hybrids with engraftment fruit at 10 days of pollination, its number for development to physiologically mature fruit decreased due to abortions in the process, with an average of 5% of fruits obtained from the total of the direct and reciprocal crosses (Table 1). There were no significant differences between direct or reciprocal crosses according to t-test (p= 0.42), suggesting no maternal effects due to the direction of the crosses. Crosses corresponding to FML*FMC in either direction did not work with ovary shedding 10 days after hybridization. In contrast, Rêgo et al. (2009), report significant differences in reciprocal effects on 56 direct and reciprocal hybrids obtained from eight progenitors of C. baccatum, suggesting maternal effects on crosses of this species; even, the same authors mention the existence of cytoplasmic effects in different species of this genus.
Table 1 Proportion of success of mature fruit obtaining in direct and reciprocal crosses between three progenitor genotypes of C. pubescens.
From six qualitative characters measured, four of them contrasting, in stem color, corolla color, filament and style color, were found in the FB parent and were recessive to the corresponding FML and FMC in the direct resulting hybrids and reciprocal FML*FB and FMC*FB (Table 3). However, the color of the fruit was contrasting for the three progenitors and hybrids showed intermediate values to those of their progenitors, suggesting codominance in the allelic relationship of this character (Table 2). The position of the style in both the parents and the hybrids were heteromorphic, with values, depending on the progenitor, from 70 to 90% for the position “at the same level” of the anthers and from 10 to 30% for the excerpt position.
Table 2 Qualitative characters in direct and reciprocal F1 progenitors and hybrids of three contrasting collections of Manzano pepper grown in southern México state.
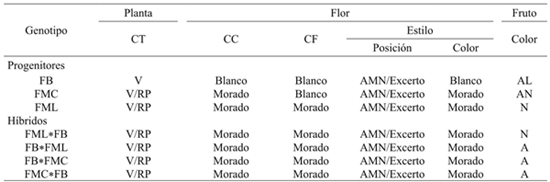
Table 3 Mean of different plant, flower and fruit characters in F1 progenitors and hybrids, direct and reciprocal from three C. pubescens collections grown in the southern México state.
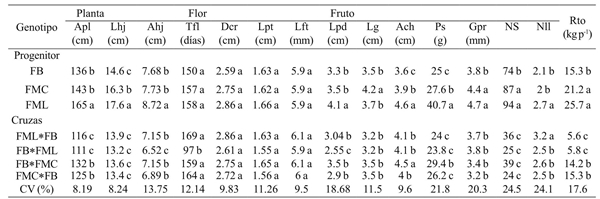
Medias con una letra común no son significativamente diferentes (Duncan, p> 0.05).
The FMC collection showed the highest values for the excerpt position, while the lowest were for FB. According to different authors (Azurdia, 1995; Berke, 2000; Barrios et al., 2007), the stigma position at the anthers level facilitates self-pollination, whereas when the stigma is excerpt the percentage of cross-pollination increases. Such is the case of creole pepper morphotypes (C. annuum) collected in Yucatan that commonly show an excerpt stigma, which has favored cross-pollination and therefore the formation of new variants or hybrids is frequent (Latournerie, 2001). In C. pubescens stigma in most cases at the same level of the anthers has possibly contributed to self-fertilization, in addition to the characteristic dehiscence of anthers before floral opening, those characteristics have favored local farmers that propagate this species by seed, without obvious variations in the offspring and therefore the propagation method has contributed to the conservation of its different genotypes. However, it should be noted that, regardless of whether C. pubescens is autogamous, the mechanisms of natural pollination by self-fertilization or crossbreeding show a continuous variation in its relative frequencies.
In quantitative plant variables, the hybrids behaved similarly to the FB and FMC progenitors, in the three measured variables of plant height, leaf length and leaf width. Even at plant height, the FML*FB hybrid and its reciprocal showed significantly lower values than FB, identified as the smallest progenitor (Table 3). The hybrids leaf length in all cases was like the FB parent type; while for the width it corresponded to the FMC parent, except for the FB*FML cross, whose width corresponded to the FB parent. The results show that the low-carcass character with short and narrow leaves were dominant over those of plants with higher height as FML, which shows significantly greater height and leaf length and width. These results coincide with that reported by Rêgo et al. (2009), who cite the presence of non-additive genes as responsible for the canopy size in the C. baccatum plant.
Although in general the plant architecture is the sum of a varied number of physiological and genetic pathways that give rise to a unique appearance for each species (Sussex and Kerk, 2001), the simpoidal branching system (non- apical dominance) of Capsicum genus and development of unique axillary flowers is regulated by the Fasciculata gene, whose recessive (fa) expresses a certain growth with multiple flowers in the axillae (Elitzur et al., 2009). In this investigation the parents and descendants also showed sympoidal branching with development of one or two axillary flowers, this last only for the FB progenitor; and regarding to the plant size, this was affected by the internodes length expressed in plant height and leaf size (Table 3).
From the quantitative traits corresponding to flowering only significant differences were found in time at the beginning of flowering (Table 3), where the FB*FML cross was one third of the time earlier than FB, a parent described as an early flowering one. This represented a difference of 50 days, time that from the commercial point of view could be significant. According to Elitzur et al. (2009), this character is pleiotropically affected also by the Fasciculata gene, which means a wide variation among the different species of Capsicum genus or even within them as it is observed in the evaluated phenotypic variants of C. pubescens (FB, FMC And FML).
Of the seven characters evaluated in fruit, five (Lpd, Lg, Ps, Gpr and Nll) corresponding to the type of the FB parent were expressed in the majority of evaluated hybrids (61%). Exceptions were the fruit width (Ach) that in three of the evaluated hybrids showed characters of the FMC progenitor and only the FB*FMC cross showed a value comparable to FML, which is the progenitor with the largest fruit width. In the variable number of seeds, there were no significant differences between the hybrids, but there existed with the three progenitors, showing on average 58% lower seed number than the parent with the lowest value (FB). Only the FB*FMC hybrids and their reciprocal showed similar yields to the lower yielding parent which is FB, since the cross FB*FML and its reciprocal showed values even significantly lower than the FB parent.
Of the four obtained hybrids, direct and reciprocal, those corresponding to FML*FB and its reciprocal showed in five measured characters significantly lower values to the progenitor with the lowest values. Estimates of heterosis generally showed a decrease in the vigor of the hybrids compared to the best progenitor, showin in most cases negative heterosis (Table 4), possibly as a result of a close relationship between the parents; since according to Geleta et al. (2004), hybrids resulting from closely related parents show low heterosis values. However, the same authors also mention similar values for hybrids of distantly related progenitors.
Table 4 Heterosis (%) with respect to the best progenitor of two direct crosses and their respective reciprocal for fourteen plant, flower, fruit and yield variables in C. pubescens.
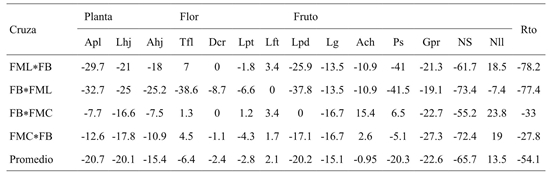
Apl= altura de la planta; Lhj= longitud de hoja; Ahj= ancho de hoja; Tfl= tiempo a floración; Dcr= diámetro de corola; Lpt= longitud de pétalo; Lft= longitud de filamento; Lpd= longitud de pedúnculo; Lg= largo de fruto; Ach= ancho de fruto; Ps= peso de fruto; Gpr= grosor del pericarpio; Ns= número de semillas por fruto; Nll= número de loculos; Rto = rendimiento.
Conversely, they mention that crosses with intermediate divergence show higher heterosis values for characters such as length, weight and fruit yield. Negative values of heterosis were observed in the plant characters fruit and yield. Although for flower characters the heterosis values were very close to zero (Table 5). One of the drawbacks in the Manzano pepper crop and basically in the most cultivated morphotype that is FML used in this research is the habit of indeterminate growth that obliges to give the plant conduction for the desired architecture, generating consumption of time and resources.
Therefore, the decrease of plant size is an advantage from the negative heterosis found for this character; however, backcross series or developing isogenic lines (Alonso-Blanco y Koornneef, 2000; Thies y Fery, 2000) will be necessary to improve fruit size and yield, which showed in the case of the latter a negative heterosis of more than 50%. According to Rêgo (2009) the canopy character of the plant in C. Baccatum is influenced by non-additive effects, which might be dominance or epistatic.
Table 5 Pearson correlation matrix for 15 characters of plant, flower and fruit of F1 progenitors and hybrids of C. pubescens.

Apl= altura de la planta; Lhj= longitud de hoja; Ahj= ancho de hoja; Tfl= tiempo a floración; Dcr= diámetro de corola; Lpt= longitud de pétalo; Lft= longitud de filamento; Lpd= longitud de pedúnculo; Lg= largo de fruto; Ach= ancho de fruto; Ps= peso de fruto; Gpr= grosor del pericarpio; Ns= número de semillas por fruto; Nll= número de lóculos; Rto= rendimiento; *= significativo (p≤ 0.05); **= altamente significativo (p≤ 0.01).
The variables Apl, Lhj and Ahj are correlated among them with highly significant positive values, demonstrating its close association and effect on plant size and vigor (Table 5). The same variables were also positively and statistically correlated with the fruit variables of Lpd, Ps, Gpr, Ns and Rto, which indicates a direct influence of plant vigor on fruit size. The number of seeds per fruit was positively and significantly correlated with the pericarp thickness, which associates this character as the one with the greatest influence on the quantity of obtained seeds.
Conclusions
In the hybridization methods, the isolation of manually pollinated flowers affected by more than 50% the fruit picking, so, in addition to the handling that must be of high skill, other materials other than those used for insulation against insects or other environmental factors that promote cross-pollination with foreign pollen or the death of the flower or fruit must be considered. The position of stigma with respect to anthers in both progenitors and hybrids was heteromorphic, with 70-90% at the anther level, suggesting a high percentage of self-pollination that allowed the conservation of morphotypes by seeds obtained from fee pollination.
The variants of four of six measured qualitative characters (stem color, corolla color, filament color and style color) were found in the FB progenitor and were recessive to the corresponding FML and FMC. While in quantitative characters the smaller variants in the plant height and leaf length and leaf width present in the FB and FMC morphotypes were dominant in the hybrids and negatively affected fruit yield and heterosis. Therefore, the decrease in plant size in the obtained hybrids was favorable for plant management aspects; however, additional genetic improvement strategies are required such as selection of segregants and backcrosses to improve fruit size.
Literatura citada
Alonso, B. C. and Koornneef, M. 2000. Naturally occurring variation in Arabidopsis: an underexploited resource for plant genetics. Trends Plant Sci. 5(1):22-29. [ Links ]
Azurdia, C. G. 1995. Capsicum spp. In: caracterización de algunos cultivos nativos de Guatemala. International Borrad for Plant Genetic Resources, Universidad de San Carlos de Guatemala, Facultad de Agronomía, Instituto de Ciencia y Tecnología Agrícola. 135-142 pp. [ Links ]
Berke, T. G. 2000. Hybrid seed production in Capsicum. Journal New Sedes. 1:49- 67. [ Links ]
Barrios, O.; Fuentes, V.; Shagarosdky, T.; Cristóbal, R.; Castiñeiras, L.; Fundora, Z. y León, N. 2007. Nuevas combinaciones híbridas de Capsicum spp. en sistemas de agricultura tradicional de occidente y oriente de Cuba. Agrotecnia de Cuba. 31(2):327-335. [ Links ]
Bosland, P. W. and Vostava, E. J. 2000. Peppers: vegetable and spice Capsicums. CABI Publishing, New York, USA. 229 p. [ Links ]
Daskalov, S. and Mihailov, L. 1988. A new method for hybrid seed production based on cytoplasmic male sterility combined with a lethal gene and a female sterile pollenizer in Capsicum annuum L. Theoretical and Applied Genetics. 76(4):530-532. [ Links ]
Di Rienzo, J. A.; Casanoves, F.; Balzarini, M. G.; González, L.; Tablada, M. y Robledo, C. W. 2011. Infostat: programa de cómputo. Versión 24-03-2011. Universidad Nacional de Córdoba. Cordoba, Argentina. 331p. [ Links ]
Elitzur, T.; Nahum, H.; Borovsky, Y.; Pekker, I.; Eshed, Y. and Paran, I. 2009. Co-ordinated regulation of flowering time, plant architecture and growth by Fasciculate: the pepper orthologue of self pruning. J. Exp. Bot. 60(3):869-880. [ Links ]
Geleta, L. F. and Labuschagne, M.T. 2004. Comparative performance and heterosis in single, three-way and double cross pepper hybrids. J. Agric. Sci. 142:659-663. [ Links ]
George, R. A. T. 1999. Vegetable seed production. CABI Publishing. 327 p. [ Links ]
Harlan, J. R. 1975. Crops and man. American Society of Agronomy. Madison Wisconsin, USA. 306 p. [ Links ]
Harrison, R. G. 1990. Hybrid zones: windows on evolutionary process. Oxford Surveys in Evolutionary Biology. 7:69-128. [ Links ]
Latournerie, L. C. S., J. L.; Pérez, M.; Castañon, G.; Rodríguez, S. and Arias, L. (2001). Valoración in situ de la diversidad morfologica de chiles (Capsicum annuum y Capsicum chinense). en Yaxcabá, Yucatán. Revista Fitotecnia Mexicana. 25(1):25-33. [ Links ]
PGRI-AVRDC-CATIE (1995). Descriptores para Capsicum (Capsicum spp.). In: Instituto Internacional de Recursos Fitogenéticos, Roma, Italia, Centro Asiático para el Desarrollo y la Investigación relativos a los Vegetales, Taipei, Taiwán y Centro Agronómico Tropical de Investigación y Enseñanza. Turrialba, Costa Rica. 50 p. [ Links ]
Pickersgill, B. 1997. Genetic resources and breeding of Capsicum spp. Euphytica. 96(1):129-133. [ Links ]
Reyes, C. P. 1985. Fitogenotecnia básica y aplicada. AGT Editor, S. A. México. 460 p. [ Links ]
Rêgo, E. R.; Rêgo, M. M.; Cruz, C. D.; Finger, F. L. and Casali, V. W. D. (2009). A diallel study of yield components and fruit quality in chilli pepper (Capsicum baccatum). Euphytica. 168(2):275-287. [ Links ]
Rieseberg, L. H. 1997. Hybrid origins of plant species. Annual Review of Ecology and Systematics. 28: 359-389. [ Links ]
Sahagun, C. J. (1999). Efectos de aptitud combinatoria en poblaciones de tomate de cáscara (Physalis exocarpa Brot.). Rev. Chapingo Ser. Hortic. 5(1):23-27. [ Links ]
Sánchez, S. H.; González, H. V.; Cruz, P. A.; Pérez, G. M.; Gutíerrez, E. A.; Gardea, B. A. y Gómez, L. M. 2010. Herencia de capsaicinoides en chile Manzano (Capsicum pubescens R. y P.). Agrociencia. 44(6):655-665. [ Links ]
Santhy, V.; Khadi, B. M.; Singh, P.; Vijaya, K. P. R.; Deshmukh, R. K. and Vishwanathan, A. 2008. Hybrid seed production in cotton. CICR Technical Bulletin Núm. 35. Central Institute for Cotton Research Nagpur, India. 25p. [ Links ]
Steiner, A. A. 1984. The universal nutrient solution. In: proceedings 6 th International Congress on Soilles Culture. Wageningen. The Netherlands. 633-650 pp. [ Links ]
Sussex, I. M. and Kerk, N. M. 2001. The evolution of plant architecture. Current Opinion in Plant Biology. 4:33- 37. [ Links ]
Thies, J. A. and Fery, R. L. 2000. Characterization of resistance conferred by the N gene to Meloidogyne arenaria races 1 and 2, M. hapla, and M. javanica in two sets of isogenic lines of Capsicum annuum L. J. Am. Soc. Hortic. Sci. 125(1):71-75. [ Links ]
Received: February 2017; Accepted: May 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
