











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El chile manzano (Capsicum pubescens R y P) con el potencial de ser una alternativa viable para el desarrollo económico de regiones como la del sur del estado de México, no ha alcanzado la importancia de cultivos como los florícolas por factores como la ausencia de un manejo tecnológico adecuado de su cultivo y frutos de calidad heterogénea; este último afectado principalmente por la variabilidad genética existente de los materiales autóctonos cultivados, los cuales contrariamente a la desventaja comercial, representan un amplio reservorio genético que puede ser aprovechado para la selección de diferentes atributos cualitativos y cuantitativos de importancia agronómica en esta especie (Pickersgill, 1997).
Del género Capsicum se reporta una vasta diversidad entre sus especies, así como variabilidad dentro de éstas para caracteres de follaje y del fruto (Bosland y Vostava, 2000), la cual no necesariamente está relacionada con los centros de origen (Harlan, 1975). Por lo tanto la observación y registro de la variabilidad crean las bases para la selección de materiales y técnicas de mejoramiento genético, como la hibridación, que permitan su aprovechamiento (Sahagun, 1999). La hibridación en sentido amplio precisa a la descendencia de la cruza de individuos de poblaciones distinguibles entre sí en uno o más caracteres heredables (Harrison, 1990) y constituye una herramienta adecuada para el análisis de la segregación de caracteres e incluye efectos como el incremento de variación genética intraespecífica, origen y transferencia de variaciones genéticas, el origen de nuevos ecotipos o especies y el reforzamiento o rompimiento de barreras reproductivas.
La amplia diversidad de micro climas en México favorece la existencia de variación dentro de especies como en la zona sur del Estado de México, donde se cultivan diferentes morfotipos de C. pubecens, los cuales representan un valioso reservorio de germoplasma que pudiera ser de utilidad en el mejoramiento de esta especie. El objetivo de la presente investigación fue desarrollar híbridos de cruzas directas y recíprocas de tres morfotipos de chile manzano (Capsicum pubescens R y P) contrastantes en diferentes caracteres y evaluarlos para diferentes atributos cualitativos y cuantitativos.
Materiales y metodos
El experimento se estableció de 2012 a 2013 en invernaderos del Centro Universitario UAEM Tenancingo localizado a 18o 58’ 2” latitud norte y 99o 36’ 44” longitud oeste y a una altitud de 2064 m, en Tenancingo, Estado de México. El material biológico fueron tres colectas de la región sur del Estado de México contrastantes a diferentes caracteres morfológicos y que se identificaron como flor blanca entrenudos cortos (FB); flor morada entrenudoslargos (FML) y flor morada entrenudos cortos (FMC).
Semillas extraídas de frutos en madurez fisiológica de cada morfotipo se sembraron en sustrato cernido a base de hojas de encino en germinadores de poliestireno de 200 cavidades. El sustrato se ajustó a pH de 5.5 con cal agrícola a razón de 600 g m-3. Como fertilización de fondo se utilizó superfosfato de calcio simple (19.5%) a razón de 1 kg m-3. Las charolas se colocaron en invernadero y bajo malla sombra de 50% con humedad relativa entre el 60% y 90%. El trasplante inicial a vasos de poliestireno de 250 ml se realizó cuando las plantas mostraron10 hojas verdaderas (40 días después de siembra).
El trasplante definitivo se hizo en bolsas de plástico calibre 600 de 60 L de capacidad cuando las plantas presentaron la primera bifurcación (80 días después de siembra). La densidad de plantación fue de 1.5 plantas m-2. En ambos trasplantes el sustrato fue una mezcla de tierra arenosa, tepojal y materia orgánica, en una relación de 5:4:1 respectivamente. El pH se ajustó a 5.5 con cal agrícola a razón de 3 kg m-3. Se dio una fertilización de fondo con superfosfato de calcio simple (19.5%) a razón de 2 kg m-3 de sustrato. Las fertilizaciones periódicas se hicieron por el riego de acuerdo a la formula universal de Steiner (1984): 25% en plántula; 50% en etapa vegetativa y 100% en etapa reproductiva.
Se eligieron como progenitores tres plantas al azar por morfotipo y cuando la floración inició se hicieron las cruzas directas resultantes de acuerdo a p(p-1)/2 combinaciones y sus recíprocos, así como también autofecundaciones en los testigos. Se evaluaron dos métodos de hibridación por el manejo del progenitor femenino. El primero consistió en la remoción completa de la corola en la emasculación, mientras que en el segundo, solo se removieron dos pétalos de la flor. En todos los casos de hibridación, la emasculación se hizo en flores con anteras inmaduras previo a la antesis, lo cual ocurrió cuando los botones florales previos a su apertura exhibieron en su parte superior la coloración de los pétalos, etapa fenológica en la que el estilo y estigma están a la altura de las anteras y es indicador de receptibilidad; mientras que los progenitores masculinos fueron de flores del día que presentaron antesis con abundante polen de color cremoso e hidratado.
La polinización fue mediante desprendimiento de antera del progenitor masculino y fricción de ésta en el estigma del progenitor femenino. Las flores polinizadas manualmente y las autofecundadas fueron etiquetadas y aisladas con bolsas de organza por 72 h para evitar contaminación con polen no deseado (Sánchez et al., 2010). Se hicieron en total 60 cruzas directas y 60 reciprocas con igual número para cada combinación. El éxito de la fecundación se consideró con el desarrollo del ovario a los 10 días de la polinización, aunque el éxito de la cruza se consideró con el amarre del fruto y desarrollo a madurez fisiológica.
El contraste de progenitores e híbridos se hizo de acuerdo a 20 descriptores morfológicos cualitativos y cuantitativos de la guía de descriptores para Capsicum del IPGRI (1995) medidos a partir de cuando 50% de las plantas presentaron por lo menos una flor abierta. Para planta se midieron forma y color del tallo, altura de la planta y largo y ancho de la hoja, para flor se midieron color de la corola, color y posición del estilo, diámetro de la corola, número y longitud de pétalos y longitud de la antera; mientras que para fruto se midieron longitud de pedúnculo, color del fruto, largo y ancho de fruto, grosor del pericarpio, número de lóculos y número de semillas por fruto. Los caracteres de flor se midieron en flores completamente abiertas después de la antesis, mientras que la evaluación de fruto fue en madurez fisiológica. Los datos de las variables evaluadas se analizaron mediante estadística descriptiva e inferencial con el programa Info Stat (Di Rienzo et al., 2011).
En los caracteres cualitativos se efectuó un análisis de frecuencias de acuerdo a las escalas descritas en la guía. Para obtener las significancias estadísticas de los caracteres cuantitativos se realizó un análisis de varianza para cada variable y donde hubo diferencias se utilizó la prueba de comparación de medias Duncan (p≤ 0.05). Para estimar el grado de asociación lineal entre las variables evaluadas se calcularon los coeficientes de correlación de Pearson con su probabilidad asociada. Las estimaciones de heterosis (H) se realizaron con base en la fórmula H= ((F1 - MP)/MP))*100, donde: F1 fue el valor promedio de la cruza y MP fue el valor del mejor progenitor (Reyes, 1985).
Resultados y discusión
Las flores hibridadas y las testigo autopolinizadas con protección de bolsas de organza presentaron en promedio 25% de prendimiento de frutos a los 10 días después de la polinización (Figura 1), porcentaje bajo si se compara con el prendimiento observado en el testigo absoluto, que se dejó a polinización libre y sin protección que presentó 90% de amarre (Figura 1). La hibridación en flores en botón a punto de apertura a las que se les desprendieron solo dos pétalos laterales presentó más del doble de prendimiento que aquellas a las que les fue removida la corola completa. Este último método fue el menos conveniente con solo 12% de amarre; sin embargo, para ambos métodos de hibridación, el amarre de fruto fue por debajo de 25%.
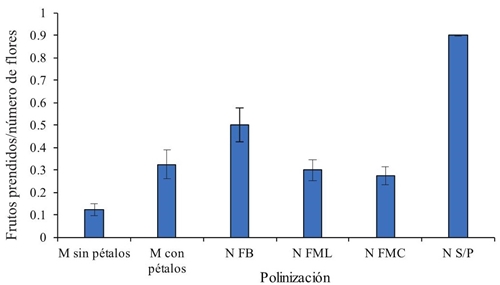
Figura 1 Proporción de frutos prendidos por número de flores con dos métodos de hibridación manual y polinización libre. Las barras son el error estándar.
En el método de hibridación en el que se retiró completamente la corola, probablemente las estructuras florales colapsaron al quedar más expuestas a factores ambientes adversos. Otro factor pudo ser el daño mecánico al pistilo durante la manipulación por la emasculación y la polinización. Sin embargo, al atender a los testigos autopolinizados con protección, éstos en promedio se vieron afectados a prendimiento de fruto en más de 50%, con respecto al testigo absoluto de polinización libre y sin protección, lo que sugiere que la cubierta utilizada para aislamiento de flores es el principal factor que afecta el prendimiento de los frutos. Resultados similares sobre hibridación con prendimiento de fruto con éxito hasta producción de semilla de entre 40 y 50% son los reportados en algodón por Santhy et al. (2008).
Las flores seleccionadas como progenitores femeninos y candidatas a emasculación fueron con botón cerrado próximo a abrir, debido a que en la mayoría de los casos en flor abierta (90%), las anteras aun sin dehiscencia completa ya habían liberado polen, dándoles un carácter de cleistogamia. Sin embargo, George (1999) señala que particularmente el género Capsicum aun cuando muchas de sus especies se autopolinizan, es difícil considerarlo como tal debido a diferencias observadas entre sus especies o incluso dentro de ellas, muchas veces influidas por el ambiente.
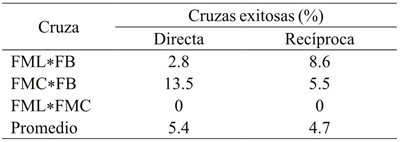
De los híbridos con fruto prendido a 10 días de la polinización, su número para desarrollo a fruto fisiológicamente maduro disminuyó por abortos en el proceso, con un promedio de 5% de frutos obtenidos del total de las cuzas directas y recíprocas hechas (Cuadro 1). No hubo diferencias significativas entre cruzas directas ni recíprocas de acuerdo a prueba de t (p= 0.42), lo que sugiere ausencia de efectos maternos por la dirección de las cruzas. Las cruzas correspondientes a FML*FMC en cualquier dirección no funcionaron con desprendimiento de ovario a 10 días después de la hibridación. Contrariamente, Rêgo et al. (2009), reportan diferencias significativas en los efectos recíprocos en 56 híbridos directos y recíprocos obtenidos con ocho progenitores de C. baccatum, lo que sugiere efectos maternos en cruzas de esta especie; incluso, los mismos autores mencionan la existencia de efectos citoplásmicos en diferentes especies de este género.
Cuadro 1 Proporción de éxitos a obtención de fruto maduro en cruzas directas y recíprocas entre tres genotipos progenitores de C. pubescens.
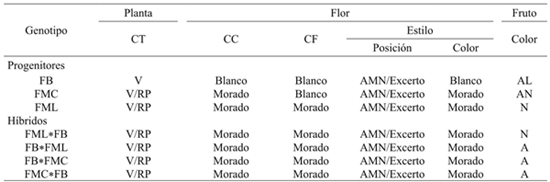
De seis caracteres cualitativos, cuatro de ellos fueron contrastantes, en color del tallo, corola, filamento y estilo, se encontraron en el progenitor FB y fueron recesivos a los de FML y FMC en los híbridos resultantes directos y recíprocos de FML*FB y FMC*FB (Cuadro 3). Sin embargo, el color del fruto fue contrastante para los tres progenitores y los híbridos mostraron valores intermedios a sus progenitores, que sugiere codominancia en la relación alélica de este carácter (Cuadro 2). La posición del estilo, tanto en los progenitores como en los híbridos resultaron heteromórficos, con valores, dependiendo del progenitor, de 70 a 90% para la posición “al mismo nivel” de las anteras y de 10 a 30% para la posición excerta.
Cuadro 2 Caracteres cualitativos en progenitores e híbridos F1 directos y recíprocos de tres colectas contrastantes de chile manzano cultivadas en el sur del estado de México.
Cuadro 3 Medias de diferentes caracteres de planta, flor y fruto en progenitores e híbridos F1 directos y recíprocos de tres colectas de C. pubescens cultivadas en el sur del estado de México.
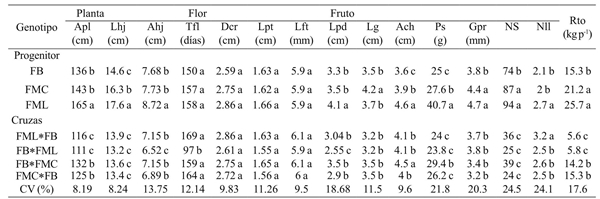
Medias con una letra común no son significativamente diferentes (Duncan, p> 0.05).
La colecta FMC presentó los mayores valores para la posición excerta, mientras que los menores fueron para FB. De acuerdo a Azurdia (1995); Berke (2000); Barrios et al. (2007) la posición del estigma al nivel de las anteras facilita la autopolinización, mientras que cuando el estigma es excerto aumenta el porcentaje de polinización cruzada. Tal es el caso de morfotipos de chiles criollos (C. annuum) colectados en Yucatán que presentan comúnmente el estigma excerto, lo cual ha favorecido la polinización cruzada y resulta frecuente la formación de nuevas variantes o híbridos (Latournerie, 2001). En C. pubescens el estigma en la mayoría de los casos al mismo nivel de las anteras posiblemente ha contribuido a la autofecundación, además de la característica de inicio de dehiscencia de las anteras antes de la apertura floral, características que han favorecido a los agricultores locales que propagan a esta especie por semilla, sin variaciones evidentes en los descendientes y por consiguiente el método de propagación ha contribuido a la conservación de sus diferentes genotipos. Sin embargo, debe tenerse en cuenta que C. pubescens sea autógama, los mecanismos de polinización natural por autofecundación o cruzada presentan una continua variación en sus frecuencias relativas.
En variables cuantitativas de la planta, los híbridos se comportaron de manera similar a los progenitores FB y FMC, en las tres variables medidas de altura de la planta, largo y ancho de hoja. Incluso en altura de la planta, el híbrido FML*FB y su recíproco mostraron valores significativamente menores a FB, identificado como el progenitor de menor porte (Cuadro 3). El largo de hoja de los híbridos en todos los casos fue del tipo del progenitor FB; mientras que para el ancho correspondieron al progenitor FMC, excepto para la cruza FB*FML, cuyo ancho correspondió al progenitor FB. Los resultados muestran que el carácter porte bajo, con hojas cortas y estrechas fueron dominantes en plantas con mayor altura como FML, la cual presenta mayor altura, largo y ancho de hoja. Estos resultados coinciden con lo reportado por Rêgo et al. (2009), quienes citan la presencia de genes no aditivos como responsables del tamaño del dosel de la planta en C. baccatum.
Aunque en general la arquitectura de la planta es la suma de un número variado de rutas fisiológicas y genéticas que dan origen a una apariencia única para cada especie (Sussex and Kerk, 2001), el sistema de ramificación simpoidal (no dominancia apical) del genero Capsicum y desarrollo de flores axilares únicas está regulado por el gene Fasciculata, cuyo recesivo (fa) expresa un crecimiento determinado con flores múltiples en las axilas (Elitzur et al., 2009). En la presente investigación los progenitores y descendientes también mostraron ramificación simpoidal con desarrollo de una o dos flores axilares, este último solo para el progenitor FB; y en referencia al porte de la planta este fue afectado por la longitud de entrenudos expresado en la altura de la planta y tamaño de la hoja (Cuadro 3).
De los caracteres cuantitativos correspondientes a floración solo se encontraron diferencias significativas en tiempo a inicio de floración (Cuadro 3), en donde la cruza FB*FML, fue un tercio de tiempo más precoz que FB, progenitor que se tiene descrito como de floración temprana. Esto representó una diferencia de 50 días, tiempo que desde el punto de vista comercial podría ser significativo. De acuerdo con Elitzur et al. (2009), este carácter es afectado pleiotropicamente también por el gene Fasciculata, lo que representa que haya amplia variación en las diferentes especies del genero Capsicum o incluso dentro de ellas como se observa en los variantes fenotípicos evaluados de C. pubescens (FB, FMC y FML).
De los siete caracteres evaluados en fruto, cinco (Lpd, Lg, Ps, Gpr y Nll) correspondientes al tipo del progenitor FB se expresaron en la mayoría de los híbridos evaluados (61%). Fueron excepción el caracter ancho de fruto (Ach) que en tres de los híbridos evaluados presentaron caracteres del progenitor FMC y solo la cruza FB*FMC presentó un valor equiparable a FML, progenitor con el mayor ancho fruto. En la variable número de semillas, no hubo diferencias significativas entre los híbridos, pero si con los tres progenitores, al presentar en promedio 58% menor número de semilla que el progenitor con el menor valor (FB). Solo los híbridos FB*FMC y su recíproco, mostraron rendimientos similares al progenitor de menor rendimiento que es FB, ya que la cruza FB*FML y su recíproco mostraron valores aun significativamente menores que el mismo progenitor FB.
De los cuatro híbridos obtenidos, directos y recíprocos, los correspondientes a FML*FB y su recíproco mostraron en cinco caracteres medidos valores significativamente menores al progenitor con los valores más bajos. Los cálculos de heterosis, mostraron de manera general una disminución del vigor de los híbridos con respecto al mejor progenitor, al observarse en la mayoría de los casos heterosis negativa (Cuadro 4), posiblemente como resultado de una estrecha relación entre los progenitores; ya que de acuerdo con Geleta et al. (2004), híbridos resultantes de progenitores estrechamente relacionados muestran bajos valores de heterosis. Sin embargo, los mismos autores también mencionan valores similares para híbridos de progenitores distantemente relacionados.
Cuadro 4 Heterosis (%) con respecto al mejor progenitor de dos cruzas directas y su respectivo reciproco para catorce variables de planta, flor, fruto y rendimiento en C. pubescens.
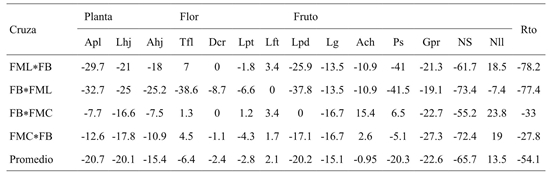
Apl= altura de la planta; Lhj= longitud de hoja; Ahj= ancho de hoja; Tfl= tiempo a floración; Dcr= diámetro de corola; Lpt= longitud de pétalo; Lft= longitud de filamento; Lpd= longitud de pedúnculo; Lg= largo de fruto; Ach= ancho de fruto; Ps= peso de fruto; Gpr= grosor del pericarpio; Ns= número de semillas por fruto; Nll= número de loculos; Rto = rendimiento.
Contrariamente, citan que cruzas con divergencia intermedia muestran mayores valores de heterosis para caracteres como, longitud, peso y rendimiento de fruto. Se observaron valores negativos de heterosis en caracteres de la planta, fruto y rendimiento. Aunque para caracteres de la flor los valores de heterosis fueron muy próximos a cero (Cuadro 5). Uno de los inconvenientes que se tiene en el cultivo de chile manzano y básicamente el morfotipo más cultivado FML utilizado en esta investigación, es el hábito de crecimiento indeterminado que da a la planta conducción para la arquitectura deseada, generando consumo de tiempo y recursos.
Cuadro 5 Matriz de correlación Pearson para 15 caracteres de planta, flor y fruto de progenitores e híbridos F1 de C. pubescens.

Apl= altura de la planta; Lhj= longitud de hoja; Ahj= ancho de hoja; Tfl= tiempo a floración; Dcr= diámetro de corola; Lpt= longitud de pétalo; Lft= longitud de filamento; Lpd= longitud de pedúnculo; Lg= largo de fruto; Ach= ancho de fruto; Ps= peso de fruto; Gpr= grosor del pericarpio; Ns= número de semillas por fruto; Nll= número de lóculos; Rto= rendimiento; *= significativo (p≤ 0.05); **= altamente significativo (p≤ 0.01).
Por lo tanto, la disminución de porte de la planta es una ventaja a partir de la heterosis negativa encontrada para este carácter; sin embargo, series de retrocruzas o desarrollo de líneas isogénicas (Alonso-Blanco y Koornneef, 2000; Thies y Fery, 2000) son necesarias para el tamaño y rendimiento de fruto, y este último tuvo heterosis negativa más de 50%. Segun Rêgo (2009) el carácter dosel de la planta en C. Baccatum es influido por efectos no aditivos, que podrían ser de dominancia o epistáticos.
Las variables Apl, Lhj y Ahj se correlacionaron entre ellas con valores positivos altamente significativos, lo que demuestra su estrecha asociación y efecto en el porte y vigor de la planta (Cuadro 5). Las mismas variables también se correlacionaron positivamente y con valor estadístico con las variables de fruto de Lpd, Ps, Gpr, Ns y Rto, lo que denota una influencia directa del vigor de la planta en el tamaño del fruto. El número de semillas por fruto se correlacionó positiva y significativamente con el grosor del pericarpio, lo que asocia a este último carácter como el de mayor influencia en la cantidad de semillas obtenidas.
Conclusiones
En los métodos de hibridación, el aislamiento de flores polinizadas manualmente afectó en más de 50% el prendimiento de fruto, por lo cual, además del manejo que debe ser de alta destreza, también se deben considerar otros materiales diferentes al empleado para el aislamiento contra insectos u otros factores ambientales que promuevan polinización cruzada con polen extraño o muerte de la flor o fruto. La posición del estigma con respecto a las anteras tanto en progenitores como en los híbridos fue heteromórfica, con 70-90% al nivel de las anteras, lo que sugiere un alto porcentaje de autopolinización que ha permitido la conservación de los morfotipos por semilla obtenida de polinización libre.
Los variantes de cuatro de seis caracteres cualitativos medidos (color del tallo, color de la corola, color del filamento y color del estilo) se encontraron en el progenitor FB y fueron recesivos a los correspondientes de FML y FMC. Mientras que en caracteres cuantitativos los variantes de menor tamaño en las variables de altura de la planta y largo y ancho de hoja presentes en los morfotipos FB y FMC fueron dominantes en los híbridos y afectaron negativamente en el rendimiento de frutos y heterosis. Por lo tanto, la disminución en el porte de la planta en los híbridos obtenidos resultó favorable para aspectos de manejo de la planta; sin embargo, se requiere de estrategias de mejoramiento genético adicionales como selección de segregantes y retrocruzas para mejorar el tamaño del fruto.

