










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.7 spe 14 Texcoco feb./mar. 2016
Articles
Tree structure of cocoa agroforestry system in Cárdenas, Tabasco, Mexico
1Universidad Autónoma de Chiapas. Escuela Maya de Estudios Agropecuarios, Playas de Catazaja. (sanchez.facundo@colpos.mx).
2Colegio de Postgraduados-Campus Tabasco. Periférico Carlos A. Molina S/N, km 3.5 A. P. 24 Cárdenas, Tabasco, México. C. P. 86500. (obradoro@colpos.mx; sol@colpos.mx).
3Colegio de Postgraduados-Campus Veracruz. Carretera Federal Veracruz-Xalapa-Vía Paso de Ovejas, km 26.5. A. P. 421, Veracruz, Veracruz, México. C. P. 91700. (octavior@colpos.mx).
In this paper the floristic composition and structure and tree species diversity between cacao agroforestry systems (SAF-cacao) of different age were compared. In 20 sampling sites of 5 000 m2 each, from each tree measured diameter at breast height (DAP1.3 m) and total height (Ht); basal area (AB, m2 ha-1), importance value index (IVI), diversity indices (H') and Shannon index (E) and similarity indices were calculated. 2 856 trees were recorded, belonging to 67 species from 58 genera and 28 botanical families. The most diverse families were Fabaceae, Moraceae, Rutaceae, Sapotaceae and Verbenaceae. The 25 and 20 years SAF had the highest and lowest basal area (AB) with 22.7 and 12.16 m2 ha-1, respectively. As for IVI, Erythrina americana, Cedrela odorata, Poeppigiana erythrina, Gliricidia sepium and Colubrina arborescens, represented 53.1% of the 67 species recorded. The 35 and 6 years SAF had the highest and lowest diversity with H' values of 2.89 and 0.92, respectively. The older the SAF, were more uniform. The 18 and 38 years and between 25 and 30 years SAF-Cocoa were the most similar in tree species composition, respectively.
Keywords: cacao agroforestry system; structure and tree diversity
En el presente trabajo se comparó la composición florística, y la estructura y diversidad de especies arbóreas entre sistemas agroforestales de cacao (SAF-cacao) de diferente edad. En 20 sitios de muestreo de 5 000 m2 cada uno, de cada árbol se midió el diámetro a la altura del pecho (DAP1.3 m) y la altura total (Ht); y se calculó el área basal (AB, m2 ha-1), índice de valor de importancia (IVI), los índices de diversidad (H’) y uniformidad de Shannon (E) y los índices de similitud. Se registraron 2 856 árboles, pertenecientes a 67 especies de 58 géneros y 28 familias botánicas. Las familias más diversas fueron Fabaceae, Moraceae, Rutaceae, Sapotaceae y Verbenaceae. Los SAF de 25 y 20 años presentaron la mayor y menor área basal (AB) con 22.7 y 12.16 m2 ha-1, respectivamente. En cuanto al IVI, Erythrina americana, Cedrela odorata, Erythrina poeppigiana, Gliricidia sepium y Colubrina arborescens, representaron 53.1% de las 67 especies registradas. Los SAF de 35 y 6 años presentaron la mayor y menor diversidad con valores H’ de 2.89 y 0.92, respectivamente. A mayor edad de los SAF, estos fueron más uniformes. El SAF-cacao de 18 y 38 años y los de 25 y 30 años de edad fueron los más similares en composición de especies arbóreas, respectivamente.
Palabras clave: estructura y diversidad arbórea; sistema agroforestal cacao
Introduction
Globally 70% of SAF-cocoa is grown in association with shade trees and annual and perennial crops (Salgado et al., 2007). Shade trees constitute an element which may contribute to the sustainability of this system due for the production of dead leaves, nutrient recycling and prevention of soil erosion (Alvim and Nair, 1986; Salgado et al., 2007). Besides SAF-cocoa can be a space for biodiversity conservation, carbon capture and cushion for adverse weather conditions, aspect that is worthless even for most of the cocoa growing areas of Mexico and the world (Parrish et al., 1999; Roa et al., 2009).
In Mexico most of the cocoa production is located in Chiapas and Tabasco. These states have a combined area of 61 444 ha, of which 47 000 farmers depend economically. The planted area in Tabasco is 41 117 ha, from which are obtained 16 560 t of dry cocoa, sustaining 31 139 families (SAGARPA, 2010; FAOSTAT, 2011); 96% of the cocoa area is distributed in the Chontalpa region and 4% in the Sierra region (López et al., 2005). Therefore, the Chontalpa region is the main cocoa producing area in Mexico and where most studies on the cultivation of cocoa have developed.
The studies on shade tree diversity in SAF-cocoa in Mexico are scarce but have determined that the most common species are Erythrina americana Mill, Diphysa robinoides Benth, Gliricidia sepium (Jacq.) Walp, Samanea saman (Jacq.) Merr and Colubrina arborescens (Mill.) Sarg, fruit trees Mangifera indica, Citrus sp. and Pouteria sapota (Jacq.) H.E. and forest timber Cedrela odorata L. (Ramos 2001; Córdova et al., 2001). In Nigeria, cocoa is associated with Cola nitida (Sterculiaceae), plant considered as industrial species (Salgado et al., 2007).
Most studies of FAS-cocoa focus on agronomic description of production systems and some in wildlife (Parrish et al., 1999). Ibarra and Estrada (2001) and Salgado et al. (2007) cite the cacao maintain a wide diversity of birds, bats, non-flying mammals and invertebrates, similar to the natural forest and superior to other agricultural habitats of intensive use. Mammal species like the mantled howler (Alouatta palliata) have persisted in cacao farms in the municipality of Comalcalco, Tabasco, where they feed on tree species such as Ficus cotinifolia, F. obtusifolia, Ficus sp, Gliricidia sepium and Samanea saman; this mammal is reported as endangered species according to NOM-059 of 2002 (Muñoz et al., 2005). Pérez (2008) indicates that cacao germplasm diversity affects the abundance and diversity of beetles from the family Scolytidae; this author collected and identified 19 species of host plants belonging to 11 botanical families, being cocoa (Theobroma cacao) and mahogany (Swietenia macrophylla) the tree species where more organisms were collected.
Ramírez et al. (2013) analyzing the floristic diversity of 30 and 50 years old SAF-cacao in Tabasco, Mexico reported that older systems are more diverse than younger. This diversity is higher than in other intensive systems and similar to species diversity of evergreen tropical forest from Tabasco (Zarco et al., 2010). Therefore the objective of this study was to compare the floristic composition, structure and diversity of tree species in the SAF-cocoa with different age Cárdenas, Tabasco.
Materials and methods
Study area
The study was conducted in SAF-cocoa from different localities of the municipality of Cardenas: village C-11, C-28. Cárdenas-Coatzacoalcos road, km 21, Santa Ana 2nd section and Miguel Hidalgo 2nd section. The municipality of Cárdenas is located in the Chontalpa region in Tabasco, main producing area of cocoa in Mexico. The extreme coordinates of the municipality are 17° 59 north latitude and 91° 32' west longitude and is located at an altitude of 2-17 masl; it has a warm-humid climate, with annual and monthly rainfall of 2 643 and 355 mm, respectively, and an average annual temperature of 26 °C and maximum 45 °C.
Study plots and sampling procedure
The study plots were selected from field trips and based on geographic information of cocoa fields made by Secretaria de Desarrollo Agropecuario Forestal y Pesca (SEDAFOP) and the list of cocoa producers 2008 (SEDAFOP and OEIDRUS, 2008; OEIDRUS, 2007). Also, the authorities from the ejido and municipal delegates of each locality were interviewed, in order to contact cooperating producers (owners of the land) and seek their support in the investigation.
20 sampling sites of 100 x 50 m (5 000 m2) each were established. At each site the floristic composition was determined by taxonomic identification of tree species. The trees were geographically located and listed. At each site, the age and surface was recorded through an interview addressed to the cooperating producer (Zarco et al., 2010). The recorded forest variables were: diameter at breast height (DAP1.3 m) and total height (Ht) with tape measures and an inclinometer Haga type, respectively (Scheelje, 2009). The SAF were grouped according to age and the structural parameters, diversity indices and floristic composition were measured from species.
Structural analysis
The structural index was evaluated by the importance value index (IVI) (Zarco et al., 2010). IVI was estimated by the equation: IVI= DR + ABR + FR.
Where: DR is relative density; ABR relative basal area and FR is relative frequency (Scheelje, 2009). DR, AB, ABR and FR were calculated with the following equations:
Diversity and uniformity analysis
For the diversity analysis the diversity index of ShannonWiener (H’) were used. This index is based on the proportional abundance of species (Magurran, 1989). The equation used was H’= -Σpi(lnpi) where: Pi is the ratio of total sample found for species i and ln is the natural logarithm.
The equity index Shannon-Wiener (E) was calculated to determine the relative abundance at species level represented as equity or uniformity. This index was used to determine the degree to which individuals of different species are represented equally at SAF level and was estimated with the equation:
Where: H’= diversity index of Shannon-Wiener, lnS is the natural logarithm of the number of species.
Similarity indices
The similarity analysis between the reported ages of SAF-cocoa was determined by Jaccard and Sorenson method for qualitative data and the Sorenson and Morisita Horn method for quantitative data. In the four methods, the values tend to be equal to one in case of complete similarity and zero if populations are different and have no species in common (Mostacedo and Fredericksen, 2000). The equations used to calculate these indices are shown in Table 1.
Table 1 Similarity indices used to compare cocoa agroforestry systems of different ages in Cárdenas, Tabasco, Mexico.

A= número de especies presentes en el sitio a; b= número de especies presentes en el sitio b; c= número de especies presentes en ambos sitios; aN= número de individuos en el sitio A; bN= número de individuos en el sitio B; jN= suma de las abundancias de especies en el sitio que presenta la abundancia inferior; ani= número de individuos de la i-esima especie en el sitio A; bni= número de individuos de la i-esima especie en el sitio B.
To calculate the diversity, equity and similarity indeces the InfoStat software version 0.1 and BIO-DAP (Di Rienzo et al., 2008) were used. For diversity and equity with the age of SAF-cocoa, a simple linear correlation analysis was performed to determine the trend number and uniformity of species as the age of SAF-cocoa advances.
Results and discussion
Floristic composition
Tree species in the SAF-cocoa play an important role for biodiversity conservation and germplasm, availability in Tabasco, Mexico. 10 ha were sampled from 36.5 ha of SAF-cacao visited. The average size of the SAF was 1.8 ha, varying from 0.5 to 5 ha, indicating that cacao plantations are in the hands of small farmers. The SAF visited were grouped into nine ages ranging from 6 to 35 years (Table 3). 2 856 trees belonging to 67 species and 58 genera, grouped in 28 families were recorded.
The five species with the highest number of individuals from the total sampled sites were Erythrina americana Mill. (812 individuals), Cedrela odorata L. (573) Gliricidia sepium (Jacq.) Walp. (247) Colubrina arborescens (Mill.) Sarg. (246) and Diphysa robinioides Benth. (188); representing 72.4% of total trees registered. The 10 most diverse families were Fabaceae with 10 species, Moraceae 4, Rutaceae 4, Sapotaceae 4, Verbenaceae 4, Arecaceae 3, Meliaceae 3, Rubiaceae 3 and Sterculiaceae 3 species. Families Fabaceae, Meliaceae, Rhamnaceae and Bignoniaceae were present at the SAF-cocoa age of 9 and in them highlighted by frequency E. americana, C. odorata, C. arborescens and Tabebuia rosea, respectively.
Ramos (2001) in 9 ha of SAF-cacao in Tabasco, recorded 33 species belonging to 31 genera and 17 families in our study the species D. robinoides and G. sepium species were the most frequent. In the same locality Ramirez et al. (2013) in 6 ha of SAF-cocoa recorded 1,741 trees, from 49 species, 45 genera and 25 families and as in this investigation, reported that the species D. robinoides and C. arborescens were the most frequent. In the Soconusco region, Chiapas Salgado et al., (2007) in 7.2 ha recorded 720 trees, 47 species, 38 genera and 23 families; the species G. sepium was the most frequent in the SAF-cocoa. This study and the three studies cited agreed on the Fabaceae family as the most common for shade in the SAF-cocoa in Mexico. In contrast, Guiracocha et al. (2001) in Costa Rica, Rosa (2003) in Brazil, Orozco and Somarriba (2005) in Bolivia and Hervé and Vidal (2008) in Cameroon reported the Moraceae and Anacardiaceae families as the most frequent in the SAF-cocoa; the authors attributed the above to native tree species have not been completely eliminated from the forests where SAF-cocoa were established, and in these forests abound the genera Artocarpus, Ficus and Brosimum of the cited families.
Vertical and horizontal stratification
The average height of the tree component in the SAF-cocoa was 10.1 m, varying from 2 to 35.5 m. 90.1% of registered trees measured less than 15 m. The average DAP was 23 cm, varying from 1 to 146.6 cm. The total average density was 286 trees ha-1, varying from 96 to 618 trees ha-1. The SAF-cocoa of 6 and 33 years had the highest and lowest density, respectively (Table 2). The 25 years SAF recorded greater AB, with a mean value of 22.7 m2 ha-1 predominating Erythrina poeppogiana and Samanea saman; the lowest AB were for the 20 years old SAF-cacao with 12.16 m2 ha-1 (Table 2) made up of small trees. Particularly in the 20 years SAF-cacao the producer changed mote (E. americana) and chipilcohite (D. robinoides) established as shade trees for cedar (C. odorata) as timber species; however, this SAF ranked second in density.
Table 2 Average values of forest variables, families and plant species estimated in cacao agroforestry system in Cárdenas, Tabasco, Mexico.
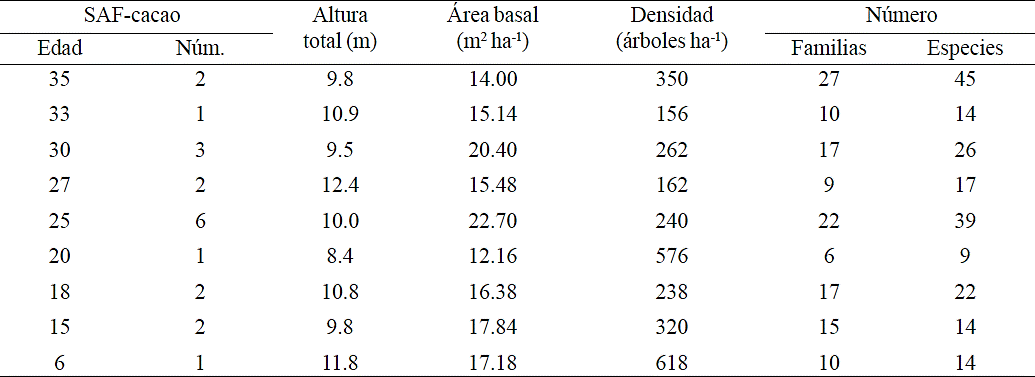
In the SAF-cocoa ≤20 years, recorded higher tree density and in SAF ≥25 years, recorded higher diversity. This can be explained because as the trees develop, the producer cut some trees (thinning the system) and plants other (diversifying the system). The trees planted are fruits, timber and ornamental species. This diversification also contributes to the distribution of seeds that birds and rodents do. In the 35 and 25 years SAF were found 96 and 79% of the 28 families and 67 and 58% of the 67 species recorded; The 20 years are less diverse with 13% of species and 21% of families registered in all SAF-cocoa sampled (Table 2).
Structural indices
The five species with the highest importance value index (IVI) present in the 20 sites sampled were: E. americana, C. odorata, E. poeppigiana, G. sepium and C. arborescens representing 53% of the recorded species. In relative density (DR) and relative dominance (ABR) E. americana had the highest values and C. odorata had the highest relative frequency (FR). Erythrina poeppigiana had the lowest DR and FR among the five species cited but ranked second and third place in ABR and IVI, respectively (Table 3) due to its greater AB.
Table 3 Density, dominance, frequency and importance value index of tree species in cacao agroforestry systems from nine ages in Cárdenas, Tabasco, Mexico.
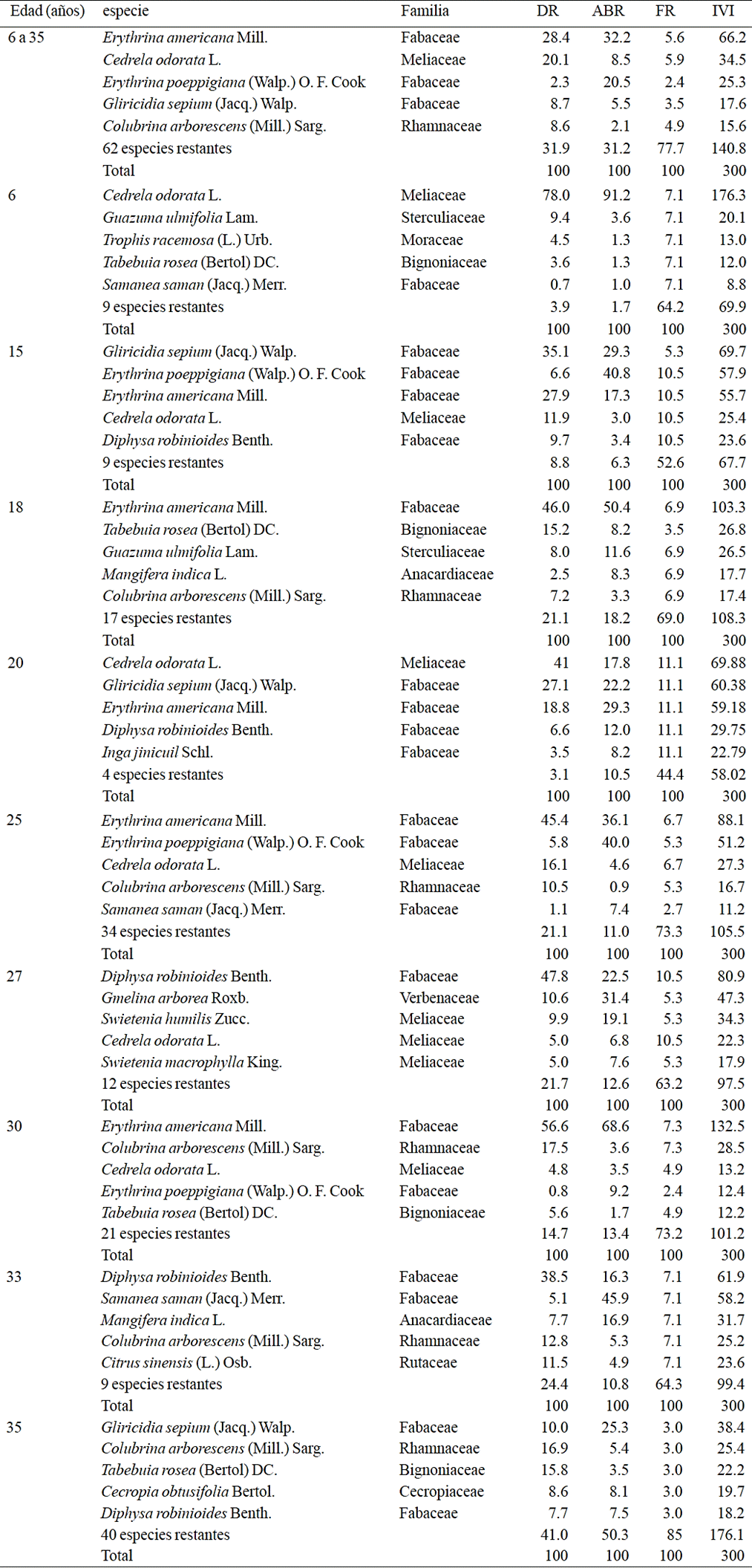
DR= densidad relativa (%); ABR (%)= dominancia relativa (%); FR= frecuencia relativa (%), IVI= índice de valor de importancia.
The younger SAF-cocoa had a higher tree density per hectare. At structural level the 25 years SAF-cocoa had higher AB averaging 22.7 m2 ha-1, mainly due to a higher tree density of E. poeppigiana and S. Saman which are species of larger diameter. Although in density ranked second the 20 years old SAF-Cocoa with lower AB averaging 12.16 m2 ha-1. The latter is due to the introduction of timber species C. odorata and C. arborescens by CONAFOR. The 30 years SAF-Cocoa had an average AB of 20.4 m2 ha-1. Ramirez et al. (2013), in 50 and 30 years old plantations reported a AB of 58 and 38 m2 ha-1, respectively; the AB difference of the 30 years SAF was primarily attributed to the recorded species among which S. saman excelled.
Based on IVI and the age of the SAF-cocoa, the outstanding species were C. odorata in the 6 and 20 years old SAF, G. sepium in 15 and 35 years, E. americana in 18, 25 and 30 years and D. robinioides in 27 and 33 years (Table 3); the variation of tree species used as shade in the SAF-cocoa, is due to objective and preference of each producer, where Fabaceae and Meliaceae are the most common.
The structurally important species according to IVI, E. americana, C. odorata, E. poeppigiana, G. sepium and C. arborescens, agreed with that reported by Ramírez et al. (2013) in 30 and 50 years old plantations in Cárdenas, Tabasco. Leon (2006) in Ecuador reported species with higher IVI to Mangifera indica, Tabebuia rosea, Artocarpus altilis, Quararibea cordata and Inga spectabilis. Orozco and Somarriba (2005) in Bolivia reported with the highest IVI, Schyzolobium parahyba, Swietenia macrophylla, Amburana cearensis, Centrolobium ochroxylum and C. odorata. Rosa (2003) in Brazil reported species with the highest IVI, Schefflera morototoni, Trema micrantha, Helicostylis tomentosa, Sloanea obtusifolia and Macrolobium latifolium; these species differ from those in this study because cocoa farms in Brazil are established under undisturbed forests, unlike plantations in Chontalpa, Tabasco that are established under tree species introduced by the producer. However, all cases confirm the importance of SAF-cocoa as centers of biodiversity conservation, by the diversity of tree species that possess and habitat function for flora and fauna.
Diversity (H’) and equity (E)
According to the diversity index of Shannon the older the SAF-cocoa there is greater diversity at species level. The SAF-cocoa with age of 35 years old were those that recorded greater diversity of tree species (H’= 2.89), followed by 25 and 33 years which were equal in diversity (H’= 2.07), and the 6 years old SAF had lower diversity (H’= 0.92). In equity value of Shannon (E) the 33 years old SAF-cocoa recorded higher equity in number of individuals per species, the 15, 20 and 27 years old showed the same equity value (E = 0.67) and the 6 years SAF recorded the lowest value (E = 0.35) (Table 4).
Table 4 Diversity and equity of species in cacao agroforestry systems from nine ages in Cárdenas, Tabasco, Mexico
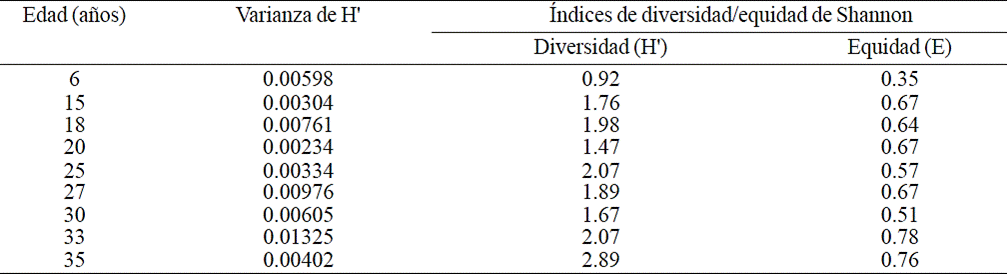
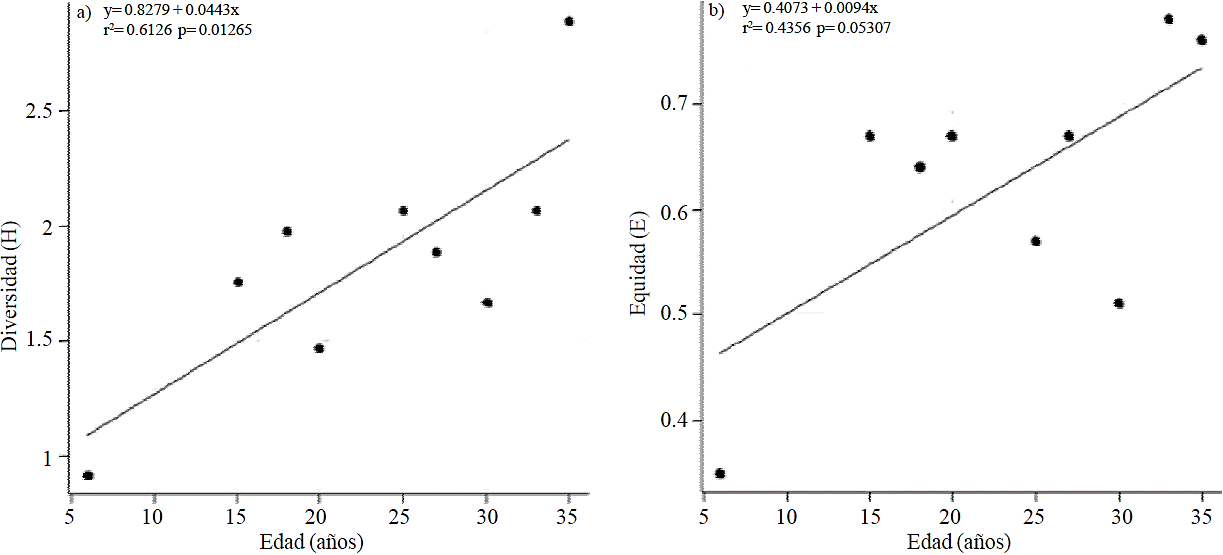
The correlation between age and diversity (r2= 0.6126, p= 0.01), also indicated that as SAF grows older, are more diverse. This agreed with that reported by Ramírez et al. (2013) in Cardenas, Tabasco for 50 and 30 years old SAFcocoa (H’= 2.89 and 2.57, respectively). In contrast, Ramos (2001) in cocoa plantations in Tabasco, found that the older, less diversity (H’= 1.24, 1.75 and 1.46, for 15, 20 and 30 years, respectively), probably due to thinning of the tree species during plant development.
In the Soconusco region in the state of Chiapas Salgado et al. (2007) reported diversity values (H’= 2.74 to 2.79) coinciding with those found in our study. However, these values and ours are lower than those reported by Rosa (2003) in Brazil (H’= 3.31 to 4.22) because in this country the cultivation of cocoa is established in disturbed forests, where there is greater plant diversity. Guiracocha et al. (2001) in Talamanca, Costa Rica reported diversity values (H’= 2.56, 1.75 and 1.10, for forest, SAF-cacao and banana plantations, respectively), indicating a greater degree of species conservation in forest. The present study recorded greater diversity of species, than the three studied systems. Bojorges and López (2006) and Zarco et al. (2010) in evergreen tropical forest of Veracruz and Tabasco, respectively, found values of H’= 2.5. Corral et al. (2002) in the cloud forest in Tamaulipas reported H’= 2.39; both results are lower than those of 35 years SAF-cocoa (H = 2.89) in this study, indicating that the SAF-cocoa Mexico could have more tree diversity than forest and jungle.
The correlation between age of the SAF-cocoa and diversity (H’), indicated a good positive relationship (r2= 0.6126) (Figure 2) statistically significant (p= 0.01), indicating that the older the SAF, these have greater diversity of species and families. This diversity helps the producer to plant other trees (fruit, ornamental, and timber) and wildlife by seed dispersal. The correlation of age with equity indicated that there is a moderate positive relationship (r2= 0.43569) (Figure 1), statically not significant (p= 0.05); indicating that the older the SAF-cocoa, these are more uniform in number of individuals per species.
Similarity indices
The values from Sörenson and Jaccard indices for 18 and 33 years old SAF-cocoa, indicated that these tend to be slightly similar in species composition (0.67 and 0.50), while the 6 and 15 years old SAF tend to greater dissimilarity (0.21 and 0.24); this indicates that elderly SAF-cocoa tend to similarity in number of species, compared to younger. Likewise, the values from Morisita -Horn and Sorenson indices for 25 and 30 years old SAF-cocoa showed that these tend to be similar in number of trees of the same species (0.94 and 0.67), while 6 year old with 30 and 33 years old SAF-cocoa tend to greater dissimilarity (0.08 and 0.10, 0.11 and 0.07, respectively); this indicates that older SAF-cocoa tend to be similar in number of individuals per species.
The values from the similarity indices of both qualitative and quantitative showed a clear trend of similarity in number of species and individuals among older plantations (25-30 years). These results agree with those reported by Ramírez et al. (2013) in 30 and 50 years old plantation. Both studies contrast with the results from Ramos (2001) who reported that the older the SAF-cocoa there is greater dissimilarity at species level. Hervé and Vidal (2008) in Cameroon indicated that the type of management is a factor that determines a smaller percentage of similarity, inherent situation to every cocoa region of the world. In the present study, the SAF-cocoa neighbored with home orchards, reason why older SAF-cocoa tends to be more similar to each other at the species level. The latter is because it is a common practice for cocoa producer to introduce new tree species in the SAF, both native and exotic species as time passes.
Conclusions
In the 20 sites sampled 2856 trees belonging to 28 families, 58 genera and 67 species were recorded. The species E. americana, E. poeppigiana, G. sepium (Fabaceae), C. odorata (Meliaceae) and C. arborescens (Rhamnaceae), were the most common species for shade in the SAF-cocoa. By frequency, density and basal area, these five species showed the highest structural index at different ages of the SAF sampled. The older SAF-cocoa tended to be more diverse in species and botanical families than the youngest. For qualitative similarity indices the 18 and 33 years old SAF-cocoa were the most similar in number of trees and species; the highest quantitative similarity was recorded between the 25 and 30 years old SAF. By diversity of tree species recorded in the SAF-cocoa, it can be proposed as a center of biodiversity conservation, or as a natural area for wildlife refuge.
Literatura citada
Alvim, R. and Nair, P. K. R. 1986. Combination of cocoa with other plantation crops. Agroforestry Systems. 4:3-15. [ Links ]
Bojorges, B. J. C. y López, M. L. 2006. Asociación de la riqueza y diversidad de especies de aves y estructura de la vegetación en una selva mediana subperennifolia en el centro de Veracruz, México. Revista Mexicana de Biodiversidad. 77:235-249. [ Links ]
Córdova, A. V.; Sánchez, H. M.; Estrella, C. N. G.; Macías, L. A.; Sandoval, C. E.; Martínez, S. T. y Ortiz, G. C. F. 2001. Factores que afectan la producción de cacao (Theobroma cacao L.) en el ejido Francisco I Madero del Plan Chontalpa, Tabasco, México. Universidad y Ciencia. 17(34):93-100. [ Links ]
Corral, R. J.; Aguirre, C. O. A.; Jiménez, P. J. y Návar, C. J. J. 2002. Muestreo de diversidad y observaciones ecológicas del estrato arbóreo del bosque mesófilo de montaña “El Cielo”, Tamaulipas. Revista Chapingo. Serie Ciencias Forestales y del Ambiente. 8:125-131. [ Links ]
Di Rienzo, J. A.; Casanoves, F.; Balzarini, M. G.; González, L.; Tablada, M. y Robledo C.W. 2008. InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
FAOSTAT. 2011. Sistema Estadístico de la Organización de las Naciones Unidas para la Agricultura y la Alimentación. Consultado: Consultado: http://faostat.fao.org/site/567/default.aspx . 20/01/2012. [ Links ]
Guiracocha, G.; Harvey, C. A.; Somarriba, E.; Krauss, U. y Carrillo, E. 2001. Conservación de la biodiversidad en sistemas agroforestales con cacao y banano en Talamanca, Costa Rica. Agroforestería en las Américas. 8(30):7-11. [ Links ]
Hervé, B. D. y Vidal, S. 2008. Plant biodiversity and vegetation structure in traditional cocoa forest gardens in southern Cameroon under different management. Biodiversity and Conservation. 17(8):1821-1835. [ Links ]
Ibarra, M. A. y Estrada, M. 2001. Avifauna asociada a dos cacaotales tradicionales en la región de la Chontalpa, Tabasco, México. Universidad y Ciencia. 17:101-112. [ Links ]
López, A. P. A.; Delgado, N. V. H.; Azpeitia, M. A.; López, A. J. I.; Jiménez, C. J. A.; Flores, R. A.; Fraire, S. L. y Castañeda, C.R. 2005. El cacao en Tabasco: manejo y producción. INIFAP; ISPROTAB. Tercera edición. Instituto para el Desarrollo de Sistemas de Producción del Trópico Húmedo de Tabasco. Villahermosa, Tabasco, México. 54 p. [ Links ]
León, M. R. A. 2006. Diversidad vegetal asociada a cacaotales de dos zonas agroecológicas en la región litoral del ecuador. Tesis, Facultad de Ingeniería Mecánica, Escuela Superior Politécnica del Litoral, Guayaquil, Ecuador. 114 p. [ Links ]
Magurran, E. A. 1989. Diversidad ecológica y su medición. (A. M. Cirer, Trad.) Barcelona, España: Vendra. 200 p. [ Links ]
Mostacedo, B. y Fredericksen, T.S. 2000. Manual de métodos básicos de muestreo y análisis en ecología vegetal. Proyecto de Manejo Forestal Sostenible (BOLFOR). Santa Cruz, Bolivia. 92 p. [ Links ]
Muñoz, D.; Estrada, A. y Naranjo, E. 2005. Monos aulladores (Alouatta palliata) en una plantación de cacao (Theobroma cacao) en Tabasco, México. Universidad y Ciencia. 2:35-44. [ Links ]
OEIDRUS. 2007. Mosaico estatal de cultivo Geo-Referenciado de cacao. SAGARPA; SEDAFOP. 1 p. Consultado: Consultado: http://www.campotabasco.gob.mx/ . 16/01/12. [ Links ]
Orozco, L. y Somarriba, E. 2005. Árboles maderables en fincas de cacao orgánico del Alto Beni, Bolivia. Agroforestería en las Américas. 43:43-44. [ Links ]
Parrish, J.; Reitsma, R.; Greenberg, R.; Mclarney, W.; Mack R. y Lynch, J. 1999. Los cacaotales como herramienta para la conservación de la biodiversidad en corredores biológicos y zonas de amortiguamiento. Agroforestería en las Américas. 6(22):16-19. [ Links ]
Pérez, C. M. 2008. Diversidad y abundancia de escolitidos (Coleoptera: Scolytidae) asociados al agroecosistema cacao en Tabasco, México. Tesis Doctoral Colegio de Postgraduados Montecillo. Edo. México. 100 p. [ Links ]
Ramírez-Meneses, A.; García-López, E.; Obrador-Olán J.J.; RuizRosado, O. y Camacho-Chiu, W. 2013. Diversidad florística en plantaciones agroforestales de cacao en Cárdenas Tabasco, México. Universidad y Ciencia. 29(3):215-230. [ Links ]
Ramos, R. R. 2001. Análisis del uso sustentable de los suelos con plantaciones de cacao (Theobroma cacao L.) en Tabasco, México, aplicando sistemas de información geográfica. Tesis Maestría en Ciencias Colegio de Postgraduados Campus, Montecillo. Edo. México. 146 p. [ Links ]
Roa, R. H. A.; Salgado, M. M. G. y Álvarez, H.J. 2009. Análisis de la estructura arbórea del sistema agroforestal de cacao (Theobroma cacao L.) en el Soconusco, Chiapas, México. Acta Biológica Colombiana. 14(3):97-110. [ Links ]
Rosa, S. R. H. 2003. Ecología da vegetação arbórea de cabruca - mata atlântica raleada utilizada para cultivo de cacau - na região sul da Bahia. Universidad de Brasília. 161 p. [ Links ]
SAGARPA. 2010. Impulsa SAGARPA producción de cacao. Consultado: http://www.actualidadesmexico.com.mx/2010/03/impulsasagarpa-produccion-de-cacao/.24/03/11. [ Links ]
Salgado, M. M. G.; Ibarra, N. G.; Macías, S. J. E. y López, B. O. 2007. Diversidad arbórea en cacaotales del Soconusco, Chiapas, México. Interciencia. 32(11):763-768. [ Links ]
Scheelje, B. J. M. 2009. Índice de la legislación sobre el aprovechamiento del recurso maderable en sistemas silvopastoriles de Costa Rica. CATIE. 177 p. [ Links ]
SEDAFOP y OEIDRUS. 2008. Padrón de productores de cacao actualizado hasta mayo 2008. 114 p. Consultado: Consultado: http://www.campotabasco.gob.mx/ . 16/01/12. [ Links ]
Zarco, E. V. M.; Valdez, H. J. L.; Ángeles, P. L. y Castillo, A. O. 2010. Estructura y diversidad de la vegetación arbórea del parque estatal agua blanca, Macuspana, Tabasco. Universidad y Ciencia. 26(1):1-17. [ Links ]
Received: December 2015; Accepted: March 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
