











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En México, el maíz (Zea mays L.) es el cultivo más importante debido a que anualmente se siembran más de 7.4 millones de hectáreas, con una producción global de más de 23.2 millones de toneladas. El promedio del rendimiento de grano unitario (3.3 t ha-1) en 2014 (SIAP, 2016) indica rendimientos bajos en virtud, principalmente, a que el maíz se cultiva en alrededor de 85% de la superficie en condiciones de temporal o secano, que depende del régimen de lluvias, frecuentemente errático e insuficiente para cubrir la demanda hídrica del cultivo. Para estas condiciones variables, los programas de mejoramiento genético de maíz en México han generado una amplia diversidad de genotipos con base en la aplicación de diferentes estrategias de selección y evaluación (Ángeles-Arrieta, 2000). Específicamente, en el norte del estado de Tamaulipas la superficie cultivada con maíz varía cada año entre 30 mil y 100 mil hectáreas. Por ejemplo en 2014 se cultivaron más de 67 000 ha con rendimientos globales de más de 360 000 t (5.4 t ha-1). Las variaciones en superficie dependen fundamentalmente de la disponibilidad de agua para riego o bien, la incidencia de plagas y enfermedades. Por tal razón, alternativas culturales, genéticas, biológicas y químicas de manejo del cultivo se evalúan constantemente para maximizar rendimiento y calidad de grano (Reyes-Méndez y Cantú-Almaguer, 2006).
Los progresos sustantivos en el incremento del rendimiento de grano se fundamentan en lograr la óptima adaptación del genotipo a cada ambiente de producción, así la conjunción de caracteres deseables tales como la calidad del grano y la resistencia al estrés abiótico y biótico. Otro factor que apoya al fitomejorador es la evaluación de la magnitud y la dirección de la asociación entre el rendimiento de grano y sus componentes así como la importancia relativa de cada factor involucrado en la contribución al rendimiento de grano (Agrama, 1996). No obstante lo anterior, es frecuente observar germoplasma sin buena adaptación a ambientes variables o bien, germoplasma con adaptación ambiental específica o local (González-Martínez et al., 2014; Lozano-Ramírez et al., 2015). En maíz, se ha determinado que el número de mazorcas por planta tienen el mayor efecto directo en rendimiento de grano, aunque también hay efectos indirectos vía el número de granos por mazorca, la prolificidad o el tamaño de grano, características que sirven en el mejoramiento del rendimiento del maíz (Agrama, 1996).
Márquez-Sánchez (1991) definió como una variedad estable aquella con capacidad de amortiguar o de ajustarse a las condiciones ambientales prevalentes. En mejoramiento genético la estabilidad espacial y temporal debe evaluarse para incrementar la eficiencia del proceso de selección, debido a que las características económicamente importantes son medibles, muestran distribución continua y son poligénicas, así como afectadas por el ambiente (Lozano-Ramírez et al., 2015).
En maíz se han desarrollado variedades e híbridos con comportamiento estable mediante la aplicación de metodologías que consideran parámetros de estabilidad, particularmente del rendimiento de grano (Fan et al., 2007; Cargnelutti et al., 2009; García et al., 2009; Kandus et al., 2010; Lozano-Ramírez et al., 2015). Igualmente, diversos estudios han enfatizado que, a través del análisis de sendero secuencial, la selección de germoplasma superior en rendimiento se basa en caracteres como el número de granos por mazorca, tamaño del grano o número de mazorcas por planta, estrecha y positivamente asociados con el rendimiento del maíz (Agrama, 1996; Mohammadi et al., 2003; Nemati et al., 2009).
Entre las alternativas de análisis de la IGA están el modelo AMMI (Análisis de la interacción de efectos principales aditivos y multiplicativos) basado en un modelo lineal-bilineal y que estima estabilidad, evalúa ambientes y los clasifica (Crossa et al., 1990); así como el modelo de análisis de regresión en los sitios (SREG), también conocida como gráfico GGE biplot (Yan et al., 2000). Por otro lado, la evaluación del comportamiento de genotipos y de los efectos de los componentes en la expresión del rendimiento de grano se puede determinar con base en el análisis de sendero secuencial (ASS), donde se estiman las correlaciones entre caracteres con efectos directos e indirectos en una variable respuesta (Whittaker et al., 2009). Para seleccionar genotipos, se descomponen las correlaciones de variables en efectos causales, identificando cadenas de causalidad entre ellas. Así, el ASS discierne entre los efectos reales de las variables predictoras y el ajuste de varios grupos de datos, ofreciendo ventajas sobre el análisis de sendero convencional (Kozak y Antunes-Azevedo, 2014).
El objetivo de este trabajo consistió en evaluar el comportamiento productivo y la adaptación de híbridos de maíz a diferentes fechas de siembra y años de prueba (ambientes) en el norte de Tamaulipas, para ello se aplicó el AMMI, GGE biplot y el análisis de sendero secuencial (ASS).
Materiales y métodos
El estudio se condujo durante tres ciclos de cultivo (2006, 2007 y 2008) en el Campo Experimental Río Bravo del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) en Río Bravo, Tamaulipas, México (25º 57’ latitud norte, 98º 01’ longitud oeste; 30 msnm). Los suelos del sitio experimental son de textura arcillosa, pH= 7.5 y bajo contenido de materia orgánica (1.1%). Cada experimento incluyó cinco híbridos de maíz con grano blanco (H-437, H-439, H-440, P-3025, Tigre) y tres híbridos con grano amarillo (G-8222, P-31G98, DK- 697) (Cuadro 1) mismos que se sembraron en cuatro fechas de siembra cada siete días durante enero y febrero de cada año, todas dentro del rango óptimo para maíz en el sitio de prueba (Cuadro 2) (Reyes-Méndez y Cantú-Almaguer, 2006).
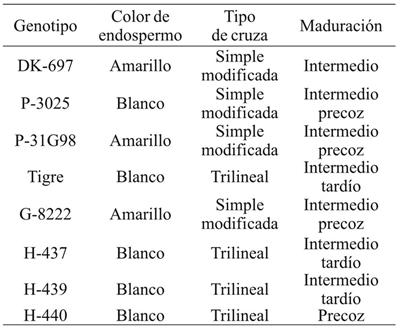
Cuadro 2 Condiciones ambientales en los experimentos con híbridos de maíz sembrados en diferentes años y fechas de siembra. Río Bravo, Tamaulipas.
AMB= ambiente; TMax= temperatura media máxima; TMin= temperatura media mínima; TMed= temperatura media; PP= precipitación pluvial total.
Cada experimento se estableció en un diseño experimental de bloques completos al azar con un arreglo en parcelas divididas, donde la parcela grande correspondió a las fechas de siembra y la parcela chica a los híbridos de maíz. Cada tratamiento tuvo cuatro repeticiones; la parcela experimental constó de cuatro surcos de 5 m de largo espaciados a 0.8 m entre sí. Los terrenos y experimentos se prepararon y condujeron de acuerdo con el paquete tecnológico para maíz generado por INIFAP en la región (Reyes-Méndez y Cantú-Almaguer, 2006).
En cada parcela experimental se estimó el peso de mazorcas ‘en campo’ (PT, que incluyó grano y olote sin ajuste de humedad), peso de mazorcas (PM, con grano y olote ajustado a 14% de humedad), rendimiento de grano (RG, ajustado al 14% de humedad), días a floración masculina (DFM) y femenina (DFF), número de mazorcas sanas (NMS) y enfermas (NME), altura de planta (AP) y de mazorca (AM), relación altura de planta mazorca (RAPM).
El análisis de la interacción genotipo-ambiente se llevó a cabo con el modelo AMMI en el programa SAS versión 9.0 (Crossa et al., 1990), además del análisis GGE Biplot (Vargas y Crossa, 2000; Burgueño et al., 2002). Para ello se generaron 12 ‘ambientes’ producto de la combinación de cuatro fechas de siembra por tres años de prueba. Posteriormente, se realizó el análisis de sendero secuencial (ASS) con base en la estimación de los valores de inflación de la varianza (VIFs) y los de tolerancia. En este caso, las variables peso de mazorca (PM), altura de planta (AP), relación altura de planta mazorca (RAPM) y número de mazorcas sanas (NMS) se consideraron variables de primer orden. Para identificar las variables de primer orden, el procedimiento se repitió considerando las variables antes mencionadas como variables dependientes de segundo orden. Los coeficientes de determinación parcial (análogo al R2 del análisis de regresión lineal) se calcularon del coeficiente de sendero para todas las variables (Williams et al., 1990).
Resultados
El análisis de varianza del modelo AMMI para rendimiento de grano (RG) mostró diferencias significativas (p≤ 0.05) entre ambientes, indicando que las diferencias entre los promedios de dicha fuente de variación contribuyeron a las diferencias en el rendimiento de grano entre genotipos de maíz. La contribución del efecto de la IGA en la suma de cuadrados fue mayor que la de genotipos, demostrando diferencias genotípicas en respuesta a los ambientes de prueba (Cuadro 3). El análisis de componentes principales indicó que los dos primeros componentes principales fueron significativos (p≤ 0.01) y, en conjunto, explicaron 76.6% (CP1 56.2% y CP2 20.4%) de la variación total debida a la interacción (datos no incluidos). El híbrido más estable y con menor IGA mostró valores entre -0.3 a 0.3 en el primer componente principal (CP1) del AMMI y fue DK-697 (Figura 1).
Cuadro 3 Cuadrados medios de análisis de varianza del rendimiento de grano de híbridos de maíz sembrados en años y fechas de siembra en Río Bravo, Tamaulipas.
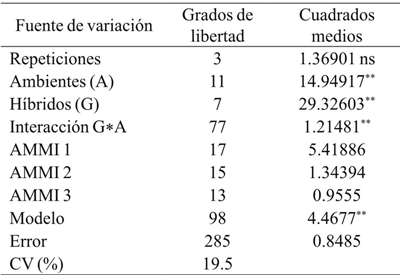
CV= coeficiente de variación; ns= no significativo; *= significativo (p≤ 0.05); **= significativo (p≤ 0.01).
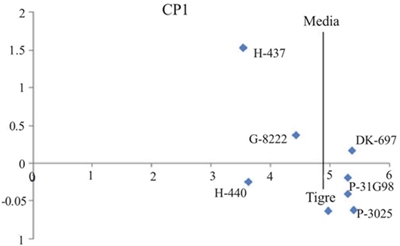
Figura 1 Dispersión de híbridos de maíz cultivados en el norte de Tamaulipas, México (2006-2008), con base en el rendimiento de grano (t ha-1) en el eje de las abscisas y el primer componente CP1 del AMMI.
Los ambientes (año*fecha de siembra) más alejados fueron 2006A, 2006B, 2006C y 2007A; mientras que los vectores más cercanos al origen correspondieron a 2008B y 2008C (Cuadro 2, Figura 2). Los híbridos más alejados del centro del biplot (H-437 y G-8222) con rendimiento de grano (RG) más alto y la mayor variabilidad a través de experimentos. Por el contrario, DK-697, H-440 y Tigre tuvieron RG estable al ubicarse más cerca del origen del biplot (Figura 2). El ASS indicó que peso de mazorca (PM), altura de planta (AP), relación altura de planta mazorca (RAPM) y número de mazorcas sanas (NMS) son variables de primer orden para RG, debido que explicaron 86% de la varianza total del RG de los híbridos evaluados; con presencia de colinealidad moderada para todas las variables medidas, con efectos fuertes por ejemplo del número de mazorcas sanas (NMS) (VIF= 1.41) y peso de mazorca (PM) (VIF= 1.19) (Cuadro 4).
Cuadro 4 Efectos directos de variables predictoras del rendimiento de grano (RG) y medidas de colinealidad en ocho híbridos de maíz cultivados en el norte de Tamaulipas, México.
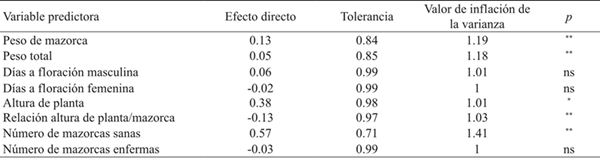
ns= no significativo; *significativo (p≤ 0.05); **= significativo (p≤ 0.01).
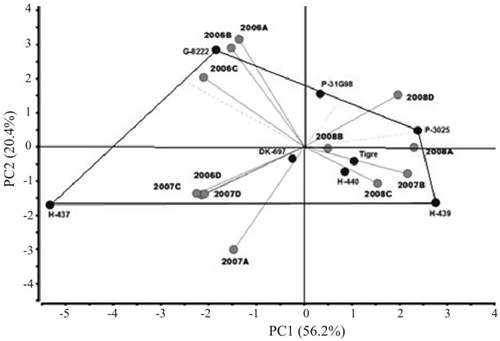
Figura 2 Gráfico biplot de híbridos de maíz cultivados en el norte de Tamaulipas, México durante 2006-2008 con base en rendimientos de grano (RG).
Las variables de primer y segundo orden demostraron que 32% de la variación total en peso de mazorca (PM) se explica con las variables días a floración masculina (DFM), días a floración femenina (DFF) y peso total (PT); mientras que 64% de la variación en el número de mazorcas sanas (NMS) se explica con el número de mazorcas enfermas (NME) y el peso de mazorca (PM) (Cuadro 5; Figura 3).
Cuadro 5 Efecto directo de variables predictoras de primer y segundo orden en el rendimiento de grano y colinealidad en ocho híbridos de maíz cultivados en el norte de Tamaulipas, México.
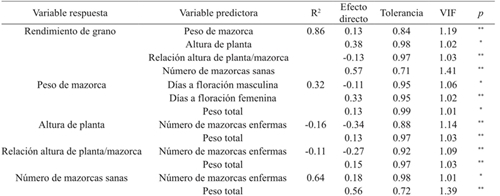
ns= no significativo; *= significativo (p≤ 0.05); **= significativo (p≤ 0.01); VIF= valor de inflación de la varianza.
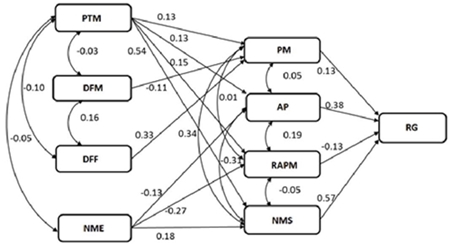
Figura 3 Análisis de sendero secuencial indica las interrelaciones de las características que contribuyen al rendimiento de grano de ocho híbridos de maíz cultivados en el norte de Tamaulipas, México. PM= peso de mazorca; PTM = peso total de mazorca; DFM= días a floración masculina; DFF= días a floración femenina; AP= altura de planta, RAPM= relación altura de planta/mazorca; NMS= número de mazorcas sanas; NME= número de mazorcas enfermas; RG= rendimiento de grano.
Discusión
En este trabajo se observaron diferencias en el comportamiento agronómico entre los ocho híbridos de maíz evaluados a través de los ambientes de prueba, producto de la combinación de tres años de evaluación y cuatro fechas de siembra, lo que a su vez refleja la influencia de las condiciones ambientales variables ocurridas durante el desarrollo de los experimentos. Efectos significativos entre genotipos de maíz y ambientes de prueba se han consignado previamente (Fan et al., 2007; Cargnelutti et al., 2009; García et al., 2009; Kandus et al., 2010; Nemati et al., 2010; Selvaraj y Ngarajan, 2011; Lozano-Ramírez et al., 2015).
El híbrido más estable y con menor IGA fue DK-697, de acuerdo con los resultados de las dos estrategias de análisis, el AMMI y el GGE Biplot. Además, ésta última estrategia indicó que tres ambientes de 2006 y uno de 2007 contribuyeron en menor proporción a la interacción genotipo x ambiente y por ende exhibieron mejor capacidad para discriminar genotipos. Dichos ambientes ofrecieron mejores condiciones ambientales en virtud de que consignan los mayores promedios de lluvias. Por el contrario, dos ambientes de 2008 tuvieron la menor interacción ambiental dado que muestran mayores temperaturas que afectaron por igual al germoplasma evaluado
Aunque Balestre et al. (2009) y Luenzzo-De Oliveira et al. (2010) coinciden en señalar que el modelo AMMI es en general más preciso para evaluar la adaptabilidad y estabilidad de germoplasma de maíz con base en rendimiento de grano, en este trabajo se observó que ambas estrategias coincidieron en identificar al híbrido más estable, DK-697, tal como también lo indican Mitrovic et al. (2012). Por el contrario, Yan et al. (2007) mencionan que, no obstante ambos métodos combinan los efectos del genotipo y de la interacción genotipo*ambiente en el análisis de mega-ambientes, el GGE biplot es superior al AMMI pues explica mejor los efectos genotípicos y de la interacción y tiene la propiedad del ‘producto interno’ del biplot, así como su poder de discriminación vs la visión de representatividad. Por ello, consideramos que la combinación de ambas estrategias mejorarían la eficiencia en el proceso de evaluación de la estabilidad del rendimiento en germoplasma de maíz, con la ventaja de que el GGE biplot además aportaría información para caracterizar y estratificar ambientes de prueba y reducir el número de experimentos o sitios de prueba hasta en 70% (Mitrovic et al., 2012) con el consiguiente ahorro de tiempo, dinero y trabajo sin pérdida de eficiencia y precisión de la evaluación.
Las variables peso de mazorca, altura de planta, relación altura de planta/mazorca y número de mazorcas sanas se identificaron como variables directamente asociadas con el rendimiento de grano del maíz (variables de primer orden), además de colinealidad entre todas las variables medidas, con efectos fuertes en número de mazorcas sanas y peso de mazorca. Estudios previos indican que la selección de germoplasma de maíz superior en rendimiento de grano se basa en la consideración de caracteres como el número de granos por mazorca, tamaño del grano o número de mazorcas por planta, estrecha y positivamente asociados con el rendimiento del maíz (Agrama, 1996; Mohammadi et al., 2003; Nemati et al., 2009; Jamshidian et al., 2013). Jamshidian et al. (2013); Rashidi et al. (2013) incluso observaron asociaciones directas positivas de los contenidos de proteína con el peso del grano y de la mazorca.
El uso conjunto de las estrategias AMMI, GGE biplot y análisis de sendero secuencial se han aplicado para evaluar la significancia y magnitud del efecto de la interacción genotipo x ambiente; así como para identificar las relaciones entre el rendimiento de grano y los componentes del rendimiento en cultivos como la soya [Glycine max (L.) Merr.] (Ascencio-Luciano et al., 2013). También, se han aplicado en la caracterización de la respuesta a la sequía en trigo (Triticum aestivum L.) (Farshadfar et al., 2012). Dichos modelos son buenas herramientas estadísticas para la evaluación, caracterización y selección de germoplasma con base en su comportamiento en ambientes variables (Tai y Tarn, 2003), de modo que el mejoramiento genético del maíz por rendimiento de grano será eficiente al considerar las variables altamente asociadas con el rendimiento de grano para el correcto diseño y uso de índices de selección en mega-ambientes.
Conclusiones
En este trabajo, la combinación del modelo AMMI, el GGE biplot y análisis de sendero secuencial (ASS) permitió la identificación de germoplasma de maíz con estabilidad del rendimiento de grano y rendimiento de grano promedio alto (DK-697), así como la identificación de características estrechamente asociadas con el rendimiento de grano (peso de mazorca, altura de planta, relación altura de planta/mazorca y número de mazorcas sanas) de híbridos de maíz cultivados en diferentes años y fechas de siembra en el norte de Tamaulipas, México.

