











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En los últimos años se han realizado muchos esfuerzos para incrementar el contenido de compuestos fenólicos y la capacidad antioxidante de plantas que vayan directo al consumo humano, empleando diversos métodos de cultivo tales como la aplicación de fertilización orgánica y convencional; en campo abierto y en invernadero (Kokaly y Skidmore, 2015). Sin embargo, los resultados han sido discrepantes acerca de la calidad de la planta y el rendimiento obtenido, concluyendo que el alto contenido de compuestos fenólicos y la capacidad antioxidante no es garantía de alto rendimiento de los cultivos (Salama et al., 2013). Esta circunstancia debe ser considerada en vista de la creciente población a nivel mundial, los altos costos de la producción de cultivos, y las cuestiones relacionadas con la contaminación ambiental, por lo cual se genera la necesidad de mejorar y seleccionar sistemas de producción más eficientes para mantener la producción de alimentos (Henderson et al., 2016).
Además de los impactos ambientales causados por los sistemas de producción agrícola, los efectos en la salud humana debido a nuevos hábitos de alimentación caracterizados por trastornos de la nutrición y el estrés, han aumentado el número de enfermedades crónicas como la obesidad, la diabetes, el cáncer, las enfermedades cardiovasculares y trastornos neurológicos entre otros (Lee y Scagel, 2009). Por lo tanto, es necesario unirse en un esfuerzo global en materia de alimentación, salud y medio ambiente, con el fin de producir alimentos seguros, que promueven la salud y reducen el riesgo de desarrollar enfermedades crónicas (Tilman y Clark, 2014). En este sentido, es importante proponer protocolos basados en sistemas biológicos controlando la aplicación de los nutrientes en soluciones nutritivas adecuadas para mejorar la calidad nutritiva y nutracéutica de las plantas (Kocira et al., 2016).
En este contexto, además de la obtención de cosechas de calidad en el contenido fenólico y capacidad antioxidante, es importante, obtener rendimientos aceptables para evitar pérdidas económicas, por lo que la producción de plantas medicinales y el control de los nutrientes en la solución nutritiva podría ser una alternativa a las cuestiones planteadas. En particular, el potasio (K) es uno de los nutrientes minerales requeridos en mayores cantidades por las plantas y tiene implicaciones fisiológicas en el mantenimiento celular y la apertura de los estomas (Nguyen et al., 2010). Además, el K promueve la rápida acumulación y translocación de carbohidratos y activa los procesos que son necesarios para la conservación del estado del agua de la planta y la presión de turgencia de las células (Pottosin et al., 2005). Este elemento es esencial para muchos procesos fisiológicos, tales como la fotosíntesis, la osmorregulación, la turgencia celular y la homeostasis en la célula vegetal (Mohd Zain e Ismail, 2016).
Algunos estudios señalan que el K, además de promover la síntesis de amidas y proteínas, es también un activador enzimático (Inthichack et al., 2012). Su función como activador enzimático se ha relacionado con la producción de fitonutrientes y por lo tanto, tiene implicaciones en la síntesis bioquímica de los productos del metabolismo secundario de las plantas (Mohd Zain e Ismail, 2016; Inthichack et al., 2012). Algunos autores señalan que el K favorece el aumento de los compuestos fenólicos (Ibrahim et al., 2012), los cuales son un gran y heterogéneo grupo de metabolitos secundarios, que se distribuyen en todo el reino vegetal (Abbasi et al., 2015). Los compuestos fenólicos se encuentran entre los fitoquímicos más deseables debido a su actividad antimicrobiana y antiviral; así como sus propiedades anti-inflamatorias y antioxidantes (Olennikov et al., 2011).
Por otra parte, una de las plantas reportadas ampliamente por su contenido de compuestos fenólicos y capacidad antioxidante es la albahaca (Ocimum basilicum L.), la cual es una especie ornamental, culinaria y medicinal perteneciente a la familia Lamiaceae, cultivada en todo el mundo y producida bajo una gran variedad condiciones de crecimiento (Kwee and Niemeyer, 2011).
De manera natural, la albahaca es una planta con una gran variedad de aplicaciones alimentarias y medicinales (Flanigan y Niemeyer, 2014; Koca y Karaman, 2015); sin embargo, existe poca información de que esta especie pueda incrementar su contenido fenólico y capacidad antioxidante sin que se vea afectado su rendimiento. Por lo tanto, el objetivo fue determinar un posible aumento del contenido en compuestos fenólicos, capacidad antioxidante y rendimiento, además de otros parámetros agronómicos como materia seca, altura de la planta y el número de hojas en tres genotipos de albahaca (Sweet Lemon, Red Rubin y Cinnamon) aplicando diferentes concentraciones de potasio.
Materiales y métodos
El trabajo se llevó a cabo en la Comarca Lagunera, ubicada entre los estados de Coahuila y Durango, México, durante la primavera de 2014 en la Localidad “El Quemado”, que pertenece al municipio de Gómez Palacio, Durango, México. El experimento se realizó en un invernadero con regulación de temperatura semiautomática, con valores entre 25-30 ºC y 70-80% para temperatura y humedad, respectivamente.
Se seleccionaron tres genotipos de albahaca (Ocimum basilicum L.): Sweet Lemon, Red Rubin y Cinnamon por representar una amplia gama de características morfológicas y genéticas (Lee y Scagel, 2009). Las semillas de albahaca se obtuvieron como una donación de la Universidad Juárez del Estado de Durango (UJED). La siembra se realizó en bandejas de germinación de poliestireno de 250 cavidades (tres semillas por cavidad) llenos de perlita (Environment®). Las bandejas se regaron dos veces al día con agua del grifo 21 días después de la siembra, las plántulas mostraron 3 a 4 hojas verdaderas y una altura de 0.1 a 0.15 m, por lo tanto, las plántulas se transfirieron a macetas de plástico negro (un galón de capacidad) que contenían una mezcla de arena: perlita (80:20).
La preparación de la solución de nutrientes se realizó utilizando el método universal de Steiner, que indica un equilibrio entre aniones y cationes (Steiner, 1961) (Cuadro 1).
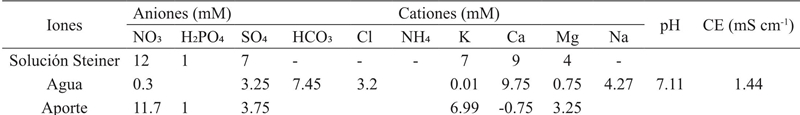
Por otro lado, en el Cuadro 2 se muestran los cálculos realizados para la aplicación de K (7, 9, 11 y 13 mM). Se utilizó KOH como fuente de potasio y las necesidades de los aniones y cationes se calcularon a partir del peso equivalente del fertilizante (Favela et al., 2006). Una vez que las plántulas se transfirieron y que las soluciones nutritivas fueron preparadas, las macetas fueron colocadas en un diseño factorial 3*4 de bloques completamente al azar con seis repeticiones por tratamiento (n= 72 plantas).
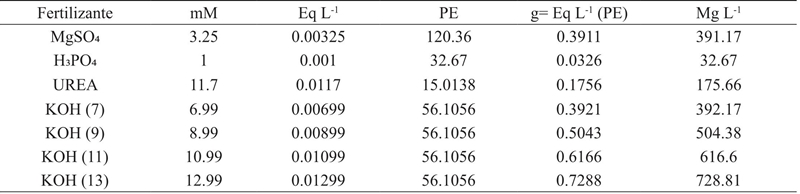
La aplicación de nutrientes se inició el día del trasplante y fue repetida dos veces al día (9:00 am y 6:00 pm). La cosecha se realizó 40 días después del trasplante. Las plantas cosechadas fueron trasladadas al Laboratorio de Bioquímica de la Universidad Politécnica de Gómez Palacio para su preparación. La albahaca fresca se lavó con solución de hipoclorito sódico al 3% y se colocó en papel marrón para su secado a temperatura ambiente (25 ± 2 ºC) durante 15 días.
Las muestras secas se molieron en una licuadora (Hamilton Beach) y se almacenaron a 5 °C para la posterior obtención de extractos. Se realizó una extracción sólido-líquido mediante la adición de 20 g de muestra en 100 ml de etanol. Se utilizó un agitador “Stuart” para mantener la mezcla bajo agitación durante 24 h a 30 ºC. Después de 24 h, la muestra fue llevada a secado a vacío y el etanol fue evaporado con rotavapor (Buchi, Modelo-210) y un baño de agua a 35-40 ºC. Los extractos obtenidos se almacenaron en refrigeración hasta su análisis respectivo.
El contenido fenólico total se midió usando una modificación del método de Folin-Ciocalteu (Esparza et al., 2006). 30 μl de extracto se mezclaron con 270 μl de agua destilada en un tubo de ensayo. En seguida 1.5 ml de reactivo de Folin-Ciocalteu (Sigma-Aldrich, St Louis MO, EE.UU.) diluido (1:15) se añadió a la mezcla, con agitación vortex durante 10 s. Después de 5 min se añadió 1.2 ml de carbonato de sodio (7.5% w/v) y se agitó durante 10 s. La solución se colocó en baño de agua a 45 ºC durante 15 min, y después se dejó enfriar a temperatura ambiente. La absorbancia de la solución se leyó a 765 nm en un espectrofotómetro UV Genesys 10. Se calculó el contenido fenólico mediante una curva de calibración utilizando ácido gálico como estándar, y los resultados se registraron en mg de equivalente de ácido gálico por 100 gramos de extracto (mg AGE/100 g de extracto).
La determinación de la capacidad antioxidante total de las diferentes muestras se llevó a cabo basándose en el método Brand-Williams et al. (1995) con ligeras modificaciones. La solución de radical libre 1.1-difenil-2-picrilhidrazil (DPPH) (Aldrich, St. Louis MO, EE.UU.) se preparó en un matraz completamente cubierto con papel de aluminio con DPPH+ (5 mg/100 ml de etanol grado analítico). La mezcla se agitó vigorosamente y el matraz se mantuvo cubierto para evitar la degradación rápida. 300 μl muestra del extracto diluido en tubos de ensayo por triplicado y 1 200 μl de agua destilada se agitó a 3 000 rpm durante 10 s. Se agregó 1 ml de DPPH+ y agitó con vórtex de nuevo a 3000 rpm durante 10 s. Las lecturas se realizaron a 517 nm después de 90 min. La capacidad antioxidante total se calculó utilizando una curva estándar con el antioxidante de referencia Trolox y los resultados se expresaron en μM Trolox por 100 gramos de extracto (μM Trolox/100 g extracto).
Después de la cosecha, la altura de la planta se midió desde la parte basal hasta la parte apical con una cinta métrica retráctil de 5 m de longitud (Moncayo et al., 2015). Posteriormente, las plantas se cortaron de la base para la medición del rendimiento. Los rendimientos se obtuvieron ponderando la planta en una báscula granataria (OHAUS) y reportados como kg por planta en base peso fresco (kg planta/PF). Para determinar el número de hojas, éstas se separaron de los tallos y cuantificadas. La materia seca (MS) se cuantificó de acuerdo con el método establecido por la AOAC (2005), colocando 12 g de la muestra fresca en cajas de aluminio en un horno de aire forzado a 70 °C hasta peso constante. Todos los análisis y las mediciones se realizaron por triplicado.
Los datos de las variables para factores e interacciones se analizaron mediante un análisis de varianza utilizando el software estadístico SAS (1999). Además, se obtuvo una correlación y regresión de los datos obtenidos. Para las comparaciones de los medios se utilizó la prueba de Tukey (p< 0.05).
Resultados y discusión
El interés por la determinación de los compuestos fenólicos en albahaca en este estudio fue dilucidar los mecanismos de biosíntesis como resultado del cambio en la concentración de potasio (K). Los resultados de este estudio mostraron diferencias altamente significativas (p≤ 0.001) para los compuestos fenólicos totales (CFT). Estos se vieron afectados por genotipo (G) y nivel de potasio (NK) (Figura 1). Como se puede ver en la Figura 1a Sweet Lemon cultivar mostró el contenido fenólico más alto (1 353 mg AGE/100 g de extracto) seguido de los cultivares Cinnamon y Red Rubin (1 251 y 1 230 mg AGE/100 g de extracto, respectivamente). Javanmardi et al. (2003) estudiaron los compuestos fenólicos totales en 23 cultivares diferentes de albahaca y reportaron valores de 22.9 a 65.5 mg AGE/g de peso seco (PS).
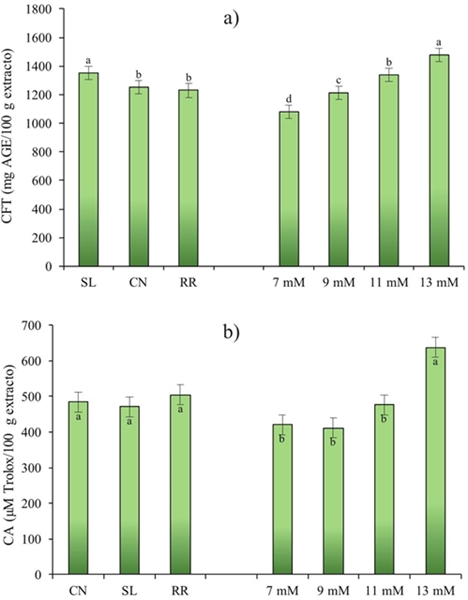
Figura 1 Efecto del cultivar (C) y nivel de K (NK) en el contenido de compuestos fenólicos totales (a); y capacidad antioxidante (b), de Ocimum basilicum L. CN= Cinnamon; SL= Sweet Lemon; RR= Red Rubin.
Por otra parte, Kwee y Niemeyer (2012) en un estudio de 15 cultivares de albahaca indicó rangos de 3.47 a 17.58 mg AGE/g PS. Por otro lado, Flanigan y Niemeyer (2014) en 10 cultivares encontraron valores de 13.1 a 26.9 mg AGE/g PS. Los autores mencionados indicaron que el contenido fenólico en las plantas esta determinado no solo por el cultivar, sino también por las condiciones y lugar de cultivo, además la concentración de fenólicos totales e individuales como flavonoides y ácidos fenólicos puede variar por dichos parámetros. De acuerdo con el factor NK, se observó que cuanto mayor es el nivel de potasio mayor el contenido de compuestos fenólicos totales. Así, se obtuvo el mayor contenido de compuestos fenólicos (1 480 mg AGE/100 g de extracto) con 13 mM (Figura 1a).
Estos resultados están de acuerdo con otros datos reportados en la literatura. Por ejemplo, Knuyen et al. (2010) evaluaron concentraciones de K en el intervalo de 1 a 5 mM y obtuvieron valores de compuestos fenólicos de 7 a 16 mg AGE/g PS. En otro estudio, Ibrahim et al. (2012) reportaron un incremento de 1.22 a 1.82 mg AGE/g de extracto mediante la aplicación de 0 a 270 kg de K ha-1 en Labisia pumila Benth. Estos resultados son coincidentes con este estudio debido a que confirman una correlación entre la absorción de potasio y el contenido de compuestos fenólicos en la planta (Cuadro 3) observándose una correlación positiva significativa en todos los cultivares (R2= 0.972) entre NK y CFT. Además, se encontró una interacción entre el genotipo y el nivel de potasio (G*NK) (Figura 2a). La interacción mostrada sugiere un aumento lineal en el contenido fenólico de los tres genotipos en respuesta al aumento del nivel de potasio. Generalmente, el ion K+ regula el equilibrio interno con el propósito de absorción y translocación de hidratos de carbono que afectan indirectamente la formación de las estructuras básicas de los fenilpropanoides (Kuum et al., 2015). Sin embargo, no es concluyente que el aumento del contenido de compuestos fenólicos en la planta pueda ser siempre lineal con el incremento de la aplicación de K, por lo tanto, podría ser interesante seguir investigando el rango de aplicación de K, con la posibilidad de encontrar una respuesta cuadrática para relacionar el nivel de K y la producción de compuestos fenólicos totales.
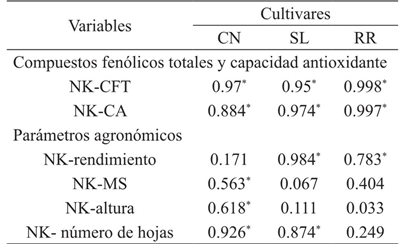
*= significancia a p≤ 0.05; CN= Cinnamon; SL= Sweet Lemon; RR= Red Rubin; CFT= compuestos fenólicos totales; CA= capacidad antioxidante; MS= materia seca.
Cuadro 3 Coeficiente de correlación R2 entre la concentración de potasio y el contenido de compuestos fenólicos totales, capacidad antioxidante y parámetros agronómicos en Ocimum basilicum L.
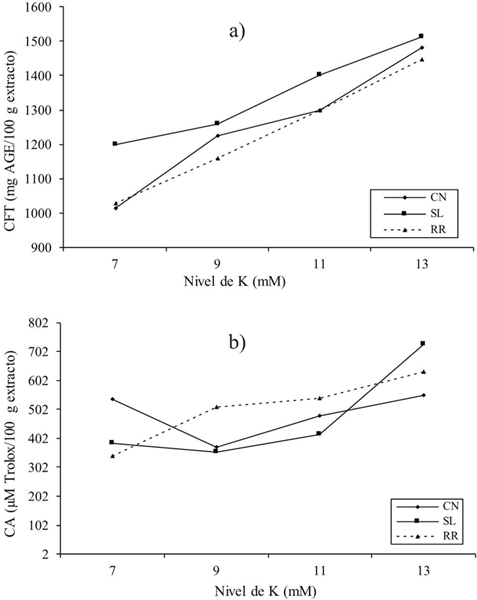
Figura 2 Interacción del cultivar y nivel de K (C*NK) para contenido de compuestos fenólicos totales (a); y capacidad antioxidante (b) de Ocimum basilicum L.CN= Cinnamon; SL= Sweet Lemon; RR= Red Rubin.
Respecto a la capacidad antioxidante (CA) en este estudio no se encontró diferencia significativa entre genotipos, pero si para el nivel de potasio (p≤ 0.05) (Figura 1b). La capacidad antioxidante de albahaca se ha reportado en numerosos estudios (Szymanowska et al., 2015), lo cual indica que independientemente del genotipo, la albahaca es una fuente potencial de compuestos bioactivos con capacidad antioxidante. Por otra parte, los resultados mostraron que la capacidad antioxidante de la albahaca fue mayor con el aumento del nivel de potasio, demostrándose esto con la correlación positiva significativa obtenida (R2= 0.951) entre NK y CA en los tres genotipos estudiados (Cuadro 3).
La interacción G*NK para compuestos fenólicos fue altamente significativa (p≤ 0,001) (Figura 2b), lo que indica que uno o más niveles de potasio utilizados afectan a los genotipos en estudio. Red Rubin mostró una respuesta lineal con el aumento de NK, aunque la respuesta fue similar en 9 y 11 mM, y se obtiene una mayor capacidad antioxidante en 13 mM. Mientras tanto Cinnamon presentó esta respuesta lineal en el intervalo de 9 a13 mM; sin embargo, es de interés observar que el nivel 7 obtuvo un valor similar al nivel 13. Por último, Sweet Lemon tiene capacidad antioxidante similar en el rango de 7 a 11 y aumenta considerablemente en 13 mM.
Por otra parte, se obtuvo una correlación entre los compuestos fenólicos totales y capacidad antioxidante (R2= 0.65) (Figura 3), lo que podría sugerir que 65% de la capacidad antioxidante obtenida es debido al contenido de compuestos fenólicos.
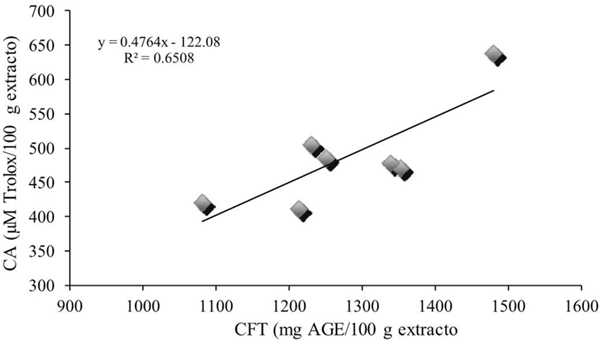
Figura 3 Correlación entre el contenido de compuestos fenólicos totales y la capacidad antioxidante de Ocimum basilicum L.
Los valores de rendimiento mostraron diferencias altamente significativas (p≤ 0.001) para G y NK (Figura 4a). El mayor valor de rendimiento se obtuvo con Cinnamon (0.938 kg planta-1 PF), seguido de Sweet Lemon y Red Rubin (0.9 y 0.87 kg planta-1 PF, respectivamente). Para el factor NK, los rendimientos fueron estadísticamente similares con los niveles de 9, 11 y 13 mM, el rendimiento más bajo fue de 7 mM.
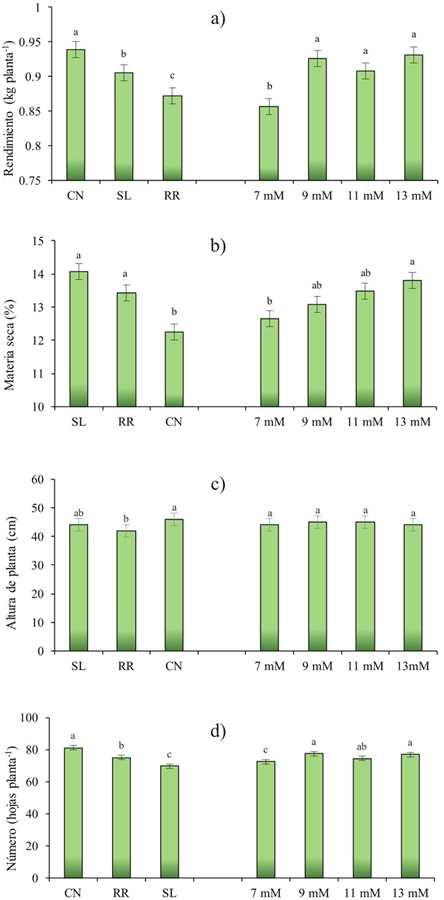
Figura 4 Efecto del cultivar (C) y nivel de K (NK) en rendimiento (a); materia seca (b); altura de planta(c); y número de hojas (d); de Ocimum basilicum L.CN= Cinnamon; SL= Sweet Lemon; RR= Red Rubin.
Para la altura de planta, el análisis de varianza mostró diferencias altamente significativas (p≤ 0.001) debido al genotipo (G), pero no hubo diferencia para el factor potasio (NK) (Figura 4c). Estos resultados podrían indicar que NK no afecta a la altura de la planta, pero esta variable está relacionada con el genotipo (Al-Kateb y Mottram, 2014), siendo un factor a considerar al elegir un cultivar con esta característica. Como se puede ver en la Figura 4c Cinnamon presenta altura mayor (46 cm), al lado de Sweet Lemon (44 cm) y estos son seguidos por Red Rubin (42 cm).
Además, se obtuvo una interacción altamente significativa (p≤ 0.001) para la altura de la planta debido a G y NK (G*NK) (Figura 4). La Figura 4c muestra que en Cinnamon la altura de la planta fue mayor en el rango de 9 mM y 11 mM y el más bajo en 7 y 13 mM. Sweet Lemon tienen valores similares en todos los niveles de K; y Red Rubin mostró valores similares en el rango de 9 a 13 mm y más altura en 7 mM. La altura es un parámetro que está influenciado por las características genéticas de la planta; sin embargo, sus propias características genéticas producen diferentes respuestas biológicas que definen un estado particular del organismo en un rango óptimo (Li et al., 2016).
Rubin es una variedad de albahaca ópalo oscuro, siendo un cultivo que tiene hojas de color rojizo-púrpura en color y un sabor más fuerte que la albahaca dulce (Bekhradi et al., 2015). Posiblemente su altura más baja podría relacionarse con una respuesta fisiológica y bioquímica en el metabolismo primario y secundario debido a la movilización y la translocación de los nutrientes en la planta, produciendo una menor altura y dirigir la respuesta a la concentración de pigmentos (Landi et al., 2013). El número de hojas resultó con diferencias altamente significativas (p≤ 0.001) para G y NK (p≤ 0.05) (Figura 4). La Figura 4d muestra que la Cinnamon presentó mayor número de hojas (81.67 hojas por planta) seguido de Red Rubin y Sweet Lemon (75 y 69 hojas por planta, respectivamente).
El número de hojas se encontró dentro de los rangos reportados en otros estudios, como lo indica Moncayo et al. (2015) en un estudio con albahaca blanca, donde los valores fueron de 66.5 a 103 hojas por planta.
En este estudio se observó que con 9 mM de K, se obtuvo más hojas por planta (77.73), seguido por el nivel de 13 mM (76.88) y 11 mM (74.33). Finalmente el número inferior de las hojas se obtuvo con 7 mM. Algunos autores sugieren que el desarrollo de la hoja está inicialmente relacionado con la aparición de la hoja primordial en el ápice, que puede depender de las características genéticas de la planta, a continuación depende de factores ambientales y nutricionales (Uzun, 2006). En un estudio en el melón se encontró que existen diferencias en el número de hojas por la concentración de K, indicando que la importancia fisiológica de este nutriente se encuentra en un área mayor de impacto para la fotosíntesis y, por tanto, una mayor producción de esqueletos de carbono, que utilizará o almacenará para más adelante y será transportado a los sitios de la demanda (Preciado et al., 2002).
El efecto del potasio en el número de hojas se puede demostrar mediante la correlación obtenida (Cuadro 2) en Cinnamon y Sweet Lemon donde se podría sugerir que 92 y 87% (respectivamente) del número de hojas en estos cultivares es debido al aumento en la concentración de potacio.
Conclusiones
En este trabajo se realizó una manipulación de la concentración de potasio en la solución de nutrientes para incrementar la producción de compuestos fenólicos y la capacidad antioxidante de albahaca como una opción para crear valor añadido al cultivo. Se demostró que el contenido de los compuestos fenólicos y la capacidad antioxidante de la albahaca podría mejorarse mediante la selección de cultivares y aumentando la concentración de potasio durante el crecimiento de la planta. Se obtuvo una correlación lineal significativa entre la concentración de potasio aplicado y el contenido total de compuestos fenólicos y la capacidad antioxidante para los tres genotipos de albahaca investigados. Para el rendimiento se concluye que los resultados muestran que este es afectado por el incremento en la concentración de potasio y que varía por la selección del genotipo.

