










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.7 spe 13 Texcoco Jan./Feb. 2016
Articles
Presence of parthenocarpic fruit in ‘Ataulfo’ mango and its relation with environmental temperature and fertilization treatments
1 Campo Experimental Santiago Ixcuintla-INIFAP. A. P. 100, Santiago Ixcuintla, Nayarit 63300, México.
2 Investigador(a) independientes. Santiago Ixcuintla, Nayarit, México.
3 Universidad Autónoma de Nayarit-Unidad Académica de Agricultura. A. P. 49, Xalisco, Nayarit 63780, México.
In the state of Nayarit, Mexico, 'Ataulfo' shows high production of parthenocarpic fruits and there is little information on the causes of this anomaly. The objectives of this study were 1) to quantify temperature occurrence suggested as harmful in pre- and post-anthesis; and 2) to evaluate the influence of soil fertilization on the presence of parthenocarpic fruit. The research was conducted in two “Ataulfo” commercial orchards located in a warm humid climate from the coast of Nayarit. Environmental temperatures were recorded every 15 min during the 2011-2012, 2012-2013 and 2013-2014 cycles and were associated with values reported as harmful to flowering and fruit set at different stages of floral development and post-anthesis weeks. Two levels of balanced fertilizer (N, P, K, Ca, Mg, Fe, Mn, Zn and B) were evaluated, based on nutritional requirements and nutritional status of the tree, soil fertility and fertilization efficiency. The witness did not receive fertilizer. The presence of temperatures ≤13 °C, ≤14 °C and ≤15 °C, at flowering, mainly in the states of floral development E-8 (elongation of the main axis form the panicle - cauliflower state; the integuments are being formed in the egg, microspore stem cells are visible in the anther) and E-9 (elongation of the secondary and tertiary axes; meiosis has occurred, microspore tetrad and the integument of the ovule are formed), seems to be the responsible of parthenocarpic fruit in 'Ataulfo' mango, as it was not found any evidence of stenospermocarpy. Fertilization treatments did not affect the proportion of parthenocarpic mango that reached harvest maturity.
Keywords: Mangifera indica L.; climate change; ecophysiology; parthenocarpy; stenospermocarpy
En el estado de Nayarit, México, el mango ‘Ataúlfo’ presenta elevada producción de frutos partenocárpicos y existe poca información sobre las causas de esta anomalía. Los objetivos de este estudio fueron: 1) cuantificar la ocurrencia de temperaturas sugeridas como dañinas en pre- y post-antesis; y 2) evaluar la influencia de la fertilización al suelo sobre la presencia de fruto partenocárpico. La investigación se realizó en dos huertos comerciales de ‘Ataúlfo’ ubicados en el clima Cálido subhúmedo de la costa de Nayarit. Se registraron temperaturas ambientales cada 15 min en los ciclos 2011- 2012, 2012-2013 y 2013-2014 y fueron asociadas con valores mencionados como dañinos para floración y/o cuajado de fruto en varias etapas de desarrollo floral y semanas post-antesis. Se evaluaron dos niveles de fertilización balanceada (N, P, K, Ca, Mg, Fe, Mn, Zn y B), basada en los requerimientos y condición nutrimental del árbol, fertilidad del suelo y la eficiencia de la fertilización. El testigo no recibió fertilizante. La presencia de temperaturas ≤13 °C, ≤14 °C e ≤15 °C en prefloración, principalmente en los estados de desarrollo floral E-8 (elongación del eje primario de la panícula -estado coliflor; los tegumentos se están formando en el óvulo, las células madre de la microspora son visibles en la antera) y E-9 (elongación de los ejes secundarios y terciarios; la meiosis ha ocurrido, la tétrada de la microspora y los tegumentos del óvulo se han formado), parece ser las causante de frutos partenocárpicos en el mango ‘Ataúlfo’ ya que no se encontraron evidencias de estenospermocarpia. Los tratamientos de fertilización no modificaron la proporción de mango partenocárpico que alcanzó la madurez de cosecha.
Palabras clave: Mangifera indica L.; cambio climático; estenospermocarpia; ecofisiología; partenocarpia
Introduction
'Ataulfo' mango is the most important in the state of Nayarit as it occupies close to 11 000 of the 25 000 hectares planted with mango. Yields of this cultivar has decreased as in 2004 it was 14.9 t ha-1 and in 2014 was 7.1 t ha-1 (SAGARPA-SIAP, 2015). This decrease has been associated with a greater presence of seedless fruit that does not reach a commercial size, known locally as "mango niño" (Pérez-Barraza et al., 2007).
According to the statement made by Gehrke-Velez (2008) it may occur simultaneously the “mango niño” and the so called "parthenocarpic mango.” The first case refers to underdeveloped fruits showing a slit on the back and sharp peak. The fruit contains exosperm (seed coat) and sometimes you can be seen remnants of stunted embryo. Other authors point them out as stenospermocarpy fruits; i.e. small seedless fruit caused by the abortion of the embryo after fecundation (Stout, 1936; Soule, 1985). In Australia, the latter are named "nubbins" (Whiley et al., 1988). On the contrary, the term parthenocarpy describes the development of seedless fruit in which fecundation did not occur (Ledbetter and Ramming, 1989).
Low temperatures have been associated with the production of both types of fruits in mango. Lakshminarayana and Hernandez-Aguilar (1975) mentioned that temperatures from 8 to 10 °C during flowering of cv. Haden (monoembryonic) gave 67 and 52% of parthenocarpic fruits in the northern state of Sinaloa, Mexico. Similar observations were made for the same cultivar in Israel (Kadman and Oppenheimer, 1978). Temperatures ≤12 °C during floral differentiation and floral opening in cv. Irwin (monoembryonic) gave an abnormal development of flowers and deformed fruit production and seeded (Lim et al., 1996). In a similar study in 'Kensington' mango (polyembryonic), Whiley et al. (1988) also found that temperatures ≤12 °C during pollination and fruit set resulted in the presence of seedless fruit.
Sukhvibul et al. (2000) found that at 10 °C, in vitro germination of grain pollen for cvs. Nam Dok Mai, Kensington and Irwin was 53.9%. The percentage increased from 76.2 to 77.4% between 15 and 25 °C and decreased to 68.2% at 30 °C. The number of pollen tubes that reached the egg was higher between 20 and 25 ° C and stopped at 10 °C. Temperatures of 20/10 °C (day / night) for three days after pollination significantly increased the presence of stenospermocarpy fruits: 38.3, 21.4 and 6.8% in 'Nam Dok Mai', 'Kensington' and 'Irwin', respectively. Subsequently, Sukhvibul et al. (2005) mentioned that at the same temperatures, the proportion of seedless mango for the same cultivars was 21, 11 and 3%, respectively.
Nutritional deficiencies in mango have also been associated with poor pollen germination (de Wet et al., 1989). However, Singh (2007) did not correlate this condition with embryo abortion in mango.
In Mexico, climate change has worsened the production of mango "niño" in cv. Ataúlfo. In the state of Nayarit are becoming more frequent polar air masses, known as "cold fronts", causing atypical drops in temperature that can affect the sensitive stages of flower development and fruit set. There is information on the magnitude of the problem on mango niño; however, it is necessary to know the influence of temperature and nutritional status of the orchard on the presence of seedless mango and determine whether it is a phenomenon of parthenocarpy or stenospermocarpy. The objectives of this study were 1) to quantify the occurrence of temperatures mentioned as harmful in pre- and post-flowering in 'Ataulfo' mango; and 2) to evaluate the influence of soil fertilization on the presence of parthenocarpic fruits.
Material and method
Orchards characteristics: the research was conducted from 2010 to 2014 in two commercial orchards of 'Ataulfo' located in the municipalities of San Blas and Compostela, Nayarit. In San Blas, the selected orchard was "Las Palmas" (21° 36' 46.1" north latitude, 105° 11' 19.6" west longitude) and in Compostela was "El Divisadero" (21o 07' 03.0" north latitude, 105° 11' 04.6" west longitude) at an altitude of 193 and 104 m, respectively. The orchards had no irrigation and climate is warm humid AW1(W) (Garcia-Amaro,1988). The average annual temperature and the coldest month exceed 22 and 18 °C, respectively. The rainy season is from June to October, and precipitation in the driest month is less than 60 mm. In both orchards the planting arrangement was 8 x 8 m, 10 years old on "Las Palmas" and 12 in "El Divisadero" and Acrisol humic soil and Foezem haplic respectively (INEGI, 1999).
Quantification of critical temperature (≤ 17 °C) according to growth stage
Temperature record was performed for the 2011-2012, 2012-2013 and 2013-2014 cycles. The information was obtained from the weather stations located in Las Palmas, municipality of San Blas (20° 36' 18.2'' north latitude, 105° 08' 35.6'' west longitude) and Las Varas, municipality of Compostela, Nayarit (21° 11' 30.3'' north latitude, 105° 08' 50.9'' west longitude).
The quantified temperatures at flowering were ≤ 17°, ≤ 16°, ≤ 15°, ≤ 14°, ≤ 13° and ≤ 12 °C. The stages of flower development (E) were: E-8 (elongation of the primary axis of the panicle, secondary axes still covered by their bracts, there are small and immature flowers; the integuments are forming in the egg, stem cells from microspore are visible in the anther) to E-9 (the primary axes continues its elongation and the elongation of the secondary and tertiary axes are appreciated - which bear the flowers; meiosis has occurred, microspore tetrads and the integument of the egg are formed), E-9 and E-10 (elongation of the tertiary axis and presence of full bloom but unopened. Microsporogenesis has generated pollen grain), E-10 to E-11 (mature and open f lower; the stigma is receptive and dehiscence occurs) (Salazar-García, Per. Com.). In post- flowering the same temperatures were measured during weeks 1, 2, 3 and 4 after anthesis (SDA). For each orchard total hours accumulated per temperature and phenological stage was obtained.
Fertilization treatments and presence of parthenocarpic fruit
Fertilization treatments were applied from 2010 to 2013 in 'Ataulfo' orchards already described. In each orchard selected 60 trees, which were divided into groups of 20 and applied in summer 2010, 2011, 2012 and 2013 three fertilization levels: 1) normal doses, which considered demand and foliar nutritional condition of the tree, nutrients provided by the soil and fertilization efficiency and consisted in the application per tree, depending the orchard, from 349 to 360 g N, 139 to 147g P2O5, 163-296 g K2O, 13 to 24 g Mg, 11 to 12 g Fe, 6 to 12 g Mn, 2 to 5 g Zn and 4 to 7 g B; and 2)1.5 times the normal dose; and 3) control, without fertilization. Fertilizers were fractionated in July and September and distributed at 1.5 m from the trunk of the tree in a band of 50 cm wide and 20 cm deep.
Fruit count: were made in 2012, 2013 and 2014, to evaluate the effect of fertilization from 2011, 2012 and 2013, respectively; conducted in March (fruits of 3-4 cm length) and May (fruit at cut maturity) counting normal and parthenocarpic fruits that were inside a wire frame of 50 x 50 cm placed in each of the cardinal points from each tree.
Macroscopic characteristics of parthenocarpic fruits: each year of the study, in each orchard and from each fertilization treatment, 50 fruits whose external features corresponded to normal and parthenocarpic fruits were collected. The fruits were sectioned longitudinally to verify the presence of embryos under a stereoscopic microscope (Zeiss Stereomikroskop Model Stemi 2000-C. Carl Zeiss. Göttingen, Germany) and photographed with a digital camera (Canon Power Shot G11. NY. USA).
Data analysis
Temperature data was analyzed as factorial considering the information from the two orchards, production cycles, growth stages and thermal thresholds in order to look for differences in the number of hours and thus determine if analyzed separately or as a single database data.
The incidence of normal and parthenocarpic fruit was analyzed with the method of repeated measures with factorial arrangement 3 x 2 x 3, years (2012, 2013 and 2014), orchards (Las Palmas and El Divisadero) and fertilization treatments (normal dose, high dose and control). The experimental design was completely random. SAS statistical software V. 9.1 (2009) was used. Field data obtained were expressed as a percentage of normal and parthenocarpic fruit per tree. Prior to analysis of variance the values in percentages were transformed by the square root of X + 0.5 (Steel and Torrie, 1984). The comparison of means was made with Waller- Duncan test, p≤ 0.05.
Results
Accumulation of critical temperatures (≤17 °C) according to growth stage
In Las Palmas cumulative hours ≤17 °C was significantly higher for growing stage from E-8 (cauliflower) to E-11 (anthesis); ranging from 99.2 to 146.2 h (Table 1). For temperatures ≤16 °C it was higher (87.8 h) from E-9 to E-10, although it was not different to that accumulated from E-8 to E-9 and E-10 to E-11. Regarding to the comparison of cumulative hours with the different critical temperatures (from ≤12 °C to ≤17 °C) were significantly higher the accumulation of hours with temperatures ≤17 ° C (Table 1). This proved to be the case for developmental stages from E-8 to E-11 (Table 1).
Table 1 Average (2012, 2013 and 2014) of accumulated hours with different critical temperaturas according to the f loral stage of development (E) and weeks after anthesis (SDA) in orchard Las Palmas, Municipality of San Blas, Nayarit.
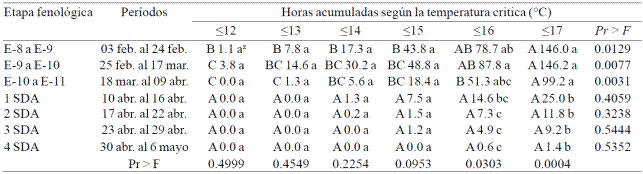
zPara letras mayúsculas y minúsculas la separación de medias es horizontal y vertical, respectivamente, según la prueba de Waller-Duncan p= 0.05.
For El Divisadero cumulative hours from ≤14 °C to ≤17 °C, showed differences between stages; however, in all cases were higher than that recorded after anthesis (SDA) (Table 2). Most hours with accumulated critical temperatures occurred in the phenological stages from E-8 to E-9 and E-9 to E-10 and were temperatures ≤15, ≤16 and ≤17 °C. Values ranged from 46.7 h (≤15 °C) to 140.4 h (≤17 °C) (Table 2). As for accumulation of hours with calculated critical temperatures from ≤12 °C to ≤17 °C the numbers were higher for ≤16 °C (phenological stages from E-8 to E-9 and E-9 to E-10) and ≤17 ° C (phenological stages from E-8 to E-11) (Table 2).
Table 2 Average (2012, 2013 and 2014) of accumulated hours with different critical temperatures according to the f loral stage of development (E) and weeks after anthesis (SDA) in orchard El Divisadero, Municipality of Compostela, Nayarit.

zPara letras mayúsculas y minúsculas la separación de medias es horizontal y vertical, respectivamente, según la prueba de Waller-Duncan, p= 0.05.
Due to variation between years of the occurrence of critical temperatures for reproductive development of 'Ataulfo', the minimum and maximum durations of each critical temperature for the three years of research were calculated. In Las Palmas, the lowest temperatures (≤12 °C) occurred in the phenological stages from E-8 to E-10 with a maximum of 11:15 hours in the period from February 3 to March 17 (Table 3). Critical temperatures ≤13 °C and up to ≤17 °C, although of varying lengths, always occurred in developmental stages from E-8 to E-10. However, temperatures ≤13 °C lasted over 20 h in the states of floral development E-8 to E-10. Critical temperatures ≤14 °C and higher showed prolonged durations, of up to 193:75 h for ≤17 °C in stages from E-8 to E-9; the duration of critical temperatures in post-anthesis stage was extended generally from ≤15 °C in stage 1 SDA (22:30 h).
Table 3 Inter-annual variation (2012, 2013 and 2014) in the duration of occurrence for different critical temperatures according to the floral stage of development (E) and weeks after anthesis (SDA) in orchards Las Palmas y el Divisadero.

For El Divisadero, the duration of minimum and maximum temperatures for each critical temperature in the three years evaluated was higher than in Las Palmas (Table 3); in the phenological stages from E-8 to E-10 (February 3 to April 9) it was notorious the occurrence of up to 26:30 h at temperatures ≤12 °C. The presence of temperatures ≤13 °C and higher was always high; for example, the variation between years for the duration of temperature ≤15 °C in stages from E-8 to E-9 was from 23:30 h to 63:15 h in the period from February 3 to February 24. For E-9 to E-10 (February 25 to March 17) the duration of temperatures ≤15 °C ranged from 48:15 to 88:30 h. The duration of the critical temperatures after anthesis reached up to 30 h at temperatures ≤15 °C in 1 SDA (April 10 to April 16) (Table 3).
Fertilization treatments and incidence of parthenocarpic fruit
The proportion of normal and parthenocarpic fruit varied between years of evaluation in trees treated with different levels of fertilization (Table 4). In the assessment made in fruit set (March), normal fruits reached a maximum of 27.1% (2014) and parthenocarpic fruits of up to 94.6% (2012). For fruit that reached harvest an inverse relationship between the amount of normal and parthenocarpic fruits was observed. The variation between years was 14% for each type of fruit; although the proportion of parthenocarpic fruit was high (43.5% to 57.5%) (Table 4).
Table 4 Influence of the year of assessment on the presence of normal and parthenocarpic fruit in ‘Ataulfo’ treated with three soil fertilization treatments. Evaluations made in fruit set (March) and maturity index (May) 2012, 2013 and 2014. Orchards Las Palmas and El Divisadero

zNúmero de frutos evaluados; y Medias con letras iguales en columnas no son estadísticamente diferentes, según la prueba de Waller-Duncan, p= 0.05.
The comparison between orchards for presence of parthenocarpic fruit in fruit set (March) and maturity index (May) showed a greater presence of this type of fruit in orchard El Divisadero, reaching 91% in fruit set and 63.7% to harvest (Table 5). For Las Palmas these ratios were lower, 72.2% and 40.1%, respectively.
Table 5 Differences between orchards on presence of normal and parthenocarpic fruit in ‘Ataúlfo’ treated with three soil fertilization treatments. Evaluations in fruit set (March) and maturity index (May) 2012, 2013 and 2014.

zNúmero de frutos evaluados; y Medias con letras iguales en columnas no son estadísticamente diferentes, según la prueba de Waller-Duncan, p= 0.05.
Fertilization treatments modified the presence of parthenocarpic fruit. However, this only happened for the evaluation in fruit set (Table 6). Normal fertilization doses had the lowest proportion of parthenocarpic fruit (79.5%) compared to control (83.5%).
Table 6 Influence of soil fertilization treatments on the presence of normal and parthenocarpic fruit in ‘Ataulfo’. Evaluations in fruit set (March) and maturity index (May) 2012, 2013 and 2014. Combined data from orchards Las Palmas and El Divisadero.

zNúmero de frutos evaluados; y Medias con letras iguales en columnas no son estadísticamente diferentes, según la prueba de Waller-Duncan, p= 0.05.
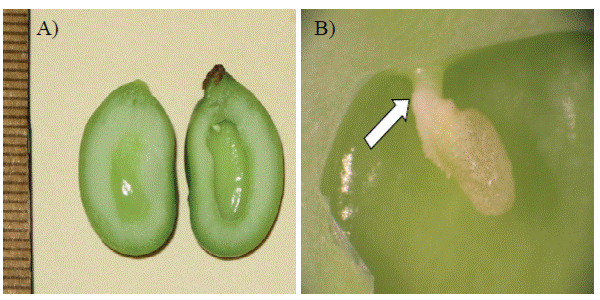
Regarding fruit characteristics, the normal always were larger than parthenocarpic (Figures 1A and 1B). Parthenocarpic fruit showed no slit in the back or a pronounced peak and no develop embryos, only rudiments of the embryo sac (Figures 2A and 2B) were observed. In all evaluated fruits was not observed aborted embryos.

Figure 1 External appearance of normal and parthenocarpic fruit of ‘Ataulfo’ (A) normal fruit with presence of embryos (left) and parthenocarpic, showing no embryos; and (B).
Discussion
Cumulative hour ≤17 °C was higher in El Divisadero than in Las Palmas. This difference may be due to El Divisadero is at latitude 29' further south and 4.7 km from the Pacific Ocean while Las Palmas is 10.5 km from it; then the cold wind from the ocean impacts more directly into the orchard El Divisadero. The difference in altitude between both orchards was minimal because Las Palmas is 89 m higher and it seems not to have influenced the difference in the accumulation of hours ≤17 °C.
In both orchards was notorious the cumulative hours ≤17 °C during stages of floral development from E-8 (cauliflower) to E-11 (anthesis). Studies with several mango cultivars have mentioned the presence of seedless fruit as a result of damage caused by temperatures ≤10 °C (Lakshminarayana and Hernandez-Aguilar, 1975; Kadman and Oppenheimer, 1978; Sukhvibul et al., 2000) or ≤12 °C (Whiley et al., 1988; Lim et al., 1996) during the stages of floral differentiation and pollination. During this study temperatures ≤10 °C or ≤11 °C were not recorded; however, there was accumulation of hours with temperatures from ≤12 °C to ≤17 °C, though depending on the orchard, it was greater the accumulation of temperatures ≤14 °C ≤15 °C and ≤16 °C. The most frequent states of floral development were this temperatures are present were from E-8 to E-9 and E-9 to E-10. These stages of development are very sensitive to damage from low temperatures as it is when egg and pollen formation occurs, as well as maturation.
As mentioned, the states of floral development E-8 and E-9 are the most susceptible to damage from low temperatures. In this regard, for both orchards the duration of temperatures ≤13 °C was more than 18 h, ≤14 °C was greater than 37 h, and ≤15 °C greater than 63 h. There was no information to be found regarding this temperatures for any mango cultivar; so the information described here indicates an increased susceptibility of the reproductive organs from 'Ataulfo' and will stimulate further research in this area, especially in topics related to vulnerability to change climate from current producing areas of this cultivar in Nayarit and possibly in other regions.
According to Sukhvibul et al. (2000), the presence of temperatures from 20/10 °C (day / night) for three days after pollination caused stenospermocarpy. None of the orchards included in this study showed similar temperature drops; however, a week after anthesis the duration of ≤15 °C temperatures in the orchards Las Palmas and El Divisadero was just for 22:30 h and 30 h, respectively. The absence of harmful temperatures for embryos may have been the cause that the abortion of embryos has not been detected in dissected fruits during the three years of study.
Overall, the proportion of parthenocarpic fruit that reached harvest in the three years evaluated ranged from 43.5 to 57.5%, which means about 50% reduction in yield potential for 'Ataulfo' in Nayarit. The lack of effect of balanced fertilization treatments, according to the requirements of each orchard is an indicator that conditions where this research was conducted for the presence of parthenocarpic fruit is from climate.
Conclusions
The results from this study indicate that the presence of temperatures ≤13 °C, ≤14 °C e ≤15 °C at pre-flowering, mainly in the stages of floral development E-8 and E-9, are the responsible for the presence of parthenocarpic fruit in ‘Ataulfo’ and there was no evidence of stenospermocarpy. The fertilization treatments did not modify the proportion of parthenocarpic fruit reaching harvest.
Literatura citada
De Wet, E.; Robbertse, P. J. and Groeneveld, H. T. 1989. The influence of temperature and boron on pollen germination in Mangifera indica L. South African J. Plant Soil. 6(4):228-234. [ Links ]
Gehrke-Vélez, M. L. 2008. Reflexiones sobre problemas de biología reproductiva del mango Ataúlfo en el Soconusco, Chiapas. Tecnología en Marcha. 21:174-183. [ Links ]
García-Amaro, E. 1988. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). ‘Climas’ (clasificación de Köppen, modificado por García). Escala 1:1000 000 México. Shapefile. http://www.conabio.gob.mx/informacion/metadata/gis/clima1mgw.xml?_httpcache=yes&_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no. [ Links ]
INEGI. 1999. Carta edafológica del estado de Nayarit. Dirección General de Geografía del Instituto Nacional de Estadística Geografía e Informática (INEGI). 2nd. Ed. Escala 1:50 000. Aguascalientes, Aguascalientes, México. [ Links ]
Kadman, A. and Oppenheimer, Ch. 1978. Some problems of cultivation of selected subtropical fruit crops under marginal conditions, and research carried out in Israel to solve these problems. Acta Hortic. 142:19-21. [ Links ]
Lakshminarayana, S. and Hernández-Aguilar, P. 1975. Effect of orchard heating in reducing parthenocarpic fruits in `Haden' mango. United States of America. Proc. Fla. State Hort. Soc. 88:502- 505. [ Links ]
Ledbetter, C. A. and Ramming, D. H. 1989. Seedlessness in grapes. In: horticultural reviews. Janick, J. (Eds.). John Wiley & Sons, Inc. Hoboken, NJ. United States of America. 11:159-181. [ Links ]
Lim, T. K.; Luders, L. and Poffley, M. 1996. Studies on fruit deformity and nubbins in Irwin mango. In: Proc. Int. Conference on Tropical fruit. Kuala Lumpur, Malaysia. 287-298 pp. [ Links ]
Pérez-Barraza, M. H.; Vázquez-Valdivia, V. y Osuna-García, J. A. 2007. Incidencia de frutos paternocárpicos en mango ‘Ataulfo’ en huertos comerciales de Nayarit. México. Rev. Chapingo Serie Horticultura. 13(2):149-156. [ Links ]
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA)- Servicio de Información y estadística Agroalimentaria y Pesquera (SIAP). 2015. http://www.siap.gob.mx/. [ Links ]
Sing, Z. 2007. Embryo abortion in relation to fruit size, quality, and concentrations of nutrients in skin and pulp of mango. J. Plant Nutrit. 28:1723-1737. [ Links ]
Soule, J. 1985. Glossary for horticultural crops. Wiley, New York, United States of America. 898 p. [ Links ]
Steel, R. G. D. and Torrie, J. H. 1984. Principles and procedures of statistics a biometrical approach. 2th. Ed. McGraw Hill Publishers. 234-235 pp. [ Links ]
Stout, A. B. 1936. Seedlessness in grapes. Technical bulletin 238. New York State Agricultural Experiment Station. Geneva, New York, United States of America. 35 p. [ Links ]
Sukhvibul, N.; Hetherington, S. E.; Whiley, A. W.; Smith, M. K. and Vithanage, S. 2000. Effect of temperature on pollen germination, pollen tube growth and seed development in mango (Mangifera indica L.). Acta Hortic. 509:609-616. [ Links ]
Sukhvibul, N.; Whiley, A.W. and Smith, M. K. 2005. Effect of temperature on seed and fruit development in three mango (Mangifera indica L.) cultivars. Scientia Hortic. 105:467-474. [ Links ]
Whiley, A. W.; Saranah, J. B.; Rasmussen, T. S.; Winston, E. C. and Wolstenholme, B. N. 1988. Effect of temperature on ten mango cultivars with relevance to production in Australia. In: Proc. Fourth Australasian Conference on Tree and Nut Crops. Lismore, New South Wales. 176-185 pp. [ Links ]
Young, T. W. and Sauls, J. W. 1979. The mango industry in Florida. Florida Cooperative Extension Service Bulletin 189. 70 p. [ Links ]
Received: November 2015; Accepted: February 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
