










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.8 no.2 Texcoco Fev./Mar. 2017
https://doi.org/10.29312/remexca.v8i2.51
Articles
Reproductive characteristics of nine varieties of poinsettia (Euphorbia pulcherrima, Willd. ex Klotzch)
1Posgrado en Ciencia Agropecuarias y Desarrollo Rural, Facultad de Ciencias Agropecuarias. Universidad Autónoma del estado de Morelos. Av. Universidad 1001. CP. 62209. Tel. (01) 777 3297046. Chamilpa, Cuernavaca. Morelos. (jesusvargasar@gmail.com; maria.andrade@uaem.mx; voscar66@yahoo.com.mx; acastillo-g@hotmail.com; iran.alia@uaem.mx).
2Departamento de Fitotecnia, Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5. CP. 56230. Tel. (01) 595 9521500. Chapingo, Estado de México. (lozcol@gmail.com; edivitia@hotmail.com).
Mexico is the center of origin of the poinsettia (Euphorbia pulcherrima, Willd. ex Klotzch), however, production depends on the varieties generated in other countries because they do not have national varieties. For genetic improvement of any kind study of reproductive biology is necessary. Based on the above, the objective of this research was to evaluate the reproduction of three varieties of poinsettia of sun (Bethlehem, John Paul, Christmas Dawn) and six cultivars of shadow (Freedom, Prestige, Ice Punch, Nutcracker white, Sonora White glitter and Burgundy) to define the bases of breeding methods. The viability of pollen was determined by staining with aceto-carmine and pollen germination; the schedule was studied when the flowers were receptive, applying a drop of H2O2 on stigma; four treatments were established to determine the type of pollination of the poinsettia plants. The research was developed in a completely randomized design. An analysis of variance and mean comparison test with Tukey test (p≤ 0.05). Was performed. The viability of pollen varied from 68 to 93%. The greater receptivity of the stigma was observed at 14:00 h. The sun varieties could act as females in a breeding program because of greater viability of pollen, receptive flowers, and fruit binding. The Nutcraker White and Prestige varieties showed no stigma receptivity or fruit binding. It was determined that the pollination type of the poinsettia was crossed.
Keywords: poinsettia; stigma receptivity; type of pollination; pollen viability
México es el centro de origen de la nochebuena (Euphorbia pulcherrima, Willd. ex Klotzch), no obstante, la producción depende de las variedades generadas en otros países porque no se tienen variedades nacionales. Para el mejoramiento genético de cualquier especie es necesario el estudio de la biología reproductiva. Con base en lo anterior, el objetivo de la presente investigación fue evaluar la reproducción de tres variedades de nochebuena de sol (Belén, Juan Pablo, Amanecer navideño) y seis cultivares de sombra (Freedom, Prestige, Ice punch, Nutcracker white, Sonora White glitter y Burgundy) para definir las bases de métodos de mejoramiento genético. Se determinó la viabilidad del polen mediante tinción con aceto-carmín y germinación de polen; se estudió el horario en que las flores estuvieron receptivas, aplicando una gota de H2O2 sobre el estigma; se establecieron cuatro tratamientos para determinar el tipo de polinización de las plantas de nochebuena. La investigación fue desarrollada en un diseño completamente al azar. Se realizó un análisis de varianza y una prueba de comparación de medias con la prueba Tukey (p≤ 0.05). La viabilidad del polen varió de 68 a 93%. La mayor receptividad del estigma se observó a las 14:00 h. Las variedades de sol podrían fungir como hembras en un programa de fitomejoramiento por tener mayor viabilidad del polen, flores receptivas, y amarre de fruto. Las variedades Nutcraker white y Prestige no presentaron receptividad del estigma ni amarre de fruto. Se determinó que el tipo de polinización de la nochebuena fue cruzada.
Palabras clave: nochebuena; receptividad del estigma; tipo de polinización; viabilidad del polen
Introduction
The poinsettia (Euphorbia pulcherrima, Willd. ex Klotzsch), also known as easter, christmas flower, christmas star or poinsettia, is native to the region from the northern state of Morelos to the municipality of Taxco, Guerrero (Trejo et al., 2012). Although Mexico is the center of origin of this plant, the production of poinsettia depends on the varieties generated in other countries. The new varieties that arrive in Mexico have been released at least two years earlier in the country where they were generated. The payment of royalties for the use of these new genotypes is approximately 10% of the cost of each plant, which is included in the sale price of the cutting.
The plants of the genus Euphorbia are characterized by having a female flower without petals and sepals usually without surrounded by male flowers, all in a cup-shaped structure called ciato hermaphrodite; they can also have female and male ciato. In the inflorescences there are modified leaves called bracts that can be red, pink and white (Recanses and Flores, 1983), in different shades or combinations of colors.
The studies related stigma receptivity, pollen quality and type of pollination of the species are of great importance in research related to sexual reproduction, as they allow ensure the success of the hybridizations and increasing efficiency improvement (Weller et al., 2007).
The stigmatic receptivity is a floral parameter that determines the time in which stigmas maintain the ability to generate an environment conducive to the germination of pollen grains, directly affecting reproductive success and impacting on the harvest when it comes to cultivated species; however, the factors that determine it are unknown (Lozada and Herrero, 2006). In nature, stigma receptivity is usually given in conjunction with flower anthesis (Shivanna and Sawhney, 1997), but in the case of breeding, the need to perform targeted pollinations in certain phenological states often prevails. The tests currently used primarily in fruit to observe the stigmatic consists of applying drops of hydrogen peroxide in the stigmatic wall, observing bubbling it indicates receptivity (Osborn et al., 1988).
The viability of pollen is a factor that affects the mooring of fruits; although there are methods to determine the in vivo pollen germination, viability and germination in vitro they have been widely used as alternative methods (Karapanos et al., 2010). The in vitro tests can be simple and quick to determine some aspects of the physiology of the pollen grain. The pollen viability Euphorbias was studied by Narbona et al. (2008) in E. nicaeensis, to compare the level of viability between hermaphrodites and male flowers. Regarding the germination of pollen tubes, Cavalari (2010) evaluated the germination of Euphorbiaceae in croton (Codiaeum variegatum) and could observe pollen grains germinated pollen tube with 7 mm. Similarly, mode Lyra et al. (2011) determined 17 and 30% germination of pollen grains for two species of Jatropha.
Although the study of reproductive biology is of fundamental importance in genetic improvement, on poinsettia it is almost nil. Huang and Chu (2008) report that evaluated fertility in poinsettia by pollen viability in vitro generated in order to improve results in their crossbreed’s plants. Based on the above, the objective of the present research was to evaluate the reproductive characteristics of three varieties poinsettia of sun and six shade cultivars, as a basis for genetic improvement.
Materials and methods
The poinsettia varieties of sun were studied Bethlehem, John Paul and Dawn Christmas; and cultivars shadow Freedom, Prestige, Nutcracker white, Ice Punch, Sonora white glitter and Burgundy (Figure 1), which were selected to present male and female reproductive structures (essential for carrying out the study), being of high economic importance, and the most commercialized in Mexico. In the nine poinsettia varieties the viability of pollen was studied by pollen tube germination and staining; were also determined the stigma receptivity, timing appropriate for pollination, as well as the type of pollination.

Figure 1 Shade poinsettia. A) Freedom; B) Prestige; C)Nutcracker white; D) Ice punch; E) Sonora white glitter; F) Burgundy; Sun poinsettia: G) Bethlehem; H) John Paul; I) Christmas dawn.
Viability of pollen by pollen tube germination
When the anthers were developed were observed with a magnifying glass and recorded the day when these were opened, when the pollen was fresh was taken and was grown in Petri dishes containing medium based on agar (4 g L-1) and sucrose (0.06 mol L-1). It was cultured in incubation at 36 °C; the number of germinated pollen tubes was observed at 3, 6, 12, 24, 48 and 72 h. The count of germinated pollen grains per box was determined to determine the percentage of germination and the time required to reach at least 1% of germination. The length of the pollen tubes was measured by using a (Carl Zeiss®) compound microscope observing 10 to 40 diopters, in order to know the time required for germination and pollen fertility measuring. A completely randomized design was used, with three replicates per variety, each repetition consisted of a Petri dish containing the three anther pollen grains taken at random.
Viability of pollen by staining
The viability of the pollen was also evaluated by staining with an acetic-carmine acid solution. In each plant, two anthers were collected per flower (before pollen release). These were placed in one tube per plant and fixed in Farmer solution (3:1, absolute ethyl alcohol, glacial acetic acid), in solution were stored in refrigeration until evaluation. For the study, an anther was placed on a slide, cut transversely, a drop of aceto-carmine dye was added and with the blade of a scalpel the anther was gently crushed to release the pollen into the dye drop, coverslip and allowed to elapse 5 to 10 s to observe the microscope ((40x) (Carl Zeiss®); for the pollen grain count seven preparations were made per plant, in each preparation four fields were observed in which the number of stained grains and the number of uncoloured pollen grains were evaluated. With these data the percentage of viability of pollen in each variety was determined. The study was conducted in a completely randomized experimental design with seven replications per variety.
Receptivity of stigma
Stigma receptivity was found by modifying the method of Osborn et al. (1988) which consisted of placing a drop of hydrogen peroxide (H2O2) of 80% of the stigmas and take a positive result bubble production, the adequacy of the method was performed because this concentration was greater reaction and stigma bubbling was more efficiently observed. This experiment was carried out in a completely randomized design with five replicates (plants) per variety, in each plant four inflorescences were taken.
Moment of pollination
Since there is no antecedent of the receptivity moment of the stigma, pollinations were carried out in three schedules of the day (8:00, 14:00, 18:00) taking as a parameter the amount of nectar present in the nectar of the hermaphrodite flowers, which was measured with a micro-pipette of 1 000 µL, being performed in the nine varieties, in four inflorescences of five plants. In addition, the test was performed Osborn et al. (1988), to know stigmatic receptivity. The synchronization was observed between the amount of nectar and the receptivity of the stigma. A completely randomized design with five replicates (plants) was used per variety, the receptivity of the stigma and volume of the nectar were evaluated.
Type of pollination
To determine the type of pollination of the poinsettia plants, four treatments were evaluated: 1) emasculate hermaphrodite flowers and cover completely to avoid the arrival of external pollen; 2) cover the female flowers and when they were receptive pollinate with pollen from another plant of the same variety and then re-cover, remove the male flowers as they appeared; 3) cover the female flowers and when they were receptive they were pollinated with pollen from the same plant and covered again; and 4) cover the female flowers and when they were receptive they were pollinated with flower pollen from plants of another variety and covered again. The experiment was developed in a completely randomized design, with five replicates (plant) per variety, pollinating 15 flowers per replicate. The percentage of mooring of fruit per plant was evaluated.
Analysis of data
In the five experiments, data were studied by analysis of variance, mean comparison test with Tukey test (p≤ 0.05) with program SAS 9.2 (SAS, 2002).
Results and discussion
Viability of pollen by pollen tube germination
In the poinsettia varieties the gynoecium style had a length of 6 mm (Figure 2). The pollen tube germination in vitro start at 6 h after initiating the culture but was up to 72 h when it reached 6 mm needed to transport the male gametes from the stigma to the ovary.
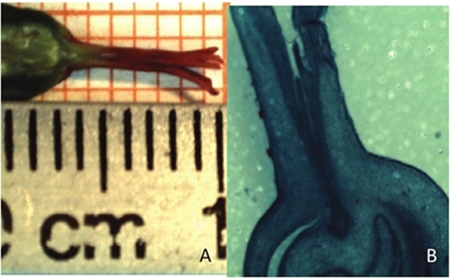
Figure 2 A) length of poinsettia style, measured with ruler and millimeter paper; and B) anatomical cut of a female flower where the length of the style is observed.
The analysis of variance showed highly significant differences in germination of the pollen tube of the nine varieties of poinsettia. The poinsettia of sun had the highest germination of pollen tube (>90%), four of the shade cultivars had 80-84% of viable pollen grains; Nutcraker white presented the lowest viability of pollen (Table 1). The above indicated that the varieties of sun could be good males, the opposite happened with Nutcracker white. These values were higher germination 30 and 17% observed by Lyra et al. (2011) in the study conducted with Jatropha molissima and J. ribifolia.Narbona et al. (2005) observed an average of 31.8% of pollen viability in E. boetica, which was also lower than what was observed in the present investigation.
Table 1 Viability of pollen by germination of pollen tube at 36 °C and by staining with acetic acid-carmine in nine varieties of poinsettia.
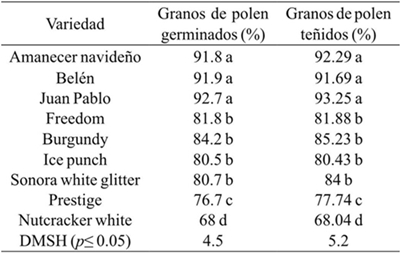
Valores con la misma letra son estadísticamente iguales de acuerdo con la prueba de Tukey (p≤ 0.05).
As for the time required for germination of pollen tube, Cavalari (2010) conducted a study in Euphorbia with Codiaeum variegatum, and observed that pollen grains of this genus reached maturity at 65 h in the medium of germination compared to the 72 h required by the poinsettia varieties of this research. This indicated that it was required about 72 h after pollination for fertilization to occur.
Viability of pollen by staining
In the preparations made to observe under the microscope two types of stains were presented, the grains of pollen that showed intense coloration were the viable grains, while the almost transparent pollen grains were the sterile grains (Figure 3). The viability of pollen by staining was higher (91 to 93%) in the sun varieties; four shade varieties had 80-85% viable pollen, while Prestige and Nutcraker white showed the lowest values (Table 1).
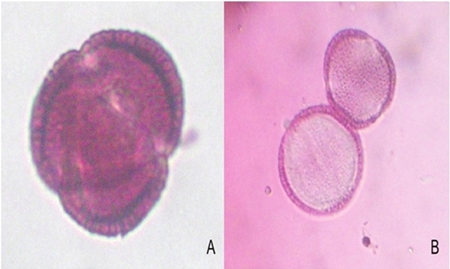
Figure 3 A) viable pollen grain on poinsettia; and B) nonviable pollen grains, observed at 10 diopters.
The viability of pollen by carmine staining was very similar to that obtained with the germination of the pollen tube; therefore, either methodologies were reliable to evaluate the male gametes (Table 1); given the above, it was considered that the assessment of viability by staining pollen grains was adequate because it is a simpler methodology.
Receptivity of stigma
The sunflower varieties Bethlehem, John Paul and Dawn Christmas had a higher percentage of flowers (43, 43.7 and 46.3) that presented bubbling, which showed the receptivity of the stigma (Figures 4 and 5) without differences between them. In contrast, shade varieties had a lower percentage of receptive flowers, of which Sonora White glitter was the one with the least receptivity (24%), whereas Prestige and Nutcraker white did not show bubbling in the stigma, indicating absence of receptivity.

Figure 4 Receptivity of the stigma on poinsettia. A and B) bubbling in the stigma. Osborn method (1988).

As for the time when the stigmas were receptive, it was observed that with the exception of Nutcraker white and Prestige, all the varieties had more receptive flowers at 14:00 h, in the majority of the varieties the less receptivity was to the 8:00 in the morning.
The results suggest that poinsettia was receptive at the time of day when the weather was warmer (14:00 h) (Figure 5), which could indicate that in hotter places, stigmatic receptivity could occur before the hour of the highest receptivity observed in this research, where the average annual temperature is 21.5 °C, according to the above, Campos (2013) observed that the best time to perform the pollination at Christmas in Zacatepec, Morelos (24 °C annual mean temperature ) were at 11:00 am as they obtained the highest number of fruits per plant (1.85) and seeds per fruit (2.56) compared to the pollination at 12:00 of the day, in which 1.1 fruits per plant were harvested and 0.83 seeds per fruit.
Considering the results obtained by Lozada and Herrero (2006), who mentioned that stigmatic receptivity directly affected reproductive success, the Prestige and Nutcracker white varieties that did not show receptivity to the stigma were not able to bear fruit, on the contrary, poinsettia of sun presenting a higher percentage of receptive flowers could form more capsules. Based on the above results, the sun varieties are the most viable candidates for being females in a genetic improvement program at poinsettia.
Moment of pollination
The receptivity of the stigma was lower at 8:00 am in the seven varieties that showed receptivity of the female organ, while it was higher at 2:00 pm, indicating that this time corresponded to the right moment to perform the pollination; the three varieties of poinsettia of sun were the ones with the highest percentage of receptive flowers (43 to 46%) (Figure 5). Prestige and Nutcracker white did not show stigma receptivity in any of the hour in which the evaluation was done, indicating that these two varieties could not function as female or male progenitors in a breeding program, since they also had the lowest percentage of pollen viability.
In all varieties, except for Nutcracker white and Prestige, it was observed that nectar began to flow from the nectaries in male flowers after pollen dehiscence; while the stigmatic receptivity test was positive three days after pollen dehiscence (Figure 6); the above indicated absence of synchronization in the maturity of male and female structures. This may be the reason why poinsettia is cross-pollinated, as Augspurger (1983) mentions reproductive success is influenced by the degree of synchrony of flowering events.
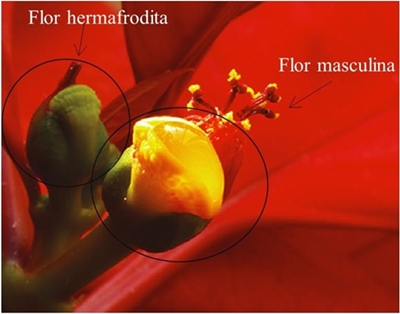
Figure 6 Floral asynchrony on poinsettia. Male flowers with dehiscence of pollen and nectaries, immature hermaphrodite flower, still does not present stigmatic receptivity
The amount of nectar was higher in sunshine varieties > 38 µL, while in the shade varieties it was 13.5-16.2 µL lower (Table 2). This characteristic was related to the receptivity of the stigma, which was also higher in the sun varieties (Figure 6). The volume of the nectar could be the parameter indicating the moment of pollination. However, the pollen dehiscence is more visible and the receptivity of the hermaphrodite flowers was observed three days after the release of the male gametes, pollen dehiscence being a better indicator to identify the right moment for pollination. In this regard, Vargas et al. (1999) noted that in E. heterophylla pollen release was greater with increasing temperature increase occurring after noon or late afternoon, this in Viçosa, Brasil.
Table 2 Volume of nectar in nine varieties of poinsettia.
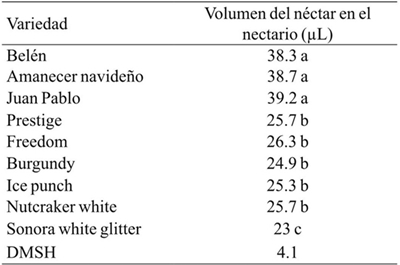
Valores con la misma letra son estadísticamente iguales de acuerdo a la prueba de Tukey (p≤ 0.05).
The adequate time for pollination is variable depending on the place, as the dehiscence of pollen grains was affected by temperature and relative humidity of cultivation, as reported by Lisci et al. (1994) who pointed out that the time of day for anther dehiscence varied over time and appears to be influenced by temperature and relative humidity.
During the development of the experiment, several pollinating insects were observed in the sun varieties between 12:00 and 16:00 h, when the nectaries had on average 35 µL of nectar; orders of insects that were observed were hymenoptera, diptera (family Tachinidae), coleoptera and lepidoptera monarch butterfly (Danaus plexippus) (Figure 7) the latter was the only observed in the varieties of shade.

Type of pollination
The treatment 1 that consisted of emasculating flowers and covering them to avoid the arrival of foreign pollen, there was no fruit formation, which indicated that poinsettia did not produce seeds by apomixis.
In treatment 2 that consisted of eliminating the male flowers as they appeared and pollinated the female flowers with pollen from another plant of the same variety, more than two fruits, having highly significant differences with the other varieties, which generated on average less than one fruit, which reinforced that poinsettia is a cross-pollinated plant.
In treatment 3 where the female flowers were covered and pollinated with flower pollen from the same plant and covered again, no fruit formation was observed which indicated that it was possible that on poinsettia there would be auto incompatibility of gametes.
Finally, in treatment 4 where the female flowers were covered and pollinated with flower pollen from another poinsettia variety, there was fruit formation, indicating that the type of pollination of the poinsettia was crossed, since most fruits were formed of the varieties, with the exception of Prestige and Nutcracker white. The highest number of fruits was observed in the sun, especially in Belen (2.6 fruits on average), while the varieties Freedom, Burgundy and Ice punch formed 0.85 fruits on average (Figure 8), the fruits were formed in the flowers hermaphrodites.

Conclusions
The sun varieties had greater viability of pollen, receptive flowers, and fruit binding, so they can serve as females in a breeding program Nutcraker white and Prestige are varieties that could not function as females because they did not present stigma receptivity or fruit binding. The greater receptivity of the stigma was at 14:00 h, which occurs in the period of activity of the pollinating insects. It was determined that the pollination type of the poinsettia is crossed.
Literatura citada
Augspurger, C. K. 1983. Phenology, flowering synchrony and fruit set of six neotropical shrubs. Biotropica. 15:257-267. [ Links ]
Campos, B. E. 2013. Hibridación en nochebuena de sol (Euphorbia pulcherrima Willd. ex Klotzsch) con la variedad Prestige early y su validación con marcadores morfológicos y moleculares. Tesis de Maestría en Ciencias Agropecuarias y Desarrollo Rural. Universidad Autónoma del Estado de Morelos. Morelos, México. 71 p. [ Links ]
Cavalari, P. O. 2010. Morfologia e desenvolvimento floral em croton l. e astraea klotzsch (Euphorbiaceae sensu stricto). Programa de pós-graduação em ciências biológicas. Universidade Estadual Paulista. 80 p. [ Links ]
Huang, C.H. and Chu, C. Y. 2008. Methods to improve poinsettia fertility. Scientia Horticulturae. 3:271-274. [ Links ]
Karapanos, I. C.; Akoumianakis, K. A.; Olympios, C. M. and Passam, H. 2010. Tomato pollen respiration in relation to in vitro germination and pollen tube growth under favourable and stress-inducing temperatures. Sex Plant Reproduction. 23:219-224. [ Links ]
Lisci, M.; Tanda, C. and Pacini, E. 1994. Pollination ecophysiology of Mercuralis annua L. (Euphorbiaceae), an anemophilous species flowering all year round. Ann. Bot. 74:125-135. [ Links ]
Lozada, J. M. y Herrero, M. 2006. Estudios de receptividad estigmática en manzano (malus*pumila, Mill.). Departamento de Pomología. 10-12 pp. [ Links ]
Lyra, D. H.; Sampaio, L. S.; Pereira, D. A.; Silva, A. P. and Amaral, C. L. F. 2011. Pollen viability and germination in Jatropha ribifolia and Jatropha mollissima (Euphorbiaceae): species with potential for biofuel production. Afr. J. Biotechnol. 10:368-374. [ Links ]
Narbona, E.; Ortiz, P. L. and Arista, M. 2005. Dichogamy and sexual dimorphism in floral traits in the andromonoecious Euphorbia boetica. Ann. Bot. 955:779-787. [ Links ]
Narbona, E.; Ortiz, P. L. and Arista, M. 2008. Sexual dimorphism in the andromonoecious Euphorbia nicaeensis: effects of gender and inflorescence development. Ann. of Bot. 101:717-726. [ Links ]
Osborn, M. M.; Kevan, P. G. and Lane, M. A.1988. Pollination biology of Opuntia polyacantha and Opuntia plaeacantha (Cactaceae) in southern Colorado. Plant Systematics and Evolution. 159:85-94. [ Links ]
Recanses, P. L. y Flores, L. I. 1983. Cultivo de la poinsettia. Hortic. Inter. 13:3-13. [ Links ]
SAS Institute, Inc. 2002. SAS user’s guide: Statistics. Version 9.00. SAS Inst., Inc., Cary, NC. USA. [ Links ]
Shivanna, K. R. and Sawhney, V. K. 1997. Pollen biotechnology for crop production and improvement. Cambridge University Press. Cambridge, UK. 28-29 pp. [ Links ]
Trejo, L.; Feria-Arroyo, T. P.; Olsen, K. M.; Eguiarte, L. E.; Arroyo, B.; Gruhn, J. A. and Olson, M. E. 2012. Poinsettia’s wild ancestor in the mexican dry tropics: historical, genetics, and environmental evidence. Am. J. Bot. 99:1146-1157. [ Links ]
Vargas, L.; Borém, A. y Da Silva, A. A. 1999. Técnica de cruzamientos controlados en Euphorbia heterophylla L. Bragantia, Campinas. 581:23-27. [ Links ]
Weller, S.; Domínguez, C.; Molina, F.; Fornoni, J. and Lebuhn, G. 2007. The evolution of distyly from tristyly in populations of Oxalis alpina (Oxalidaceae) in the sky islands of the Sonoran desert. Am. J. Bot. 94:972-985. [ Links ]
Received: January 2017; Accepted: March 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
