











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Jatropha curcas L. (Euphorbiaceae), es originaria de México y Centroamérica, utilizada principalmente para obtener biodiesel mediante la transesterificación de los aceites extraídos de las semillas. Actualmente esta especie se encuentra distribuida en Centro y Sudamérica, África, India, Sureste de Asia y Australia (Carels, 2009). Por su alto contenido de aceite y proteína (40% y hasta 28%, respectivamente), está siendo estudiada ampliamente en el mundo (Francis et al., 2013). El piñón de cerro, como se le conoce comúnmente a J. curcas, es una especie que en México cuenta con diversos usos tradicionales, principalmente en la Huasteca y pueblos totonacos (Totonacapan); como tutor de la vainilla, para la elaboración de comidas típicas, para reforestación, cercas vivas, conservación de suelos, medicinal y su uso potencial industrial es como bioenergético (Vera et al., 2014).
Dentro del complejo de plagas que atacan al piñón de cerro, se encuentra Leptoglossus zonatus Dallas, (Heteróptera: Coreidae), conocido comúnmente como “chinche pata de hoja”. Este insecto es un generalista que se alimenta de un amplio rango de plantas entre las que destacan cultivos básicos como maíz, sorgo, frijol, tomate, cítricos y algunos frutales como guayaba y maracuyá (Xiao y Fadamiro, 2009). La chinche pata de hoja se ha encontrado alimentándose y reproduciéndose en el piñón de cerro. Las ninfas y adultos de L. zonatus que se alimentan de frutos de J. curcas, reducen el rendimiento en semillas hasta en 21% (Morales et al., 2011). No obstante, según Calyecac (Com. Pers.), este problema está incrementándose considerablemente en monocultivo.
Debido a que el piñón cuenta con potencial alimenticio, medicinal y a que puede asociarse con otras especies vegetales de importancia alimenticia para el hombre, la búsqueda de alternativas de control biológico para las plagas que inciden en su cultivo cobra importancia.
La utilización de hongos entomopatógenos para el control de plagas de cultivos agrícolas ha sido ampliamente estudiada y utilizada en el mundo, debido a que de manera natural en los agroecosistemas, existe una relación específica, en muchos casos, entre patógeno y hospedero (Meyling y Eigenberg, 2007). Los hongos entomopatógenos infectan individuos en todos los órdenes de insectos; principalmente en Hemiptera, Diptera, Coleoptera, Lepidoptera, Hymenoptera y Orthoptera (Kirkland et al., 2004). En Nicaragua, dos hongos entomopatógenos, Beauveria bassiana y Metarhizium anisopliae (Metsch.) Sorok., evidenciaron un control satisfactorio de L. zonatus en el piñón de cerro (Grimm y Guharay, 1998).
Debido a la importancia que tiene el cultivo de J. curcas como potencial de alimento humano y a que existen pocos trabajos encaminados al control biologico de plagas en esta, en el presente trabajo se evaluó el efecto de tres cepas mexicanas nativas de B. bassiana para el control de L . zonatus bajo condiciones de laboratorio.
Materiales y métodos
Colecta del insecto
Adultos de L. zonatus fueron capturados en una plantación experimental de J. curcas del Centro de Desarrollo de Productos Bióticos del Instituto Politécnico Nacional (CEPROBI), ubicado en Barranca Honda, Yautepec Morelos (18º 49’ latitud norte y 99º 05’ longitud oeste), a una elevación de 1 071 msnm con una precipitación media de 1 050 mm anuales y una temperatura promedio anual de 22 °C (García, 1987). También fueron obtenidos en parcelas de frijol y sorgo de la comunidad de San Isidro Yautepec, en las cercanías del CEPROBI y en parcelas de sorgo dentro de la cabecera municipal.
Después de que los adultos de L. zonatus fueron colectados, se establecieron en el laboratorio del Banco Nacional de Germoplasma Vegetal (BANGEV) ubicado en el Departamento de Fitotecnia de la Universidad Autónoma Chapingo (UACH) (Carretera México-Texcoco km 38.5, Chapingo, Estado de México).
Los insectos fueron aislados en jaulas de vidrio de 30 x 30 x 40 cm y sometidos a observación por cinco días para asegurar que estuvieran libres de patógenos y parasitoides. El ambiente dentro de las jaulas se mantuvo a una temperatura de 25 a 30 °C y humedad relativa de 65 a 78%. La humedad fue controlada con un humidificador eléctrico marca Duracraft. Le temperatura fue regulada mediante el calor emitido por focos de 100 watts. Los adultos de L. zonatus fueron alimentados con granos de maíz tierno, frutos frescos de piñón y vainas frescas de frijol, previamente lavados con agua destilada (Xiao y Fadamiro, 2009).
Características del hongo
Las cepas utilizadas en los bioensayos (Cuadro 1), fueron proporcionadas por el Comité Estatal de Sanidad Vegetal del estado de Guanajuato (CESAVEG) a través del M. C. Fernando Tamayo y reactivadas en la UACH, en medio de cultivo solidificado agar dextrosa saborau (ADS), y mantenidas en cajas Petri de plástico a 25 °C hasta su utilización en los bioensayos.
Cuadro 1 Procedencia de las cepas de B. bassiana utilizadas en los bioensayos.
| Cepa | Aislamiento | Lugar de colecta y coordenadas geográficas | Fecha de colecta | Fecha de reactivación |
| BB01 | Chapulín Melanoplus spp. | San Bartolo, Municipio de Apaseo el Alto, Guanajuato (20°30´47.77´´N, 100° 33’ 2.05’’ O) | 10/02/2004 | 14/04/2009 |
| BB02 | Chinche Lygus spp. En maíz | Irapuato (20°44´50.71´´N, 101°19’38.1’’O). | 08/08/2004 | 14/04/2009 |
| BB32 | Bactericera spp. en chile | Irapuato | 08/08/2004 | 14/04/2009 |
Evaluación de la patogenicidad de tres cepas de B. bassiana
Para la patogenicidad y virulencia se evaluaron las tres cepas y un testigo utilizando el diseño completamente al azar, cada tratamiento tuvo tres repeticiones y nueve insectos por repetición. Por cinco días, los adultos de L. zonatus se mantuvieron aislados. Al termino de la cuarentena, se sumergieron por tres segundos en una suspensión acuosa de Tween 80 al 0.03% con 1 x 108 conidias mL-1 para cada cepa. El testigo consistió solamente en la inmersión de los insectos en agua con Tween 80 a 0.03%. Inmediatamente después de la inmersión, los insectos fueron colocados en cajas Petri con papel filtro con la finalidad de eliminar el exceso de la suspensión (Grimm y Guharay, 1998). Posteriormente los insectos fueron colocados en recipientes de plástico de 500 cm3 de acuerdo a cada tratamiento y fueron alimentados con frutos frescos de J. curcas y vainas frescas de frijol. Durante el bioensayo, los insectos permanecieron en una incubadora (marca Seedburo modelo MP6-3000 B) a una temperatura de 24°C ± 1 con un rango de humedad relativa de 65 a 78% (Xiao y Fadamiro, 2009).
Evaluación de la mortalidad
La mortalidad de los insectos fue contada diariamente durante 50 días y fue ajustada mediante la fórmula de Abbott. Los insectos muertos fueron colocados de manera individual en cajas Petri con papel filtro esterilizado y humedecido con agua destilada e incubados por dos semanas a 25 °C para favorecer el crecimiento de micelio y producción de conidios.
Se determinó el porcentaje de esporulación y también la intensidad de la misma sumergiendo los insectos esporulados en una solución Tween 80 a 0.03%. Se agitó manualmente por 1 minuto y después se tomó una alícuota de 10µL y se diluyó en 1 mL de solución indicada al 0.03% y se procedió al conteo de conidios usando una cámara de Neubauer.
Germinación de conidias
El porcentaje de germinación de conidias se determinó tomando muestras de la misma suspensión utilizada en este bioensayo (1 x 108 conidios mL-1) y colocando alícuotas de 10 µL en cajas Petri con medio ADS solidificado e incubadas durante 24 h a 25±1 °C. Una vez transcurrido este periodo, se contabilizó el número de unidades germinadas de cada 300 conidias utilizando una cámara Neubauer. Este paso se realizó seis veces por cada cepa.
Efectividad biológica de la cepa BB01
Una vez que se determinó que la cepa BB01 fue la cepa más patogénica y virulenta de acuerdo a los resultados del ensayo anterior se establecieron seis tratamientos con tres repeticiones, ocho insectos por repetición. Los tratamientos consistieron en cinco concentraciones de conidios (1 x 104, 105, 106, 107, 108 conidios mL-1) suspendidas en una solución Tween 80 al 0.03% y el testigo el cual solo fue la solución Tween 80 al 0.03%.
Las concentración de conidias que provocan 50% de la mortalidad, dosis letal 50 (DL50), fue calculada tomando en cuenta el número de insectos vivos al inicio y al final de cada tratamiento, incluidas las repeticiones. El tiempo en el que murió 50% de la población, el tiempo letal 50 (TL50), fue evaluado para ambos ensayos calculado mediante el análisis Probit.
Análisis de datos
Los datos de las variables: 1) porcentaje de mortalidad; 2) porcentaje de esporulación; 3) intensidad de esporulación; y 4) germinación de esporas, fueron analizados bajo el diseño Completamente al Azar (DCA) con una prueba de análisis de varianza utilizando el programa estadístico SAS versión 9.0 y una prueba de comparación de medias utilizando la prueba de Tukey con un intervalo de confianza de α= 0.05 para el primer bioensayo y con la prueba de LSD y un α= 0.06 para el segundo bioensayo. La determinación y evaluación del porcentaje de mortalidad, porcentaje y concentración de esporulación, fueron realizadas de la misma forma que en el primer ensayo.
Resultados y discusión
Patogenicidad de tres cepas de B bassiana
La cepa BB01 aislada de Melanoplus spp. fue la cepa que presentó la mayor mortalidad (85.71%) y un mayor porcentaje de germinación de esporas (99.28%), en comparación con las otras dos cepas. En las variables porcentaje y concentración de esporulación, no hubo diferencias estadísticamente significativas (Cuadro 2).
Cuadro 2 Porcentaje de mortalidad y esporulación, concentración de esporulación y germinación de conidias de tres cepas de B. bassiana sobre adultos de L. zonatus.
| Cepa | Mortalidad (%) | Esporulación (%) | Concentración de la esporulación (1) | Germinación de conidias (%) |
| BB01 | 85.71 a* | 93.33 a | 18.8925 a | 99.28 a |
| BB02 | 42.86 ab | 66.67 a | 19.2370 a | 96.67 b |
| BB32 | 47.62 ab | 88.89 a | 18.5920 a | 95.40 b |
| TESTIGO | 22.22 b | 0.00 b | ----- | ------ |
(1) logaritmo natural (Ln) de la cantidad de conidas mL-1, *; letra diferente indica diferencia estadísticamente significativa (p= 0.05) en la prueba de Tukey.

Beavueria bassiana ha sido utilizada para regular poblaciones de diferentes coleópteros de los géneros Diabrotica, Colapis, y Maecolapis (Sosa et aI., 1994), además reduce hasta en 50 a 70% poblaciones de Ixodes scapularis y de Rhipicephalus sanguineus (Kirkland et al., 2004). La aplicación de B. bassiana en el suelo reduce la emergencia de adultos de Diabrotica undecimpunctata, y el daño a la raíz es menos severo en suelo tratado con el hongo que en los suelos infestados con la plaga sin tratamiento conidial (Krueger y Roberts, 1997). Grimm y Guharay (1998), mencionaron que también se puede utilizar como un agente de control biológico para L. zonatus y Pachycoris klugii, los cuales son de las dos principales plagas de J. curcas en Nicaragua, Guatemala y Brasíl.
Es posible que B. bassiana pueda ejercer un control efectivo en L zonatus debido al alto porcentaje de mortalidad obtenido en el presente trabajo, sin embargo, se debe tomar en cuenta que para que un organismo sea utilizado como agente de control biológico, no es suficiente un alto porcentaje de mortalidad sobre el organismo objetivo, sino también debe lograr la mortalidad con cierta rapidez, por lo que el tiempo letal 50, es una variable que cobra importancia. La cepa BB01 evaluada en este trabajo alcanzó TL50 a los 7.8 días, seguida por la BB02 y BB32 con 16.9 y 17.4 días respectivamente (Cuadro 3).
Cuadro 3 Tiempo letal 50 en días y significancia de tres cepas de B. bassiana sobre adultos de L. zonatus.
| Cepa | TL50 | Pr > ChiSq |
| BBO1 | 7.80679 | <.0001 |
| BB02 | 16.98266 | <.0001 |
| BB32 | 17.43740 | <.0001 |
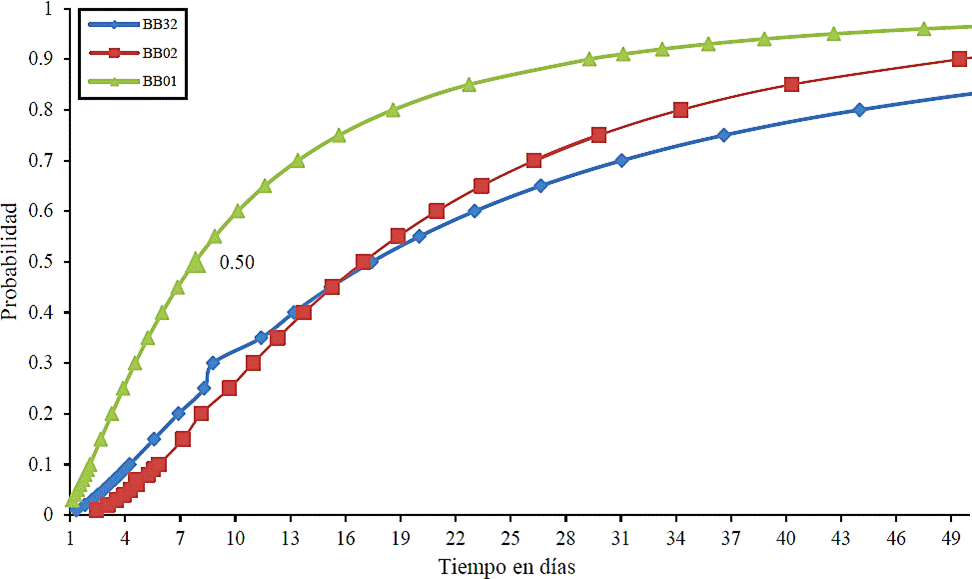
En relación a la rapidez del hongo en tener un efecto sobre sus hospedero, Lezama et al. (1998), mencionan que algunas cepas de B. bassiana presentan virulencia variable sobre Spodoptera frugiperda, con un parasitismo de hasta 90% en huevos y 100% en larvas, el aislamiento más sobresaliente presentó una TL50 de 3.1 y 2.8 días en huevos y larvas del mismo insecto, y una DL50 de 2.4 x 103 conidias mL-1. Por otra parte, en bioensayos realizados sobre el picudo del plátano Cosmopolites sordius, B. bassiana causa mortalidades de 6397% en los 35 días posteriores a la exposición de las esporas. (Kaaya et al., 1993). Dado lo anterior, es posible notar que la mortalidad y rapidez con que actúa B. bassiana, depende a) el organismo inoculado, b) la concentración de la aplicación del hongo, c) el estado de desarrollo del organismo objetivo y e) las condiciones ambientales.
En bioensayos realizados sobre M. sanguinipes, las cepas de B. bassiana GK2016, “tipo nativo” (virulento), y GK2051, presentaron un TL50 de 5.8 y 7.8 días respectivamente, aunque fueron sólo dos días de diferencia en el valor de TL50, la cepa GK2051 requirió 17 días para matar al 90% de la población y nunca produjo 100% de mortalidad, mientras que la cepa “tipo nativo” GK2016, causó 00% de muerte en 8-10 días (Kosir et al., 1991). Grimm y Guharay (1998), reportan que al evaluar la mortalidad de adultos de L. zonatus inoculados con tres diferentes cepas de B. bassiana, se alcanzó 50% de la mortalidad entre los 10 y 13 días después de la inoculación. Es posible deducir que la mortalidad del organismo, no sólo depende de las características de éste, sino también depende de las características de la cepa utilizada.
Considerando los resultados indicados, la cepa BB01 de B. bassiana evaluada en este experimento, es más patogénica y virulenta sobre adultos de L. zonatus que las cepas del mismo hongo reportadas por Grimm y Guharay (1998), para el control de éste mismo insecto.
Efectividad biológica de la cepa BB01 de B. bassiana
Debido al mayor porcentaje de mortalidad (85.7%), menor tiempo en días para alcanzar el TL50 (7.8 días) y al mayor porcentaje de germinación de esporas (99.28%), la cepa BB01, fue utilizada para realizar el segundo bioensayo. La suspensión de 1 x 108 conidios mL-1 fue la que presentó los valores más altos en las variables de mortalidad (100%), esporulación (79.16%) y en la concentración de la esporulación (19.53), en comparación con los demás tratamientos (Cuadro 4).
Cuadro 4 Porcentaje de mortalidad y esporulación, concentración de esporulación de cepa BB01 de B. bassiana sobre adultos de L. zonatus.
| Tratamiento | Mortalidad (%) | Esporulación (%) | Concentración de la esporulación (1) |
| 1x104 | 75.12 ab* | 50.233 b | 18.4533 c |
| 1x105 | 70.15 b | 62.667 ab | 19.4600 ab |
| 1x106 | 70.14 b | 53.373 b | 18.7333 bc |
| 1x107 | 80.11 ab | 68.053 ab | 19.1300 abc |
| 1x108 | 100.0 a | 79.167 a | 19.5333 a |
| Testigo | 16.67 c | 0.000 c | 0.0000 d |
(1) logaritmo natural (Ln) de la cantidad de conidas mL-1, *; diferente letra indica diferencias significativas entre tratamientos bajo la prueba LSD (p== 0.06).
Al respecto, la mortalidad de larvas y adultos de la catarina de la papa Leptinotarsa decemlineata está relacionada con aplicaciones en diferentes dosis (1 x 104, 3 x 104 y 1 x 105 conidias cm2 -1 de área foliar) de B. bassiana (Fargues et al., 1994). Por otra parte, Grimm y Guharay (1998), al estudiar el efecto de diferentes cepas de B. bassiana (1 x 107 conidias mL-1), sobre adultos de L. zonatus, encontraron mortalidades de 88 a 99%. Las diferentes dosis de la cepa BB01 de B. bassiana (Cuadro 4), evaluadas en el presente trabajo, presentaron mortalidades desde 70% hasta 100%, este último valor obtenido en la concentración de 1 x 108 conidias mL-1, por lo que la mortalidad de L. zonatus también está relacionada con las dosis aplicadas del hongo.
La susceptibilidad y la relación de los hongos con los hospederos se asocian con los nutrimentos presentes sobre el cuerpo de los insectos, los cuales a su vez son el medio para la propagación, dispersión y persistencia de dichos organismos (Meyling y Eigenberg, 2007). Debido a lo anterior, la esporulación del hongo en el cuerpo del insecto es un medio de dispersión y propagación tal y como se observa en nidos de la hormiga de fuego, Solenopsis invicta, donde B. bassiana crece y esporula (Pereira et al., 1993), por lo que representa un agente potencial de control biológico para dicha hormiga (Siebenelcher et al., 1992). En este experimento, la dosis de 1 x 108 conidias mL-1 tuvo la mayor esporulación (79.167%), (Cuadro 4), por lo que la capacidad de dispersión del hongo y de infección a otros insectos, es mayor.
La DL50 de la cepa BB01 sobre adultos de L. zonatus fue de 5.2 x 107 conidias mL-1 (Pr>ChiSq= 0.0021) (Cuadro 5), por lo que la DL50 se encuentra dentro del rango de las concentraciones evaluadas en el presente trabajo.
Cuadro 5 Dosis letal 50, en conidias mL-1 de la cepa BB01 de B. bassiana en adultos de L. zonatus.
| Tiempo | DL50* | Pr > ChiSq |
| 8 Días | 52903196 | 0.0021 |
*Conidias por mililitro.
Al respecto dosis de B. bassiana cercanas a la DL50 redujeron el potencial reproductivo de S. lineatus, la fertilidad y fecundidad de C. suppressalis así como, el desarrollo de los huevecillos de las chicharritas en plantas de arroz (Feng et al., 1994). Por otra parte aislamientos de B. bassiana sobre larvas y adultos de S. frugiperda, presentaron una DL50 de 2.4 x 103 conidias mL-1 y una TL50 de 3.8 días (Lezama et al., 1996).
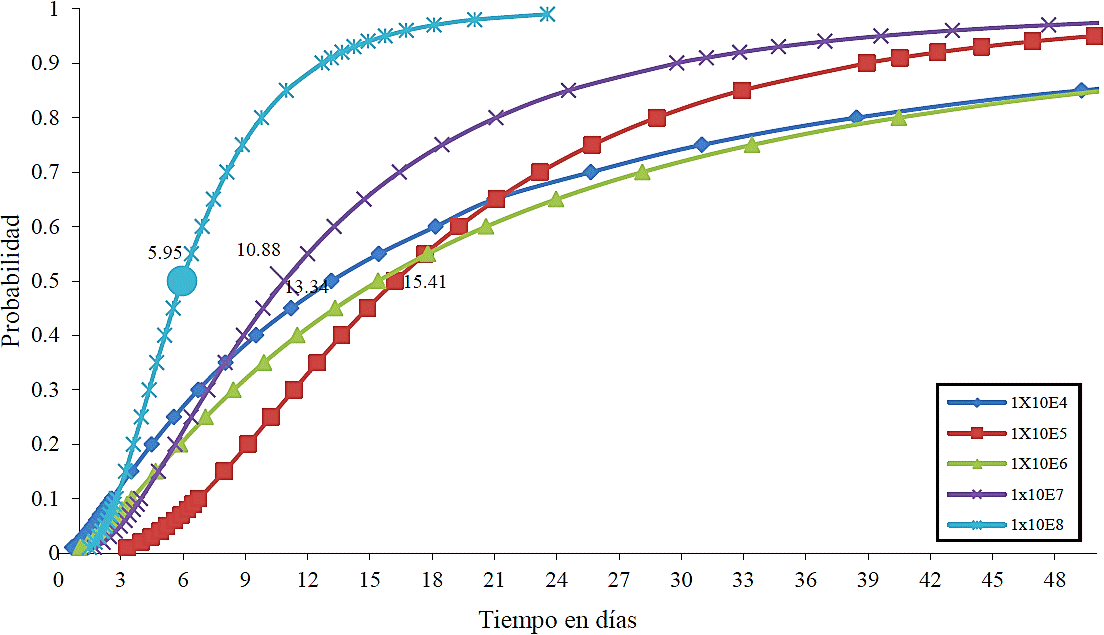
La dosis de 1 x 108 conidias mL-1 fue la que presentó el menor valor en el TL50 (5.9 días), por lo que es la dosis que mata con mayor rapidez al 50% de la población de L. zonatus (Cuadro 6).
Cuadro 6 Tiempo letal 50 en días de la cepa BB01 de B. bassiana en adultos de L zonatus.
| Cepa | TL50* | Pr > ChiSq |
| 1X104 | 13.15832 | <.0001 |
| 1X105 | 16.22547 | <.0001 |
| 1X106 | 15.41490 | <.0001 |
| 1X107 | 10.88828 | <.0001 |
| 1X108 | 5.95965 | <.0001 |
*Tiempo letal 50 en días.
La dosis letal 50 de Diatrea sacharalis está relacionada con la dosis ingerida de esporas, dadas las primeras muertes atribuidas a B. Bassiana, las cuales ocurrieron a los 7.7 días en dosis de y 1 x 108 conidias mL-1, mientras que para Tetranychus urtice con una dosis de1 x 108 conidias mL-1, la dosis letal 50 se presentó a los 3.6 días después de la inoculación (Batista et al., 2004). Por otra parte, B. bassiana es un organismo potencial para ser usado en el control biológico de áfidos. En un bioensayo realizado con dosis de 1 x 104 a 1 x 108 conidias mL-1, el hongo presentó un alto nivel de virulencia sobre el pulgón Phorodon humuli, obteniéndose una DL50 de 1.37 x 105 conidias mL-1 y el TL50 se redujo en la medida del incremento de la dosis conidial (Dorschner et al., 1991). Algo similar ocurrió en el presente trabajo debido a que las dosis más altas presentaron los valores más bajos en el TL50 (Cuadro 6).
Considerando lo citado, B. bassiana es un hongo que puede ser utilizado para el control biológico de poblaciones de L. zonatus de una forma inocua, ya que no genera contaminación al ambiente, sin embargo, existe el riesgo potencial de dañar algunos organismos benéficos. Debido a lo anterior, habrá que realizar los estudios correspondientes en condiciones de invernadero y de campo para validar la efectividad de este hongo contra L. zonatus en las condiciones diversas.
Conclusiones
El efecto de las tres cepas de Beauveria bassiana fue patogénico y virulento sobre adultos de L. zonatus.
Las cepas evaluadas evidenciaron diferente nivel de patogenicidad y virulencia sobre adultos de L. zonatus.
La cepa BB01 evidenció mayor potencial como agente de control biológico de L. zonatus.
Existe una relación directamente proporcional entre la patogenicidad y virulencia de cepas de B. bassiana (BB01) con la dosis aplicada sobre adultos de L. zonatus.

