











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La fotosíntesis está relacionada directa o indirectamente con muchos procesos finamente regulados en las células y variaciones en ellos se reflejan en cambios en la fisiología de la planta y finalmente en su crecimiento y supervivencia. Diversos estudios han reportado que la exposición a condiciones subóptimas o condiciones de estrés (agobio hídrico, temperaturas extremas, exceso de radiación solar, estrés mineral, entre otras) resultan en una disminución de la fotosíntesis como resultado de la supresión o activación de ciertas rutas metabólicas (Rakic, 2015). Por ejemplo, la disminución en la cantidad de radiación solar, y en general bajo cualquier tipo de estrés se reduce la actividad fotosintética al disminuir el contenido de clorofila (Cambrón-Sandoval et al., 2011).
Los metabolitos secundarios son fitoquímicos esenciales que son afectados por el espectro de luz y actúan como compuestos de defensa, así como de protección frente a la radiación ultravioleta y a agentes oxidantes. Aunado a lo anterior, ciertos compuestos fenólicos y flavonoides, muestran actividad microbiana, antioxidante, antifungosa y de atrapamiento de radicales libres (Ouzounis et al., 2015).
Los compuestos fenólicos, ampliamente distribuidos en las plantas, son producidos en las rutas del shikimato, las pentosas fosfato y fenilpropanoide; su importancia, además del papel que juegan en el crecimiento, reproducción y protección, radica en que también contribuyen en el color y características sensoriales de los frutos (Balasundram et al., 2006). En la piel de frutos de aguacate se han identificado diversos pigmentos, principalmente carotenoides, clorofilas y ciadinidas (Donetti y Terry, 2012). En los aguacates con piel verde el color es debido principalmente a la clorofila, mientras que en el caso de ‘Hass’, cuya piel se torna negra conforme madura, su coloración es causada en parte por el contenido de antocianinas (Ashton et al., 2006). El contenido de fitoquímicos en la piel de frutos de aguacate ha sido reportado para diferentes cultivares, destacando ‘Hass’(raza Guatemalteca x Mexicana) por su mayor concentración de compuestos fenólicos totales y menor de clorofilas A y B, comportamiento opuesto al cv. Tonnage (raza Guatemalteca x Antillana) (Wang et al., 2010).
La variación en las condiciones agronómicas (especie, cultivar, estado de desarrollo, órgano de la planta, competencia, fertilización, entre otras), la estación del año, factores climáticos, disponibilidad de agua, luz (intensidad, calidad y duración) tiene efectos significativos sobre el contenido y perfil de fitoquímicos en un cultivo (Björkman et al., 2011). El cultivo de aguacate prospera desde climas cálidos hasta templados, con un rango de temperatura para fotosíntesis de 10 a 35 °C y precipitación pluvial de 800 a 1 500 mm anuales (Ruíz et al., 2013), aunque con riego puede cultivarse en ausencia de lluvias.
Para la zona productora de ‘Hass’ en el clima semicálido del estado de Nayarit, fue elaborado un modelo de predicción del desarrollo f loral de brotes del flujo vegetativo de invierno el cual fue asociado a temperaturas ≤ 21 °C y los brotes del flujo de verano a temperaturas ≤ 19 °C (Salazar-García et al., 2007). Estos modelos fueron probados en tres tipos de clima del estado de Michoacán (cálido subhúmedo, semicálido subhúmedo y templado subhúmedo) y sólo funcionó el modelo para brotes del flujo vegetativo de verano (temperaturas ≤ 19 °C; Salazar-García et al., 2009). En otro estudio, Álvarez-Bravo y Salazar-García (2015) evaluaron un modelo de predicción del desarrollo f loral desarrollado para Michoacán (basado en días frío acumulados con temperaturas ≤ 16 °C), que mostró una buena capacidad predictiva en brotes del f lujo vegetativo de invierno de ‘Hass’ cultivado en seis tipos de clima de la región aguacatera de Michoacán. Lo anterior indica que existen diferentes umbrales de temperatura relacionados con el desarrollo floral del aguacate ‘Hass’. Por otro lado, en climas cálidos, la piel del fruto de ‘Hass’ frecuentemente presenta mayor rugosidad y espesor, lo cual suele ser tomado como argumento para disminuir el precio del fruto, respecto al proveniente de regiones más frescas en los que predominan frutos con piel más delgada y menos rugosa. Sin embargo, estas características parecen no afectar la calidad física, química u organoléptica del fruto (Salazar-García et al., 2016).
Los cambios en los componentes del sistema climático obedecen a una compleja relación entre agentes forzantes (tales como la producción de gases con efecto invernadero, cambios en la irradiación solar e insolación y cambios en la cobertura del suelo), procesos naturales y actividades humanas. Lo anterior conlleva a ciertas respuestas climáticas como la variación en la precipitación y temperatura (Pielke et al., 2007). Con el cambio climático, las variaciones en los parámetros agro-climáticos afectarán el funcionamiento celular, y por lo tanto, la fisiología de los cultivos, además de afectar la calidad de los frutos en términos de gusto, apariencia, sabor y contenido de nutrientes, fitoquímicos y otros componentes dietarios. Lo anterior puede deberse a una elevada fotooxidación, a un aumento en la síntesis de especies reactivas del oxígeno (ERO) y de metabolitos secundarios (Singh et al., 2015). El objetivo de este trabajo fue evaluar el efecto de la condición de humedad del suelo y la radiación solar incidente sobre el contenido de fitoquímicos en la piel de frutos de aguacate ‘Hass’.
Materiales y métodos
Características de los huertos. Fueron seleccionados dos huertos comerciales de aguacate ‘Hass’en el Ejido El Rodeo, municipio de Tepic, Nayarit, uno ubicado a una altitud de 950 m (sin riego) y el otro a 1 200 m (con riego), con clima Semicálido subhúmedo (A)C(w2) y tipo de suelo Andosol.
En el huerto con riego se tiene un microaspersor de 34 L h-1 por árbol y la frecuencia y lámina de riego es determinada mediante tensiómetros, procurando mantener la tensión de humedad del suelo entre 10 y 30 centibars lo que implica riegos de 3 a 4 h a intervalos de 14 a 21 días. La distribución de la precipitación pluvial en el huerto sin riego es de junio a septiembre, precipitándose en estos meses más de 1 000 mm lo que representa cerca de 90% anual.
Caracterización climática. Con datos provenientes de las normales climatológicas del servicio meteorológico nacional, se construyó un climograma mensual de la serie de tiempo 1981-2010 de la estación meteorológica 18 038 en Tepic, ubicada en las coordenadas latitud norte 21° 30' 00" y longitud oeste 104° 53' 00" con una altitud sobre el nivel del mar de 935 m.
Meteorología de la zona de estudio. Durante el tiempo que duró el estudio, mayo a septiembre 2014, en cada huerto se registró cada hora la temperatura ambiental con registradores automatizados HOBO H8 (Onset Computer, Witzprod, Englewood Cliffs, NJ, USA). Mediante el empleo del motor de base de datos Microsoft Access 2010 se integraron los datos de temperatura de los sensores de temperatura desde el inicio del periodo cálido y seco hasta cosecha (mayo- septiembre), calculándose la cantidad de días que superaron el promedio histórico mensual en temperatura máxima (Tmax) y mínima (Tmin). Asimismo, fueron cuantificados el máxima maximorum (Mmax) y mínima minimorum (Mmin) de temperatura por mes. Para el cálculo de las unidades calor acumuladas (UCA) por mes se utilizó la ecuación:

Colecta de frutos. En cada uno de los huertos se seleccionaron diez árboles y en cada árbol fueron marcados ocho frutos, cuatro para el tratamiento “cubierto” y cuatro para el tratamiento “descubierto”. Los frutos del tratamiento “cubierto” fueron cubiertos con un cono de papel de estraza, con la base descubierta, desde que alcanzaron el tamaño aceituna (2-3 cm de diámetro). Los frutos analizados fueron originados por la floración principal (invierno 2013) y cosechados en octubre 2014, con un contenido de materia seca en la pulpa ≥ 21.5%. Después de la cosecha, los frutos fueron llevados al Laboratorio de Frutifisiología del Campo Experimental Santiago Ixcuintla, donde fueron procesados. Los frutos fueron lavados con agua destilada con hipoclorito de sodio (200 mg L-1) y se pesaron en una balanza de precisión (Ohaus modelo P2001, Florham, NJ, USA.). La longitud y diámetro se obtuvieron con un vernier digital (MTC500-196, Mitutoyo Co., Japón). Posteriormente, la piel de los frutos fue separada de la pulpa con un pelador de verduras. La piel fue congelada a -20 °C hasta el momento de la preparación de los extractos, en los cuales se determinó la concentración de fitoquímicos.
Concentración de fitoquímicos en la piel. La concentración de compuestos fenólicos totales y clorofilas A, B y totales, en la piel de los frutos de ‘Hass’ se determinó espectrofotométricamente en cuatro repeticiones, cada una de las cuales consistió en tres frutos, cada uno de distinto árbol. La lectura de absorbancia se hizo por triplicado. Los compuestos fenólicos totales (CFT), se cuantificaron con la metodología propuesta por Rodríguez-Carpena et al. (2011): la extracción se realizó en 10 g de piel con 30 mL de una mezcla acetona: agua (70:10). Una vez homogeneizada, la mezcla fue centrifugada a 2 500 rpm durante 3 minutos a 4°C. El sobrenadante se colectó y el residuo se sometió a una extracción igual. Ambos extractos se combinaron. El contenido de compuestos fenólicos totales de cada extracto se determinó con el método de Folin-Ciocalteau propuesto por Soong y Barlow (2004) y modificado por Rodríguez-Carpena et al. (2011). La absorbancia fue medida a 765 nm y el contenido de compuestos fenólicos totales se calculó a partir de una curva estándar de ácido gálico y se reporta en mg equivalentes de ácido gálico por gramo de piel (mg EAG g-1 piel). Una parte del extracto obtenido para la determinación de compuestos fenólicos totales se incubó en oscuridad durante 48 h a 4°C para el análisis de clorofilas A, B y totales (CA, CB y CT, respectivamente). La muestra se centrifugó a 4 000 rpm y se midió la absorbancia del sobrenadante a 647 y 664 nm. La concentración de clorofilas A, B y totales (mg L-1), se determinó con las siguientes ecuaciones (Gandolfo- Wiederhold, 2008) y se reportan en mg g-1 piel:
CA= 12.64 Abs664 nm-2.99 Abs647 nm
CB= -5.60 Abs664 nm+23.26 Abs647 nm
CT= 7.04 Abs664 nm+20.27 Abs647 nm
Análisis estadístico. Se empleó un diseño experimental factorial 2 x 2, manejo del agua (con riego y sin riego) y cobertura del fruto (cubiertos y descubiertos). El tamaño del fruto, según su peso, longitud y diámetro, así como la concentración de fitoquímicos se sometió a análisis de varianza y prueba de medias (Duncan, p< 0.05) con el paquete estadístico SAS.
Resultados
Climatología
Del análisis climatológico de los años 1981-2010, se observó que la temperatura media mensual es superior a 15°C; el periodo más cálido ocurre entre mayo y septiembre con una Tmax media superior a 26°C; el periodo frío (diciembre a marzo) presenta Tmin media menor a 10 °C. Durante junio-septiembre se precipita 86% de la lluvia (1 100 mm) y el mes más lluvioso supera 400 mm (julio). Las medias anuales de temperatura máxima y mínima son 27.1 y 13.6°C, respectivamente, mientras que la precipitación anual es de 1 286 mm (Figura 1).
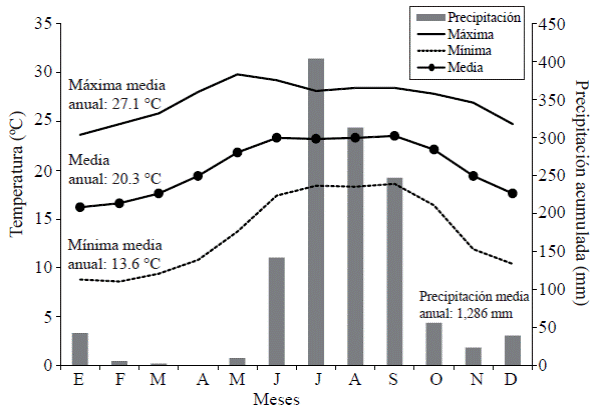
Meteorología del periodo de estudio (mayo-septiembre 2014)
Se cuantificaron los días que superaron la temperatura media histórica (Tmax) y los días con temperatura inferior a la media histórica (Tmin). En el huerto con riego se acumularon 37 días con temperatura superior a la media, mientras que en el huerto sin riego fueron 85 días. En contraste, los días acumulados en los que la temperatura fue inferior a la temperatura media fueron 47 y 17 días para los huertos con y sin riego, respectivamente. El huerto sin riego fue más caliente (mayor cantidad de días cálidos y menor en días frescos) que el que tuvo riego. La comparación entre huertos mostró que donde hubo riego se acumularon mensualmente menos días con Tmax superior a la media y un mayor número de días con Tmin menor a la media; mientras que Mmax fue superior en el huerto sin riego, fluctuando entre 0.7 y 1.8 °C; en Mmin el huerto con riego fue el más fresco, con diferencias entre -0.2 y -1.5 °C. En el huerto sin riego se acumularon 2141.6 unidades calor, siendo mayores a las del huerto con riego (1990.7). Cuadro 1.

Tamaño del fruto
En el huerto con riego, el peso, longitud y diámetro del fruto fueron mayores en los frutos descubiertos que en los cubiertos con papel. En el caso del huerto sin riego, el tamaño del fruto no fue afectado por la cobertura con papel (Cuadro 2).
Cuadro 2 Tamaño del fruto de aguacate ‘Hass’ con diferente grado de exposición a la luz y manejo del agua.

zMedias con la misma letra en una columna no presentan diferencia significativa, de acuerdo con la prueba de Duncan, p≤ 0.05.
El manejo del agua afectó el tamaño de los frutos descubiertos. Los frutos del huerto con riego presentaron mayor peso y longitud que los del huerto sin riego. Sin embargo, el diámetro del fruto no fue afectado por el manejo del agua (Cuadro 3).

Compuestos fitoquímicos
En frutos descubiertos, tanto del huerto con riego como sin él, se presentó una mayor concentración de compuestos fenólicos totales, así como de las clorofilas A, B y totales en la piel (Cuadro 4). Por otra parte, en frutos descubiertos, el manejo del agua no afectó la concentración de los compuestos fitoquímicos analizados (Cuadro 5).
Cuadro 4 Concentración de fitoquímicos en la piel de frutos de aguacate ‘Hass’con diferente grado de exposición a la luz y manejo del agua.
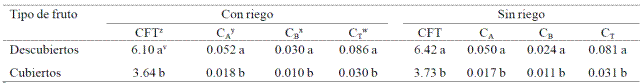
zCFT= compuestos fenólicos totales, mg equivalentes de ácido gálico (EAG) g-1 piel. yCA= clorofila A, mg g-1 piel; xCB= clorofila B, mg g-1 piel; wCT= clorofila total, mg g-1 piel; vMedias con la misma letra en una columna no presentan diferencia significativa, de acuerdo con la prueba de Duncan, p≤ 0.05.
Cuadro 5 Concentración de fitoquímicos en la piel de frutos de aguacate ‘Hass’expuestos a la luz y con diferente manejo del agua.

zCFT= compuestos fenólicos totales, mg equivalentes de ácido gálico (EAG) g-1 piel. yCA= clorofila A, mg g-1 piel; xCB= clorofila B, mg g-1 piel; wCT= clorofila total, mg g-1 piel; vMedias con la misma letra en una columna no presentan diferencia significativa, de acuerdo con la prueba de Duncan, p≤ 0.05.
Según el análisis factorial, el grado de exposición a la luz fue el factor con alta significancia estadística (p< 0.01) para todas las variables analizadas en el modelo. La interacción entre ambos factores, exposición a la luz y manejo del agua, solo resultó significativa para el peso y longitud de los frutos (Cuadro 6).

Discusión
Las condiciones climáticas de los sitios donde se desarrolló el presente trabajo coinciden con la descripción de temperatura del clima tipo Semicálido descrito por García (1964), donde la media anual y la del mes más cálido superan el umbral térmico de 18 °C y 22 °C, respectivamente. Estas condiciones climáticas coinciden con trabajos previos en la región de estudio, donde se determinó la presencia de días frío favorables para el desarrollo reproductivo del aguacate ‘Hass’ (Salazar-García et al., 2007; Salazar-García et al., 2009).
La condición de humedad afectó el tamaño del fruto ya que el huerto con riego produjo frutos con mayor peso y longitud que el huerto sin riego. Esto difiere con lo encontrado por Salazar-García et al. (2011) en Michoacán donde no hubo diferencia en el peso de frutos de ‘Hass’ provenientes de huertos con riego y sin riego. Esta divergencia de resultados podría ser atribuida a que en Michoacán los frutos se desarrollan en climas con mayor precipitación acumulada anual y una más amplia distribución de la lluvia durante el año, lo que podría ocasionar un menor estrés hídrico en el cultivo.
La concentración de compuestos fenólicos totales en la piel de frutos descubiertos, resultó más baja que lo reportado por Wang et al. (2010), quienes realizaron la extracción de fitoquímicos con una mezcla de acetona: agua: ácido acético (70:29.7:0.3, v/v/v), lo cual podría explicar la diferencia entre sus resultados y los de esta investigación en la que el solvente de extracción fue una mezcla acetona: agua (70:10 v/v). Ciertos grupos de fenólicos, tales como los flavonoides, actúan como mecanismo de protección contra la radiación ultravioleta (Cheynier et al., 2013), de ahí que los frutos expuestos a la luz hayan sido los que presentaron una concentración mayor de fenólicos totales. En uva (Vitis vinifera), los racimos descubiertos presentaron contenidos más altos de CFT, comparado con frutos que fueron cubiertos con redes que retenían parte de la luminosidad (Leguizamón-M et al., 2008).
El contenido de clorofilas, menor en los frutos cubiertos, evidenció la condición de estrés a la que fueron sometidos dichos frutos. En los frutos descubiertos el contenido de clorofilas A, B y totales, fue más alto que lo encontrado por Wang et al. (2010), en frutos de ‘Hass’, quienes para estos pigmentos reportaron 0.019, 0.010 y 0.029 mg g-1 piel, respectivamente; esta diferencia puede explicarse debido a que Wang et al. (2010) analizaron frutos en madurez de consumo, mientras que en la presente investigación el análisis se realizó en madurez fisiológica. Cox et al. (2004), reportaron en ‘Hass’ una disminución en el contenido de clorofilas entre los frutos de color verde bosque y los de color oscuro.
El contenido de metabolitos secundarios suele presentar variaciones estacionales (Scogings et al., 2015), lo que podría explicar las diferencias entre los resultados de este estudio y los previamente mencionados. Al obtener la relación clorofila A/B en frutos descubiertos (1.73 y 2.08, huerto con y sin riego, respectivamente) y en frutos cubiertos (1.80 y 1.54, huerto con y sin riego, respectivamente) se encontró que fue menor en estos últimos lo cual fue similar a lo encontrado en pino (Pinus pseudostrobus) y asociado a una disminución en la cantidad de luz (Cambrón-Sandoval et al., 2011).
Conclusiones
El manejo de agua en huertos de aguacate ‘Hass’ no modificó el contenido de fitoquímicos en la piel de los frutos. Respecto al grado de exposición a la radiación solar, los frutos descubiertos presentaron una concentración mayor de compuestos fenólicos totales y clorofilas, comparados con aquellos que se sometieron a estrés por falta de luz. Como la variación en la radiación solar incidente es un agente forzante del cambio climático, este factor deberá ser considerado al analizar la producción de fitoquímicos en los cultivos.

