










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.8 no.1 Texcoco ene./feb. 2017
https://doi.org/10.29312/remexca.v8i1.78
Articles
Grain yield of Tuxpeño corn populations adapted to High Valleys de México
1Campus Montecillo-Colegio de Postgraduados. Carretera México Texcoco, km 36.5. Montecillos, Texcoco, Estado de México. CP. 56230.
2Universidad Autónoma Agraria Antonio Narro. Periférico Raúl López Sánchez y Carretera a Santa Fe, km 2, S/N. Torreón, Coahuila, México. CP. 27000.
3C. E. San Luis- INIFAP. Carretera San Luis-Matehuala, km 14.5. Ejido Palma de la Cruz, Soledad de Graciano Sánchez, San Luis Potosí, México. CP. 78431.
4C. E. Valle de México-INIFAP. Carretera Los Reyes-Texcoco, km 13.5. CP. 56250. Coatlinchán, Texcoco, Estado de México, México.
This work aimed to evaluate the agronomic behavior of maize (Zea mays L.) Tuxpeño adapted to High valleys. Four populations of Tuxpeño corn were evaluated in their versions of cycle one (misfit) and cycle seven (adapted): P1(C1), P1(C7); P2(C1), P2(C7); P3(C1), P3(C7), and P4(C1), P4(C7); four crosses of Chalqueño compounds 1, 2, 3, and 4 by P4 (C10), three crosses of populations P1, P2, and P3 by P4 (C10); and three witnesses: H-S2, Promesa and San Jose. The evaluation was done in the spring-summer cycle of 2013, in fields of the College of Postgraduates and the National Institute of Forestry, Agriculture and Livestock Research, Texcoco, State of Mexico, under a randomized complete block design with three replicates. The experimental plot had 26 mats of two plants each 50 cm in two rows of 6 m separated to 0.8 m. The grain yield of the materials and other agronomic characteristics were measured. The results indicated that exotic adapted populations yielded up to 6 t ha-1, significantly exceeding the original varieties, although they had a late vegetative cycle and were of lower height. Performance components had significant increases in populations; the crosses of Tuxpeño*Chalqueño yielded from 13 to 14 t ha-1, surpassing the commercial hybrids, San Jose and H-S2. This shows the existence of heterosis in the crosses and that tropical exotic corn adapted in High Valleys is a useful resource for genetic improvement.
Keywords: Zea mays L.; adaptation; crosses; selection; tropical maize
Este trabajo tuvo como objetivo evaluar el comportamiento agronómico de maíz (Zea mays L.) Tuxpeño adaptado a Valles Altos. Se evaluaron cuatro poblaciones de maíz Tuxpeño en sus versiones de ciclo uno (desadaptado) y ciclo siete (adaptado): P1(C1), P1(C7); P2(C1), P2(C7); P3(C1), P3(C7), y P4(C1), P4(C7); cuatro cruzas de los compuestos Chalqueño 1, 2, 3, y 4 por P4 (C10), tres cruzas de las poblaciones P1, P2, y P3 por P4 (C10); y tres testigos: H-S2, Promesa y San José. La evaluación se hizo en el ciclo primavera-verano de 2013, en terrenos del Colegio de Postgraduados y del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Texcoco, Estado de México, bajo un diseño experimental de bloques completos al azar con tres repeticiones. La parcela experimental tuvo 26 matas de dos plantas cada 50 cm en dos surcos de 6 m separados a 0.8 m. Se midió el rendimiento de grano de los materiales y otras características agronómicas. Los resultados indicaron que las poblaciones exóticas adaptadas rindieron hasta 6 t ha-1, superando significativamente a las variedades originales, aunque tuvieron ciclo vegetativo tardío y fueron de menor altura. Los componentes del rendimiento tuvieron incrementos significativos en las poblaciones; las cruzas de Tuxpeño*Chalqueño rindieron de 13 a 14 t ha-1, superando a los híbridos comerciales, San José y H-S2. Lo anterior, evidencia la existencia de heterosis en las cruzas y que el maíz exótico tropical adaptado en Valles Altos constituye un recurso útil para el mejoramiento genético.
Palabras claves: Zea mays L.; adaptación; cruzas; maíz tropical; selección
Introduction
The maize (Zea mays L.) is one of the world’s most widely distributed and productive crops (Poehlman, 2003). It is also the world’s largest crop and per capita consumption (Sleper and Poehlman, 2006) which ranks first in international agricultural production (Turrent et al., 2012). In Mexico, maize is the basic cereal in the food of the population and it is in this species that has the greatest genetic diversity for different environments and uses. Regarding their uses, 69% of the maize produced is destined for human consumption, 20% to the livestock sector; 10% for its industrialization, and 1% for seed production (Ortega and Ochoa, 2003).
Although Mexico has a wide variety of maize (Sánchez et al., 2000), the use of such diversity in breeding programs has been limited to a few local populations in each agricultural region (Molina, 1990; Castillo, 1993). The state of Mexico occupies the third place as a producer of maize (Ortega and Ochoa, 2003), where 98.5% of the crops are made mainly in the spring-summer cycle and the remaining 1.5% are carried out during the Autumn-Winter.
The state has an average yield of 2.7 t ha-1 (SAGARPA, 2013). Based on the magnitude of this average yield, it can be said that the production per unit area is low (Soto and Mijares, 2007) and it is necessary to increase it, but the area for the crop is becoming smaller and the demand continues to increase (Virgen et al., 2016). The alternative to increase the productivity of the local varieties of High Valleys maize is the genetic improvement, since in this region a high percentage of native maize is currently used and the production technology is deficient.
The local creole maize, which has been cultivated for several generations, is still used today by producers, for qualities such as adaptation, production and consumption characteristics and, in most cases, by tradition.
However, such materials are susceptible to genetic improvement, mainly by selection and hybridization or by the introduction of exotic alleles useful for yield, drought tolerance, resistance to post-harvest pests or higher nutrient content (Bergvinson et al., 2007).
The introduction of exotic genetic material to diverse environments, in addition to expanding the genetic base of local materials with new useful alleles, constitutes an alternative to increase genetic variability and maize productivity (García et al., 2002) in breeding programs. This could realize the goals of modern technology of using new improved varieties favored by producers, consumers and industry, but mainly of farmers who do not have the economic resources to buy hybrid or improved each year (Hallauer et al., 2010).
The exotic germplasm is generally not used per se, with adaptation problems being the main objection for use in local breeding programs; these problems of adaptation are mainly reflected in a high susceptibility to diseases, alteration of the vegetative cycle and decrease or no production of offspring (Molina, 1990). Therefore, before crossing exotic maize with local materials or using it per se, it is suggested to improve it by selection at low pressures for several cycles to adapt it and increase the frequency of its favorable alleles for yield and adaptation. Thus, the benefits of the tropical germplasm adapted by selection have been demonstrated in several studies in High valleys (Navas and Cervantes, 1991; Molina, 1993; Pérez-Colmenares et al., 2000, Gómez-Espejo et al., 2015) has demonstrated the benefits of the potential of tropical germplasm adapted by selection as a strategy to increase genetic diversity and productivity of local maize.
One of the first works of adaptation of exotic maize was made in the United States (Troyer and Brown, 1972), where, for the purpose of selection for performance and adaptation of exotic material, populations were obtained with adaptation to the US maize belt after 10 years of recombining Mexican races with local lines. In Mexico, temperate climate adaptation of tropical and subtropical climates has been proposed and achieved through the stratified visual mass selection developed by Molina in 1983 (Pérez et al., 2000; 2002; 2007).
Because yield is a measure of the reproductive capacity of plants (Hallauer et al., 2010), it is inferred that the degree of adaptation of exotic maize is related to its ability to produce a sufficient and robust offspring in the area where It was introduced. In this respect, the evaluation of exotic genotypes selected for adaptation in several settings is indispensable in genotechnical programs, since their relative response often changes from one environment to another. In this regard, from the agronomic point of view, Lin and Binns (1994) indicate that adaptability is the ability of plants to produce a good yield in a region different from that of their place of origin and to which they have been introduced. Thus, a cultivar is considered to be well adapted or with some degree of adaptation to a region if its yield is high relative to the rest of the local cultivars or the maladjusted introduced.
The present work had the objective of evaluating the yield and agronomic behavior of four Tuxpeño maize populations selected for adaptation to High V alleys, three crosses between some populations, and four crosses of a population with varieties of the local race Chalqueño. The hypothesis was that the selected populations surpassed in their performance and agronomic behavior to their respective original versions and interact in a significant way favorable to yield when being crossed with local corn Chalqueño.
Materials and methods
The genetic material evaluated consisted of four populations of Tuxpeño maize in their versions of cycle one (misfit) and cycle seven (adapted): P1(C1), P1(C7); P2(C1), P2(C7); P3(C1), P3(C7), and P4(C1), P4(C7), four crosses of Chalqueño compounds 1, 2, 3, and 4 by P4 (C10); of three crosses of populations P1, P2, and P3 by P4 (C10); and the commercial witnesses H-S2, Promesa and San Jose. The selection of the four populations of Tuxpeño corn was carried out in Montecillo, Texoco, State of Mexico. The evaluation of the genetic material was carried out under temporary conditions during the spring-summer cycle of 2013, in the experimental agricultural fields of the College of Postgraduates (CP) and the National Institute of Forestry, Agriculture and Livestock Researches (INIFAP). Both located in the municipality of Texcoco, State of Mexico, whose climate is temperate, with annual average temperature of 15 °C and annual rainfall of 645 mm, at an altitude of 2 240 m.
The experimental design in the evaluation of the materials was of complete random blocks with three replicates. The sowing of the varieties was done manually by depositing two seeds per bush every 50 cm, in experimental plots of two rows of 6 m in length with 0.8 m spacing between rows. This gave a total of 26 trees and 52 plants in a plot of 4.8 m2, equivalent to a density of 60 000 plants ha-1. At the time of sowing germination was given in both localities and in the rest of the crop cycle the humidity came from the rain.
The variables studied were: grain yield (REN, kg ha-1), days at male (FM) and female (FF) flowering, recorded as the number of days since sowing until 50% of the plants of each plot was found in anthesis and presence of exposed stigmas, respectively; height of plant (AP, cm) and cob (AM, cm), which were measured from the base of the plant to the apex of the panicle or cob, respectively; diameter of cob (DM, cm), number of rows per ear (NH), grains per row (GH), and ear length (LM, cm). The prolificacy index (IP) % was also calculated on each plot by dividing the harvested ears on the number of plants. With the exception of FM, FF and REN, the variables were measured in a random sample of 10 plants or 10 cobs of each plot, and averaged per plant or ear. The REN kg ha-1 was estimated by the ratio of the surface area of the experimental unit to that of one hectare.
The analysis of combined variance of the data of the variables of the two localities was done first considering the total of genetic material (genotypes), then this source of variation was divided into six groups of materials: compounds of C1, compounds of C7, crosses Of Tuxpeño*Chalqueño, crosses of Tuxpeños, witnesses, and between groups of materials. The comparison of means of the variables was performed by the Tukey test (α= 0.05). All analyzes were done using the statistical package SAS version 9 (SAS Institute, 2003).
Results and discussion
The analysis of combined variance of the data (Table 1) detected highly significant and significant differences between environments for yield (REN) and components length (LM) and ear diameter (DM) and prolificacy index (IP); there was also significance between environments for the agronomic variables days to female flowering (FF), male (FM) and height of cob (AM). The variables plant height (AP) and number of rows per ear (NH) and grains per row (GH) were not significant. The above result indicates that the average overall yield of all genetic materials behaved differently in each environment, and that the vegetative cycle averages and cob height of the genotypes were also different in each environment. The above due to environmental differences between test locations, such as soil type, precipitation, and the average differential response of genotypes to such environmental effects.

Table 1 Combined variance analysis of yield, yield components and agronomic variables of 18 varieties of exotic and local maize. Montecillo, Mexico.
The partition of the materials into groups of genotypes detected significance for REN and LM only between the advanced cycles (cycle 7) of the populations of Tuxpeño and between the crosses of the compounds of Chalqueño by the population 4, C10, which indicates that there were differences between Tuxpeño genotypes adapted on the one hand, and between crosses of local material adapted by exotic tropical material adapted by the other (Table 1). On the other hand, there was a significance between the initial cycles (cycle 1) of the Tuxpeño populations only for the AP and AM variables, indicating that the exotic tropical mismatched materials presented variability in plant size. The corresponding partition groups (GRP) materials found highly significant differences between genetic materials for all variables, except for IP, which finds that there were phenotypic and genotypic differences between the varieties tested.
The genotype interaction by environment was significant and highly significant for most variables, but mainly for the performance variables and some of their component variables, highlighting LM, DM and GH, indicating that some genotype or several of them had a response different in their agronomic behavior through the selection process and the interaction with the test environments. This is a result of the intrinsic genetic differences between the materials and the environmental effects different from those that were submitted in each selection cycle.
The comparison of means between genotypes (Table 2) shows that genotypes with lower yields were the initial cycles of the four Tuxpeño populations, whose yields ranged from 1.5 to 2.21 t ha-1, with no significant differences between them is attributed as a response to their high maladaptation to test environments. On the other hand, the advanced cycles of the populations of Tuxpeño had an intermediate yield in comparison with the commercial controls, which varied from 5.53 to 7.91 t ha-1, the latter being significantly different and corresponding to the population one cycle seven, And although these advanced cycles did not match any of the witnesses, they did exceed the average yields of maize in the Valley of Mexico. These results confirm that through visual mass selection it is possible to adapt Tuxpeño tropical maize to the High Valleys of México, since according to the criteria of Hallauer et al. (2010) and Lin and Binns (1994) can be considered that Tuxpeño cycle 7 populations already have a degree of adaptation to the temperate climatic conditions of High Valleys, since their performance is high in relation to the rest of the disadapted cycles.
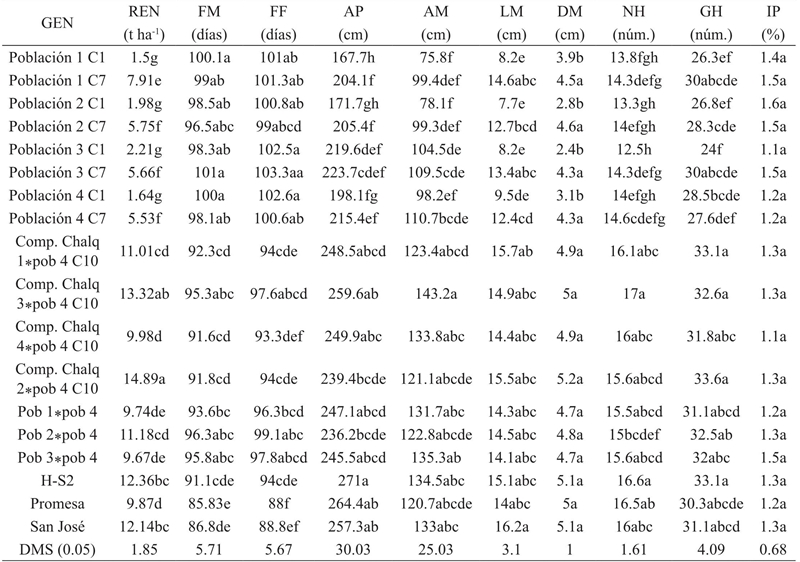
Table 2 Comparison of means of 18 maize genotypes for ten agronomic variables. Combined analysis. Montecillo, Mexico.
On the other hand, the crosses of Chalqueño (CCH) with the population four cycle 10 (Pop. 4 C10), and the crosses of the populations 1, 2, and 3 by the Pob. 4 C10 had some of the highest yields. Thus, the crosses of compounds Chalqueño 2 and 3 by Pob. 4 C10 had the highest yield (14.89 and 13.32 t ha-1, respectively), surpassing or equalizing the yield of all the controls and other crosses, which shows the importance of the adapted exotic material when crossing local High Valleys maize.
With respect to the crosses between populations of Tuxpeño, the three crosses evaluated statistically equated the witness Promesa, which yielded 9.87 t ha-1, but in particular the cross Pob. 2*Pop. 4 C10 had a yield of 11.18 t ha-1, surpassing or statistically equaling all the witnesses and some of the crosses of Chalqueño by Tuxpeño. These results are interesting, since the presence of an excellent behavior of the intervarietal cross by Tuxpeño adapted by Tuxpeño and of the intra-varietal cross Tuxpeño population 2 cycle seven by Tuxpeño population 4 cycle ten is verified. This can be attributed to the existence of heterosis in the crosses, which occurs when the hybrid overcomes its parents in phenological characteristics of growth and yield (Ramírez et al., 2007). The high performance of the advanced cycles of Tuxpeño was mainly influenced by the increases in the variables of its components, LM, DM, NH and GH during the selection process. On the other hand, the characteristics AP and AM were significantly reduced, which in Tuxpeño corn in particular is considered favorable from the agronomic point of view, since in its region of origin this variety presents a very high plant load, in some cases reaching more than two meters in height.
In agreement with these results and with the indicated by Lin and Binns (1994), it is possible to establish that the populations of Tuxpeño of this work selected in the region of High Valleys have a high degree of adaptation to conditions of temperate climate, since from the agronomic point of view, it can be defined that an exotic cultivar is adapted when its average yield is superior, or at least equal or close, to the rest of the local cultivars of the region. In this respect, having exotic germplasm in the High Valleys of Mexico, different from the region, would be advantageous, since these genotypes can be used per se, completing their cycle in the period and under the same conditions as local materials and commercial hybrids, obtaining considerable yields with respect to the average of production in the region. In addition, these exotic materials can broaden the genetic base of local varieties by providing new favorable alleles to increase their productivity (Holland, 2004).
With respect to the variables male and female flowering of Tuxpeño populations of adapted cycle, these did not present significant differences with respect to the original populations, since both presented blooms of 96 to 100 days, respectively. On the other hand, the crosses of Tuxpeño populations by Tuxpeño, and the crosses of Chalqueño compounds by Tuxpeño presented from 90 to 96 days of male and female flowering and even matched the commercial hybrids, so they can be classified as intermediate, according to the classification proposed by Pérez et al. (2007). This is important because these varieties and crosses may be suitable for irrigation or good temporary plantings in High Valleys. When comparing plant height and ear height, Tuxpeño crosses with local populations and commercial hybrids were the materials with the highest average values of 120 and 236.2 cm, respectively, significantly exceeding the values of these variables in the original populations (C1) and adapted 40%, since the original populations presented smaller plant size.
With respect to the cultivation cycle, the adapted populations of Tuxpeño reduced the days to male and female flowering and increased plant height per cycle of selection, but not equal to the local controls, reason why they are more precocious than the local material evaluated. On the other hand, crosses of compounds*populations, populations*populations, and controls had large ears with values of high yield components: ear length (LM) of 140.6 to 162.1 cm, ear diameter (DM) of 4.3 to 5.2 cm, number of rows (NH) of 15.0 to 17 cm, and grains per row (GH) of 30.3 to 33.6. The crosses of the compounds Chalqueño 2 and 3 by Pob. 4 C10 had the highest values for DM with 5.2 cm, NH of 17 and GH of 33.6. These results indicate for the compounds of Chalqueño*Tuxpeño that in their crosses there were positive interactions for yield between the Tuxpeño genes adapted with those of Chalqueño, which could be useful in the genetic improvement because these crosses are competitive with the commercial hybrids control used.
In comparing the adapted populations (C7) with the original populations (C1) differences between materials for variables LM, NH and GH, where generally adapted populations showed larger cobs, with length 12.4 to 13.4 cm were detected and cob diameters similar to those of commercial hybrids, with averages of 4.3 to 5.2 cm, higher than the values of the original populations, with number of rows and grains per row ranging from 14 rows to 30 grains.
The above results indicate that the populations of Tuxpeño cycle 7 (adapted) already have a higher frequency of genes favorable to yield and its components than the original varieties from which they were derived, due to the effect of the mass selection practiced in the populations during several years. These results also reflect in a certain way the degree of adaptation achieved by the plants of each compound in each selection cycle, mainly for yield, since this was the criterion of selection and interest. These results agree with Morales et al. (2007), who indicate that grain yield and its component variables are the most important to define the difference between genetic and phenotypic variability between original and adapted genetic materials. Finally, in relation to the variable prolificacy index (IP), no differences were detected between the materials, which is attributed to the cycle compounds one of the populations of Tuxpeño (maladjusted) already have several cycles of seed increase in Montecillo, which is why they have some adaptation, although not complete, and their plants produce ears; in addition, all materials had 1 to 2 ears per plant.
Conclusions
The results of the present investigation indicated that the exotic populations of Tuxpeño maize adapted by selection to the conditions of the High Valleys of Mexico increased their yield of grain significantly by cycle of selection with respect to the original cycles, this mainly due to the increase that they had (DM), number of rows (NH) and grains per row (GH).
The crosses of Chalqueño maize with Tuxpeño Population 4 cycle 10, and the cross between the populations of Tuxpeño 2 cycle 7 and 4 cycle 10 had some of the highest yields, surpassing the yield of the other materials and in some cases equating the yield of the witnesses, confirming that there were good behaviors of some interchannel crosses between local Chalqueño by adapted Tuxpeño, and of the intra-varietal cross Tuxpeño population 2 cycle 7 by Tuxpeño population 4 cycle 10. The results obtained can be attributed to the existence of heterosis in the crosses of local populations by adapted populations.
It was verified that the Tuxpeño tropical maize selection for adaptation to temperate conditions of High Valleys in Mexico was effective in increasing the frequency of favorable alleles for yield and adaptation in the selected populations. The use of exotic maize adapted in High Valleys is a useful resource for the genetic improvement of local maize and especially for producers, since the adapted tropical germplasm could be used per se or combined with local Chalqueño maize to take advantage of its heterosis in yield.
Literatura citada
Bergvinson, D. J.; Ramírez, A.; Flores, D. y García, L. S. 2007.Mejoramiento de maíces criollos por integración de alelos. México, D. F. CIMMYT 2008. Boletín: impulso a la producción de maíz en el Estado de México. 5(2):24-26. [ Links ]
Castillo, F. 1993. La variabilidad genética y el mejoramiento de los cultivos. Ciencia. 44:69-79. [ Links ]
García, J. J.; López, R. J.; Molina, G. T. y Cervantes, S. 2002. Selección masal visual estratificada y de familias de medios hermanos en una cruza intervarietal F2 de maíz. Rev. Fitotec. Mex.25:387. [ Links ]
Gómez, E. A. L.; Molina, G. J. D.; García, Z. J. J.; Mendoza, C. Ma. del C. y de la Rosa, L. A. 2015. Poblaciones exóticas originales y adaptadas de maíz. I: variedades locales de clima templado x variedades tropicales. Rev. Fitotec. Mex. 38(1):57- 66. [ Links ]
Hallauer, A. R.; Carena, J. M. and Miranda, F. J. B. 2010. Quantitative genetics in maize breeding. Iowa State University Press. USA.663 p. [ Links ]
Holland, J. B. 2004. Breeding: incorporation of exotic germplasm. In:encyclopedia of plant and crop science. Dekker, M. (Eds). New York, USA. 222-224 pp. [ Links ]
Lin, C. S. and Binns, M. R. 1994. Concepts and methods for analyzing regional trial data for cultivar and location selection. Plant Breed. Rev. 12:271-297. [ Links ]
Molina, G. J. D. 1990. Uso potencial de plasma germinal exótico de maíz. In: Resumen XIII Congreso Nacional de Fitogenética.Cd. Juárez Chihuahua, México. 344 p. [ Links ]
Molina, G. J. D. 1993. Comentarios a la plática. La variabilidad genética y el mejoramiento de los cultivos. In: Ciencia. Revista de la Academia de la Investigación Científica. México. Núm.Especial. 81-83. pp [ Links ]
Morales, R. M. M.; Parra, R. J.; Sánchez, G. J.; Ramírez, D. L.; De la Cruz, Mena, M. L. S.; Hurtado, P. S. y Chuela, B. M. 2007.Relaciones fenotípicas y heterosis entre híbridos comerciales y germoplasma exótico de maíz en Jalisco, México. Rev.Fitotec. Mex. 30(3):285-294. [ Links ]
Navas, A. A. A. y Cervantes, S. T. 1991 Selección para rendimiento y adaptación a Valles Altos en cruzas interraciales tropicales de maíz de Mexico. Agrociencia. 2(4):97-113. [ Links ]
Ortega, R. C. y Ochoa, B. R. 2003. El maíz: un legado de México para el mundo. Claridades Agropecuarias. 3-16 pp. [ Links ]
Pérez, C. A. A; Molina, G. J. D. y Martínez, G. A. 2000. Adaptación a clima templado de una variedad de maíz tropical mediante selección masal visual estratificada. Agrociencia. 34:533-542. [ Links ]
Pérez, C. A. A; Molina, G. J. D. y Martínez, G. A. 2002. Adaptación a clima templado de razas tropicales y subtropicales de maíz de México por selección masal visual. Rendimiento, altura de planta y precocidad. Rev. Fitotec. Mex. 25(4):435-441. [ Links ]
Pérez, C. A. A.; Molina, G. J. D.; Martínez, G. A.; García, M. P. y Reyes, L. D. 2007. Selección masal para la adaptación a clima templado de razas tropicales y sub-tropicales de maíz de México. Bioagro. (19(3):133-141. [ Links ]
Poehlman, J. M. y Allen, S. D. 2003. Mejoramiento genético de las cosechas. Traducido por Guzmán, O. M. 2a (Ed.) Ed.LIMUSA. México, D. F. 509 p. [ Links ]
Ramírez, D. J. L.; Chuela B. M.; Vidal, M. V. A.; Ron, P. J. y Caballero, H.F. 2007. Propuesta para formar híbridos de maíz combinando patrones heteróticos. Rev. Fitotec. Mex. 30:453-461. [ Links ]
SAGARPA (Secretaría de Agricultura, Ganadería, Desarrollo Rural,Pesca y Alimentación). 2013. Anuario estadístico de la producción agrícola de los Estados Unidos Mexicanos. http://www.siap.sagarpa.gob.mx. [ Links ]
SAS Institute 2003. SAS/IML Software: usage and reference. Version 9.1 Ed. SAS Institute Inc., Cary, N.C. [ Links ]
Sánchez, G. J. J.; Goodman, M. M. and Stuber, C. W. 2000. Isozymatic and morphological diversity in the races of maize of Mexico.Econ. Bot. 54:43-59. [ Links ]
Soto, R. y Mijares, P. 2007. Proyectos de investigación y transferencia de tecnología de maíz en el Estado de México. http://sedagrotecnologia.wordpress.com/2007/11/13/proyectosde-investigacion-y-tansferencia-de-tecnologia-de-maizenel-estado-de-mexico/. [ Links ]
Sleper, D. A. and Poehlman, J. M. 2006. Breeding field crops. Fifth Edition. Blackwell Publishing. Ames, Io. EE. UU. 424 p. [ Links ]
Turrent, F. A.; Wise, T. A. y Garvey, E. 2012. Factibilidad de alcanzar el potencial productivo de maíz en México. Mexican Rural Development Research Report. Reporte. 24-36 pp. [ Links ]
Troyer, A. F. and Brown, W. L. 1972. Selection for early flowering in corn. Crop Sci. 12:301-304. [ Links ]
Virgen, V. J.; Zepeda, B. R.; Ávila, P. M. A.; Espinosa, C. A.; Arellano,V. J. L. y Gámez, V. A. J. 2016. Producción y calidad de semilla de maíz en Valles Altos de México. Agron. Mesoam.27(1):191-206). [ Links ]
Received: January 2017; Accepted: March 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
