











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La radiación solar es un factor clave para el funcionamiento de los ecosistemas terrestres y acuáticos a través del control de procesos fotobiológicos, factores ambientales y ciclos naturales (Carrasco-Ríos, 2009; Bornman et al., 2015). Según Troy y Thennadil (2001), el espectro electromagnético que alcanza la superficie terrestre incluye a la radiación UV (200- 400 nm), a la fotosintéticamente activa (400-700 nm) e infrarroja cercana (800-2 500 nm).
Convencionalmente el espectro de radiación UV se ha dividido en tres componentes. El denominado UV-A comprende las longitudes de onda de 320 a 400 nm, el UV-B de 280 a 320 nm y el UV-C de 200 a 280 nm (Carrasco-Ríos, 2009). La radiación UV-A atraviesa la atmósfera y alcanza la superficie terrestre; la radiación UVB es absorbida en su mayor parte por la capa de ozono atmosférica, por lo tanto solo una pequeña cantidad de ella alcanza la superficie terrestre. Del total de radiación en la superficie terrestre la UV-B contabilizó solo 1.5%. La radiación UV-C es totalmente absorbida por el ozono atmósférico, por ello no se encuentra de forma natural en los ecosistemas (Nawkar et al., 2013).
Las plantas utilizan la radiación electromagnética como información para ajustar su crecimiento y desarrollo (Kami et al., 2010). Para ello utilizan proteínas sensoras para crear una conexión entre el estímulo ambiental y las respuestas fisiológicas como el rompimiento de la dormancia, la velocidad de germinación y las respuestas pos germinativas del embrión, permitiendo que las señales ambientales sean colectadas y procesadas inclusive por las semillas aparentemente inactivas que forman parte de un banco de semillas o de germoplasma. Este proceso de percepción y señalización de la radiación electromagnética por parte de una semilla parece estar mediado por diferentes receptores como los fitocromos (Magliano y Casal, 2004), y es muy importante desde el punto de vista de la oportunidad y capacidad de supervivencia del embrión una vez que la semilla germina (Long et al., 2015). En el caso particular de la radiación UV-B el fotoreceptor es llamado UVR8 (Kami et al., 2010; Heijde y Ulm, 2012).
A diferencia de la radiación UV-B, hasta el momento no se ha encontrado un receptor específico para la radiación UV-C, tal vez porque esta de forma natural no llega a la superficie terrestre ya que es bloqueada completamente por las moléculas de O3 de la atmosfera. La radiación UV-C ha sido investigada mayormente en cuanto a los efectos de su aplicación poscosecha en frutas, hojas y hortalizas. Se ha encontrado disminución de síntesis de proteínas, retardo en la maduración, incremento en niveles de flavonoides, acumulación de fitoalexinas y la inducción de resistencia, encontrándose también respuestas negativas en los procesos celulares, el metabolismo y crecimiento (Promyou y Supapvanich, 2012).
Tanto para la radiación UV-B como la UV-C se ha estudiado poco la utilidad de la aplicación en la irradiación de las semillas, ello a pesar de que teóricamente los cambios inducidos en la semilla por la irradiación pudieran hacerse patentes en etapas posteriores del crecimiento (Magliano y Casal, 2004; González-Aguilar et al., 2007) por contar las cubiertas de las semillas y las capas celulares subyacentes con una gran cantidad de receptores, proteínas y compuestos señalizadores (Moïse et al., 2005).
Se sabe que en el girasol y la soya las semillas irradiadas con UV-C muestran respuestas positivas frente al estrés abiótico (Foroughbakhch et al., 2015), en el melón la irradación de las semillas con UV-B causó cambios en la morfología y composición química de las plantas (Sosa-Flores et al., 2014) mientras que en el repollo la irradiación de las semillas causó tolerancia frente al estrés biótico en las plantas adultas (Brown et al., 2001).
Sería deseable aplicar esta misma técnica en especies hortícolas tan importantes como el tomate; sin embargo, no se dispone de información acerca de la respuesta germinativa, del vigor de las plántulas y del crecimiento de las plantas de esta especie cuando las semillas son sometidas a irradiación.
El objetivo de este estudio fue comprobar la factibilidad de utilizar la irradiación de semillas de tomate con UV-B y UV-C como una herramienta biofísica para modificar tanto las respuestas en germinación y vigor como el crecimiento de las plantas.
Materiales y métodos
El experimento se llevó a cabo en las instalaciones de la Universidad Autónoma Agraria Antonio Narro, en Buenavista Saltillo Coahuila, México, ubicada a 25° 21’ 19” latitud norte y a 101° 01’ 48” longitud oeste a una altura de 1 779 msnm. El material vegetal utilizado fue tomate (Solanum lycopersicum L.) híbrido Big Rio. El experimento comprendió dos etapas: en la primera etapa se determinaron a nivel de laboratorio las pruebas de germinación y vigor de las plántulas obtenidas de las semillas irradiadas con varias dosis de radiación UV-B y UV-C. En base a los resultados obtenidos de la primera etapa se eligieron las mejores dosis para irradiar las semillas a utilizarse en la segunda etapa.
La primera etapa se inició con la irradiación de las semillas el día 7 de noviembre de 2014 usando una cámara de irradiación con una lámpara modelo 3UV-36, la cual emite radiación en diferentes rangos: UV-A, UV-B y UV-C. La irradiancia de la lámpara en el rango UV-B es de 0.06 W m-2, mientras que la UV-C es de 0.03 W m-2. Los tratamientos aplicados se muestran en el Cuadro 1 (UV-B) y Cuadro 2 (UV-C).

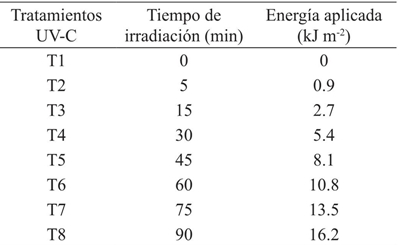
Las dosis de energía en el rango UV-C son menores ya que la lámpara emite radiación con una densidad de flujo fotónico menor. Una vez irradiadas, las semillas fueron colocadas en bolsas de papel aluminizado, para evitar que la radiación del entorno pudiera modificar el estímulo causado por la radiación UV (Musil et al., 1998), y llevadas a siembra al día siguiente.
Las semillas irradiadas se sembraron en rollos de papel anchor para germinación, cada rollo correspondió a una repetición conteniendo 25 semillas, los rollos se distribuyeron en un diseño completamente al azar dentro de una cámara de germinación LAB-LINE a 25 °C con iluminación constante. Cada día se verificó y controló la humedad del papel. Las semillas se mantuvieron bajo estas condiciones por 15 días, después se realizaron las evaluaciones de: porcentaje de semillas germinadas, porcentaje de plántulas normales, longitud de hipocótilo (cm), longitud de radícula (cm) y peso seco (mg), los datos se analizaron en el programa SAS utilizando un Anova y una prueba de comparación de medias mediante Tukey (α≤ 0.05).
La segunda etapa experimental se llevó a cabo de marzo a julio del año 2015 en un invernadero tipo capilla con cubierta rígida de policarbonato ubicado en la Universidad Autónoma Agraria Antonio Narro en Saltillo, Coahuila, México. Los tratamientos aplicados en la segunda etapa se muestran en el Cuadro 3.

Las semillas irradiadas se sembraron directamente en contenedores de poliestireno de 1L conteniendo sustrato de perlita y turba ácida en una proporción 1:1 y distribuidos bajo un diseño completamente al azar, Los contenedores se colocaron en mesas plásticas de 0.85 m de altura para facilitar el manejo y las mediciones. El riego de los contenedores se llevó a cabo inicialmente con agua corriente, a los 10 días ocurrió la emergencia de las plántulas y 10 días después se comenzó a aplicar la fertilización con solución Steiner (Steiner, 1961) a 25%, la cual se aplicó mediante tubines y goteros estaca con gasto de 1.2 L h-1.
Transcurridos 50 días después de la siembra se llevaron a cabo evaluaciones de unidades SPAD en las láminas foliares con un SPAD 502 plus marca Minolta. La altura de las plantas fue medida con una cinta métrica y el diámetro del tallo con un vernier digital a un centímetro de distancia de la base, ambas variables se midieron a los 52 días después de la siembra. A los 65 días después de la siembra se realizaron impresiones epidérmicas sobre la tercera hoja totalmente expandida para determinar la densidad estomática del haz y del envés (estomas mm-2) utilizando un objetivo de 40x de un microscopio compuesto (Carl Zeiss) con cámara digital (Pixera Winder Pro) y un software de medición (Axion Vision Rel. 4.8). Los pesos frescos y secos de las plantas fueron evaluados usando 5 plantas elegidas al azar de cada tratamiento transcurridos 65 días después de la siembra.
El análisis de datos se realizó con el programa SAS versión 9.1 utilizando una ANOVA y una prueba de comparación de medias mediante Tukey (α≤ 0.05).
Resultados y discusión
Respuestas de germinación y vigor frente a la irradiación UV-B
La radiación UV-B provocó diferencias en las variables semillas germinadas (%), plántulas normales (%) y peso seco (g), por otro lado no existieron diferencias en las variables longitud de hipocótilo (cm) y longitud de raíz (cm). En las variables en donde se observaron diferencias entre los tratamientos y el testigo (p≤ 0.05) se observa una tendencia al incremento con un máximo en el tratamiento con 5.4 kJ m-2, después una disminución que alcanza los valores más bajos con 16.2 kJ m-2, posteriormente vuelve a presentar una tendencia positiva alcanzando otro pico en el tratamiento con 27 kJ m-2 (Figura 1).

Figura 1 Diferentes dosis de radiación UV-B aplicada a las semillas de tomate híbrido Big Rio y su efecto en la germinación y vigor de las plántulas. Las barras en cada gráfico señalan valores promedio para cada variable y literales diferentes sobre cada barra indican diferencias significativas según una prueba de Tukey (p≤ 0.05).
En la variable semillas germinadas (Figura 1a), el tratamiento con 5.4 kJ m-2 de radiación UV-B es estadísticamente superior (p≤ 0.05) al testigo, esta clase de respuesta ha sido explicada por Luckey (1980), este científico menciona que la aplicación subletal de un agente capaz de inducir un estrés físico o químico puede generar respuestas positivas en las semillas. Al parecer a los 5.4 kJ m-2 se alcanza un umbral de respuesta en la semilla en el que pudieron actuar ciertos receptores que desencadenaron una respuesta enfocada a acelerar la germinación. Las dosis mayores a 5.4 kJ m-2 dieron lugar a respuestas no diferentes al testigo, pero con el tratamiento de 27 kJ m-2 se alcanza de nuevo un valor 21% superior al testigo. Posiblemente con esta cantidad de energía mayor pudo alcanzarse un segundo umbral de respuesta en la semilla, en dicho umbral pudieron verse involucrados diferentes receptores.
Esta clase de respuestas con varios máximos, y tal vez dependientes de diferentes receptores son muy valiosas para las semillas desde un punto de vista adaptativo, ya que permiten una mayor flexibilidad en la respuesta frente a diferentes contextos ambientales (ejemplos: diferentes profundidades bajo el suelo en un banco de semillas o distintos tipos o coberturas de dosel sobre un banco de semillas) y aseguran que no sea solamente un cierto valor umbral o un estímulo individual el que desencadene el proceso germinativo y el crecimiento del embrión (Long et al., 2015).
Resultados muy similares se encontraron en el porcentaje de plántulas normales (Figura 1b), los tratamientos con 1.8 y 5.4 kJ m-2 fueron superiores al testigo tal vez como consecuencia de un efecto hormético por radiación mencionado por Luckey (1980). Normalmente estas respuestas son positivas ya que la formación de una plántula normal asegura el crecimiento óptimo y mejor productividad (Delibaltova e Ivanova, 2006). El tratamiento con 27 kJ m-2 mostró un nuevo pico de respuesta al igual que en la variable semillas germinadas, lo que pudiera indicar la acción de diferentes receptores o cascadas de señalización activadas con diferentes umbrales de energía.
En la variable longitud de hipocótilo (Figura 1c) al igual que en la longitud de raíz (Figura 1) no existió diferencia significativa (p≥ 0.05). Otros autores encontraron respuesta a la irradiación en estas variables, Kacharava et al. (2009) irradiaron semillas de frijol durante 30 minutos y encontró un incremento en la altura de planta, pero con 90 min la altura disminuyó. De igual manera, Hidema et al. (2006) encontraron que la radiación suplementaria con UV-B disminuyó el crecimiento de la planta y la produción de grano en el arroz. No se dispone hasta el momento de una explicación de porqué frente al estímulo con UV-B ocurren respuestas diferentes, positivas o negativas o neutras, entre especies incluso de la misma familia (Kossuth y Biggs, 1981). Lo mismo ocurre con las distintas respuestas exhibidas por diferentes variables en la misma especie, tal como se muestra en este estudio.
En el peso seco (Figura 1e) se repite la misma respuesta ya descrita para el porcentaje de semilas germinadas y de plántulas normales, con máximos en 5.4 y 27 kJ m-2. Un resultado análogo fue descrito por Kacharava et al. (2009), quienes reportaron un incremento en la materia seca aplicando 60 minutos de radiación ultravioleta a semilla de frijol.
Respuestas de germinación y vigor frente a la irradiación UV-C
Al contrario que con la irradiación con UV-B en el caso de la UV-C no se presentaron respuestas positivas. Posiblemente que esta diferencia provenga del hecho de que, al contrario que la UV-B, la radiación UV-C no es un componente natural en los ecosistemas (Nawkar et al., 2013). En concordancia con Promyou y Supapvanich, (2012) es posible que las respuestas observadas en las semillas germinadas y las plántulas sean resultado del daño oxidativo causado por la exposición inclusive a las dosis más pequeñas de UV-C. Ello explica en parte por qué las respuestas negativas fueron evidentes en los procesos más sensibles al daño oxidativo sobre la germinación y la obtención de plántulas normales después de la germinación (Figura 2a y 2b). Mientras que los procesos posteriores que dieron lugar a la longitud de hipocótilo, la longitud de la raíz y el peso seco, que son variables evidenciadas solamente en plántulas normales, no se vieron afectadas (Figuras 2c, 2d y 2e). Estos últimos resultados son coincidentes con los reportados por Foroughbakhch-Pournavab et al. (2015) en trigo y girasol.
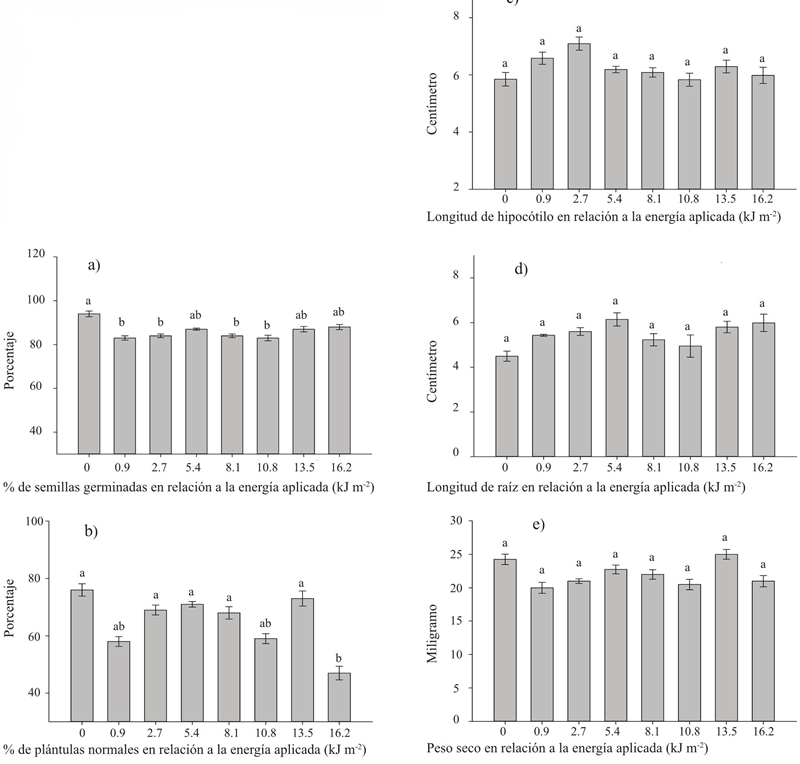
Figura 2 Diferentes dosis de radiación UV-C aplicada a las semillas de tomate híbrido Big Rio y su efecto en la germinación y vigor de las plántulas. Las barras en cada gráfico señalan valores promedio para cada variable y literales diferentes sobre cada barra indican diferencias significativas según una prueba de Tukey (p≤ 0.05).
En lo referente a semillas germinadas (Figura a2) la respuesta frente a la radiación fue prácticamente la misma con todas las dosis de UV-C, mostrando valores menores al testigo con las dosis de 0.9, 2.7, 8.1 y 10.8 kJ m-2, mientras que con las dosis de 5.4, 13.5 y 16.2 no existieron diferencias significativas (p≥ 0,05), estos resultados señalan la gran sensibilidad de la respuesta germinativa frente al estímulo con UV-C y son muy similares a los reportados por Brown et al. (2001), quienes irradiaron semillas de lechuga y encontraron respuestas de este tipo, en función de la dosis, en las variables de incidencia de Xanthomonas campestris pv. campestris y en la de unidades formadoras de colonias.
En el porcentaje de plántulas normales (Figura 2b) solo la dosis más alta de 16.2 kJ m-2 mostró un valor más bajo que el testigo, lo que de nuevo apoya la explicación de que al contrario que el estímulo con UV-B, que parece una respuesta a un estímulo fotomorfogénico, la respuesta frente a la UV-C se da por el daño oxidativo o de otra índole que causa la gran cantidad de energía acarreada por los fotones UV-C. Estos resultados coinciden con los de Brown et al. (2001), quien reporta que el diámetro de la cabeza de plantas de lechuga, obtenidas de semillas tratadas con dosis bajas de radiación UV-C no fue diferente al testigo, mientras que en las provenientes de semillas tratadas con dosis más altas el diámetro se redujo considerablemente. Otras respuestas negativas frente a la radiación UV-C aplicada en tejidos de plantas son el bajo contenido de proteína, disfunciones en los cloroplastos, daños al ADN y en varios procesos celulares (Danon y Gallois, 1998; Rastogi et al., 2010; Ruiz-López et al., 2010).
Respuestas de las plantas obtenidas de las semillas irradiadas con UV-B y UV-C
En la segunda etapa, las variables de unidades SPAD y densidad estomática del haz y envés fueron diferentes al testigo, lo que indica que la radiación aplicada a la semilla es capaz de generar cambios que se hacen visibles en etapas posteriores (Figura 3), tal como fue descrito por Musil et al. (1998) y Magliano y Casal (2004) para las señales ambientales percibidas en la pre-germinación por los fitocromos.

Figura 3 Respuesta de las plantas obtenidas de semillas de tomate híbrido Big Rio irradiadas con UV-B y UV-C y sembradas en macetas bajo invernadero. Las barras en cada gráfico señalan valores promedio para cada variable y literales diferentes sobre cada barra indican diferencias significativas según una prueba de Tukey (p≤ 0.05).
Para la variables altura de planta (Figura 3a) y diámetro de tallo (Figura 3b) ningún tratamiento fue diferente al testigo. Este comportamiento de las plantas de tomate fue diferente al reportado para otras especies como el repollo que mostró mayor crecimiento al irradiarse las semillas con 3.6 kJ m-2 de UV-C (Brown et al., 2001), Arabidopsis con disminución en los pecíolos frente a la radiación UV-B (Gruber et al., 2009) o la soya que mostró menor longitud en las plántulas cuando sus semillas fueron irradiadas con UV-C (Foroughbakhch-Pournavab et al., 2015). De nuevo parece aplicar la diferente respuesta de cada especie (Kossuth y Biggs, 1981) considerando factores como el tamaño de la semilla, el grosor de testa, la composición química u otras propiedades intrínsecas de la semilla de cada especie.
En la densidad estomática del haz (Figura 3c) y del envés (Figura 3d) se encontraron cambios inducidos por la irradiación de las semillas con UV-B y UV-C determinando que la respuesta en las plantas fue elevar la densidad estomática en el haz, mientras que para la densidad estomática en el envés se encontró una tendencia inversa frente a la mayor dosis de energía aplicada con UV-B, pero una respuesta contraria al aplicar la UV-C. Wargent et al. (2009) indicaron que este tipo de respuestas adaptativas frente a la radiación UV impactan en el desempeño de las plantas, modificando la interacción con los diferentes factores ambientales a través de cambios en la forma, fisiología y bioquímica.
Resultados análogos fueron obtenidos por Benavides-Mendoza et al. (2003) al irradiar semillas de trigo con un láser de baja intensidad (AsAlGa), observando que con una dosis de 6 kJ m-2 las plántulas obtenidas de las semillas irradiadas mostraron valores mayores de densidad estomática en el haz y en el envés al crecer en un ambiente propicio para el crecimiento, mientras que al germinar en un medio con diferentes niveles de salinidad las plántulas irradiadas presentaron una clara respuesta adaptativa disminuyendo la densidad estomática, respuesta que no fuen observada en el testigo. Es probable que las plantas de tomate obtenidas de las semillas irradiadas con UV muestren respuestas con algún trasfondo de adaptación a un ambiente en donde se detecta una mayor cantidad de energía radiante, lo cual implicaría la necesidad de ajustes en los caracteres de la epidermis.
En las variables de peso fresco (Figura 3e) y peso seco (Figura 3f) no se encontró diferencia de ninguno de los tratamientos con el testigo. De nuevo estos resultados parecen indicar una respuesta específica para las plantas de tomate, ya que Kacharava et al. (2009) observaron incrementos en el peso fresco de algunas variedades de Phaseolus vulgaris con aplicaciones de 60 y 90 min de UV-B; así como, un aumento en el peso de remolacha blanca con 60 min de UV-B aplicada a la semilla. En la lechuga la irradiación de las semillas con un láser de baja intensidad causó aumento en la biomasa radical pero no cambió el peso de las hojas y el tallo (Benavides-Mendoza et al., 2003).
Las unidades SPAD (Figura 3g) disminuyeron con los tratamientos 5.8 y 10.5 kJ m-2 de UV-B. La respuesta puede explicarse considerando que las unidades SPAD miden indirectamente la concentración de clorofila en las láminas foliares (Rodríguez-Mendoza et al., 1998) y que este pigmento responde de forma inversa a la cantidad de energía radiante detectada en el ambiente (Vu et al., 1984; Strid et al., 1990). Los resultados aquí obtenidos indican claramente que dicha señalización ocurre desde la semilla. En contraste con la UV-B en el caso de la irradiación con UV-C no hubo diferencias con el testigo, pero la Figura 3g muestra una tendencia positiva en los datos distinta a la exhibida por los de UV-B.
Conclusión
La irradiación de las semillas de tomate con UV-B y UV-C modificó tanto las respuestas en germinación y vigor como el crecimiento de las plantas obtenidas de las semillas irradiadas. La irradiación con UV-B en 5.4 kJ m-2 aumentó la germinación y la cantidad de plántulas normales obtenidas.
En cambio, la irradiación de las semillas de tomate con UV-C no presentó ningún efecto positivo en la germinación y el vigor.
Los efectos de la irradiación de las semillas con UV-B y UV-C se extendieron a las etapas posteriores de crecimiento de las plantas. La irradiación con UV-B con 10.8 disminuyó las lecturas de unidades SPAD en las hojas y disminuyó la densidad estomatica en el envés. La densidad estomática del haz fue aumentada en todas las plantas de semillas irradiadas con UV-B, independientemente de la dosis. Por su parte las plantas obtenidas de semillas irradiadas con UV-C mostraron aumento en la densidad estomática en el haz y disminución en la del envés.

