











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Durante décadas la extracción de productos de los bosques, tanto maderables como no maderables, ha sustentado y mantenido la economía de comunidades humanas de México. Los productos forestales no maderables (PFNM) constituyen recursos biológicos que provienen de áreas forestales naturales, áreas forestales bajo manejo y de otras áreas boscosas (Peters, 1996); incluyen una variedad de frutas, nueces, semillas, aceites, especias, resinas, gomas, plantas medicinales y otros productos (de Beer y McDermott, 1989). Éstos, cumplen un rol vital ya que la mayoría son de autoconsumo y forman parte del sistema económico productivo informal (Van Rijsoort, 2000).
En el noreste de México se localiza 7.1% de la población total de México, constituida en los estados de Tamaulipas, Nuevo León y Coahuila, siendo estos estados eminentemente urbanos. Sin embargo, 15% de sus habitantes se dedica a la explotación y extracción de recursos vegetales de sus diferentes tipos de vegetación, como es el orégano (Lippia graveolens Kunth), la damiana (Turnera diffusa Willd.), el chile piquín (Capsicum annum var. aviculare [Dierb.] D´Arcy y Eashbaugh), hojas de palma (Sabal mexicana Mart., Chamaedorea radicalis Mart., Brahea dulcis [Kunth] Mart.), vainas de mezquite (Prosopis juliflora [Sw.] DC.), ébano (Ebenopsis ebano Berland.), laurel (Litsea glaucescens Kunth). (Medina et al., 1999). Dentro de las plantas silvestres de Tamaulipas, Hernández et al. (1991) enlistan 610 especies útiles, de las cuales 30% son comestibles.
Sin embargo, procesos ecológicos como la herbívora causada por insectos reducen la calidad, cantidad y precio de los productos vegetales (hojas, tallos, raíces, flores, frutos y semillas) que con frecuencia son dañados total o parcialmente (Crawley, 1983). Entre los herbívoros de la clase Hexapoda más importantes económicamente, se encuentran los crisomélidos, quienes además son vectores de bacterias y virus fitopatógenos. Los escarabajos de la familia Chrysomelidae comprenden 37 000 especies descritas alrededor del mundo que están agrupadas en 13 subfamilias y más de 2 000 géneros, éstos son fitófagos y se encuentran entre las familias de insectos más diversos y conspicuos asociados a plantas (Bouchard et al., 2011).
Los individuos de Chrysomelidae son conocidos como “tortuguitas, doradillas, pulgas saltonas, chochos, diabróticas o, terroncitos”, y se caracterizan por tener su cuerpo ovalado o semioval, ligeramente cilíndrico o alargado, muy convexo generalmente brillante y con colores contrastantes o metálicos y la cabeza pequeña en su mayoría hipognata. Las antenas son largas filiformes o capitadas, usualmente con once segmentos e insertas entre los ojos y la base de las mandíbulas. Las patas son cortas y delgadas a excepción de la tribu Alticini, su fórmula tarsal seudotetrámera, debido a la reducción del cuarto artejo; el tercer artejo es bilobulado y pubescente por debajo. Su longitud es variable desde los dos hasta los 20 mm (Morón y Terrón, 1988).
La mayoría de las especies son específicas o tienen preferencias de plantas como alimento, hay monófagas o más comúnmente oligófagas. El consumo de algunas está estrechamente relacionado con las plantas (Pérez-Contreras, 1999). En muchos casos, los crisomélidos se alimentan de un grupo de plantas afines, y estos patrones son frecuentemente evidentes en todo el mundo.
Algunas especies de la subfamilia Cassidinae son conocidas como tortuguitas o escarabajos tortugas (Jolivet et al., 1988). La mayoría de los Cassidinae son consumidores especializados (oligófagos). Las especies de Cassidinae están asociadas principalmente a plantas hospederas de las familias Convolvulaceae, Asteraceae, Bignoniaceae, Boraginaceae, Lamiaceae y Solanaceae (Jolivet y Hawkeswood, 1995). Al igual que muchos miembros de la subfamilia Cassidinae, Plagiometriona clavata (Fabricius) parece preferir las plantas de la familia Solanaceae, incluyendo el género Capsicum (Jolivet y Hawkeswood, 1994) (Figura 1).
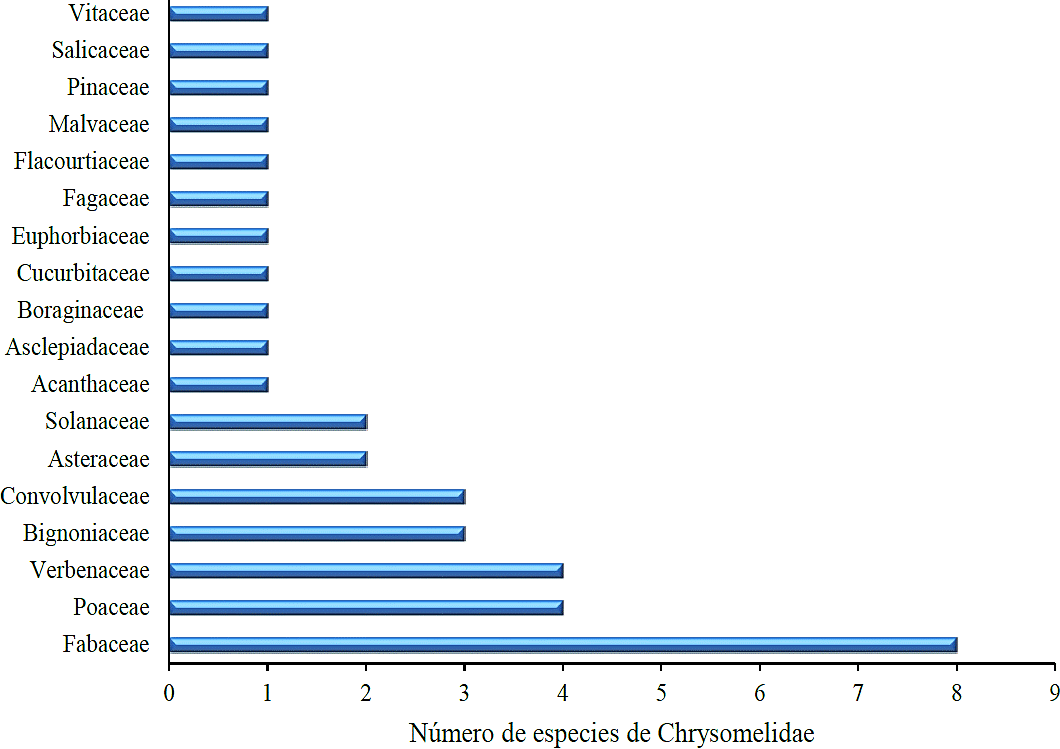
Figura 1 Número de especies de crisomélidos asociados a familias de plantas hospederas en el Cañón La Libertad y el Cañón del Novillo, Victoria, Tamaulipas.
La biología y dinámica de las poblaciones de crisomélidos son escasamente conocidas. Su estudio se ha centrado en pocas especies, principalmente plagas de cultivos agrícolas como la catarina de la papa (Leptinotarsa decemlineata [Say]), las doradillas (Diabrotica spp.), el arrocillo (Colaspis spp.) o la pulga saltona (Epitrix cucumeris [Harris]). Especies de estos géneros son univoltinas o multivoltinas, y el número de generaciones al año depende de las condiciones ambientales y la disponibilidad de alimento (García-Leaños y Marín-Jarillo, 2010; Pérez-Domínguez et al., 2010).
El término plaga tiene un sentido marcadamente antropocéntrico, puesto que el hombre lo aplica a todo aquello que le produce algún daño. Sin embargo, la noción de plaga se asocia casi exclusivamente a los insectos y otros artrópodos (Selfa y Anento, 1997).
Existe una serie de factores que regulan las poblaciones de los insectos y estos se pueden ver en dos variables. Uno es el potencial biótico, es decir la habilidad que tiene una especie para multiplicarse sin que exista una fuerza contraria que lo impida. Otro, la resistencia al medio (factores bióticos y abióticos) es precisamente la relación que existe entre ambos factores que nos indica la abundancia poblacional de una especie determinada.
Para responder a preguntas sobre posibles plagas potenciales de importancia económica agrícola o forestal, se consultó en la literatura especializada de los posibles huéspedes que causan problemas a las especies de plantas de importancia económica que se registraron en la zona de estudio.
El objetivo de este trabajo fue identificar las especies de Chrysomelidae asociados a los recursos forestales maderables y no maderables en los Cañones del Novillo y la Libertad, Victoria, Tamaulipas, México.
Materiales y métodos
El área de estudio se encuentra en la parte central del estado de Tamaulipas: el Cañón de “La Libertad”, ubicado entre los 23° 46' 34" latitud norte y los 99° 13' 21" longitud oeste, y el Cañón del Novillo ubicado entre los 23° 42' 19" latitud norte y los 99°13'31" longitud oeste, del municipio de Victoria. En la zona se presenta un clima semicálido subhúmedo con lluvias en verano. La temperatura media anual es de 24 °C y la precipitación anual fluctúa de 717.3 a 1,058.8 mm. Para el trabajo de campo se realizaron 20 cuadrantes de 20 x 10 m (200 m²), en el Cañón de La Libertad se muestrearon cinco cuadrantes para el bosque de pino-encino con una elevación entre 540 y 560 msnm y cinco cuadrantes para la selva baja subcaducifolia que oscila entre 380 y 405 msnm, y en el Cañón Del Novillo cinco cuadrantes para el bosque de pino-encino con una elevación entre 710 y 580 msnm y cinco cuadrantes para la selva baja subcaducifolia entre 510 y 520 msnm, durante un año de colecta, comprendiendo de febrero 2012 a enero 2013, preferentemente de las 9:00 a 17:00 h.
La colecta de los especímenes se obtuvo principalmente sobre la vegetación herbácea y arbustiva de acuerdo con los hábitos fitófagos de los Chrysomelidae (Jolivet, 1988), y se reforzó la colecta con 50 golpes de red por cuadrante. Se utilizaron las técnicas y herramientas habituales en entomología: red de golpeo, sábana o paraguas invertido y colecta directa. Los hospederos de los escarabajos adultos fueron registrados en el campo, y comprenden las plantas sobre las que se observó a los insectos alimentándose, y se tomaron fotografías para complementar lo observado. En la Figura 2a y b, se muestra a P. clavata en estado adulto y una larva alimentándose de hojas de chile piquín, estas observaciones cayeron dentro de un cuadrante ubicado en la vegetación de selva baja subcaducifolia a una elevación de 390 msnm, en el Cañón de La Libertad.

Figura 2 a) Adulto de Plagiometriona clavata, b) Larva de P. clavata alimentándose de hojas de chile piquín. (Foto: I. Martínez-Sánchez).
La investigación se llevó a cabo en dos tipos de vegetación, selva baja subcaducifolia y bosque de pino-encino preferentemente en zonas no perturbadas alejadas de las actividades antropogénicas, puede que las especies de importancia agrícola o económica se encuentren presentes pero en un número bajo.
Trabajo de laboratorio
En laboratorio se separaron las muestras realizando los siguientes pasos: vaciado del material biológico, separación, preservación, montaje, etiquetado, revisión, ordenamiento, determinación taxonómica.
Determinación taxonómica del material
Para la identificación de las plantas hospederas se tomaron partes de éstas y se colocaron en una prensa botánica para transportarlas hasta el “Herbario de Tamaulipas” donde se llevó a cabo su determinación taxonómica a género y especie mediante bibliografía especializada (Miranda y Hernández, 1963; Puig, 1976; Rzedowski, 1978 y García, 2009), y el apoyo de especialistas Arturo Mora Olivo y Jacinto Treviño Carreón. Para la determinación de Chrysomelidae, se utilizó el criterio de clasificación taxonómica reconocido para Norteamérica basado en Riley et al. (2002). Para la identificación a distintos niveles taxonómicos se utilizaron las siguientes claves: a nivel familia y subfamilia se utilizaron las claves de Triplehorn y Johnson (2005), para géneros y especies las claves de Monrós (1959), Wilcox (1965), Scherer (1983), Noguera (1988), Domínguez (1990), White (1993), Flowers (1996 y 2004) y Chamorro-Lacayo y Konstantinov (2009).
Resultados y discusión
Se obtuvo un total de 119 ejemplares, 81 ejemplares para el Cañón de la Libertad y 38 para el Cañón del Novillo, se registra un total de 26 especies pertenecientes a cinco subfamilias y 24 géneros de Chrysomelidae (Cuadro 1) asociados a 18 familias de plantas hospederas (Cuadro 2). La subfamilia Cassidinae registró la mayor diversidad específica con 12 especies, seguido de Galerucinae con seis y en menor número le siguen Chrysomelinae y Cryptocephalinae con tres especies cada una, en último lugar a Eumolpinae representado por dos especies (Cuadro 1). De acuerdo con el tipo de vegetación se registran 16 especies para la selva baja caducifolia y 15 para el bosque de pino-encino del Cañón de La Libertad, en tanto el Cañón del Novillo tiene seis especies para la selva baja subcaducifolia y 11 para el bosque de pino encino, cabe hacer mención que de las 26 especies registradas en la zona de estudio, ocho especies son exclusivas para la selva baja subcaducifolia, nueve especies son exclusivas del bosque pino-encino de ambos cañones y nueve especies de crisomélidos compartieron los dos tipos de vegetación. El mayor número de especies se ve reflejada en la subfamilia Cassidinae con el 46.15% del total de la colecta y Galerucinae con 23.07%, en otro trabajo similar de Sánchez-Reyes et al. 2015 registraron para la zona del Cañón de la Peregrina 157 especies, de la cual Galerucinae obtuvo 49% del total de las especies colectadas, mientras que Cassidinae con 20%, totalmente inverso a nuestro trabajo.
Cuadro 1 Listado taxonómico de Chrysomelidae en el Cañón del Novillo y el Cañón de La Libertad, Tamaulipas, México.
| Nombre científico | Nombre científico |
| Cassidinae (Gyllenhal, 1813) | Cryptocephalinae (Gyllenhal, 1813) |
| Tribu Chalepini (Weise, 1910) | Tribu Cryptocephalini (Gyllenhal, 1813) |
| Brachycoryna pumila (Guérin-Méneville, 1944) | Subtribu Cryptocephalina (Gyllenhal, 1813) |
| Chalepus amabilis (Baly, 1855) | Cryptocephalus umbonatus (Schaeffer, 1906) |
| Chalepus digressus (Baly, 1885) | Tribu Clytrini (Lacordaire, 1848) |
| Heterispa vinula (Erichson, 1847) | Subtribu Clytrina (Lacordaire, 1848) |
| Microctenochira punicea (Boheman, 1855) | Anomonea rufifrons (Lacordaire, 1848) |
| Sumitrosis rosae (Weber, 1801) | Subtribu Babiina (Chapuis, 1874) |
| Xenochapelus omogerus (Crotch, 1873) | Babia tetraspilota (LeConte, 1858) |
| Tribu Cassidini (Gyllenhal, 1813) | Eumolpinae (Hope, 1840) |
| Charidotella sexpunctata (Fabricius, 1781) | Tribu Adoxini (Baly, 1863) |
| Charidotis auroguttata (Boheman, 1855) | Fidia albovittata (Lefèvre, 1877) |
| Coptocycla texana (Schaeffer, 1933) | Xantonia sp. (Baly, 1863) |
| Plagiometriona clavata (Boheman, 1855) | Galerucinae (Latreille, 1802) |
| Tribu Mesomphaliini (Hope, 1840) | Tribu Alticini (Newman, 1835) |
| Chelymorpha pubensces (Boheman, 1854) | Alagoasa bipunctata (Chevrolat, 1834) |
| Chrysomelinae (Latreille, 1802) | Centralaphthona fulvipennis (Jacoby, 1885) |
| Tribu Chrysomelini (Latreille, 1802) | Systena contigua (Jacoby, 1889) |
| Subtribu Doryphorina (Motschulsky, 1860) | Tribu Luperini (Chapuis, 1875) |
| Labidomera suturella (Chevrolat, 1838) | Subtribu Diabroticina (Chapuis, 1875) |
| Subtribu Chrysomelina (Latreille, 1802) | Acalyma vittata (Fabricius, 1975) |
| Plagiodera semivittata (Stål, 1860) | Gynandobrotica lepida (Say, 1835) |
| Plagiodera thymaloides (Stål, 1860) | Neobrotica sexmaculata (Jacoby, 1887) |
Cuadro 2 Lista de Chrysomelidae asociados a familias forestales maderables y no maderables.
| Especie | Familia hospedera | Planta hospedera | Importancia y uso de la planta |
| Especies encontrados en selva baja subcaducifolia del Cañón de La Libertad | |||
| Acalymma vittatum Fabricius (1775) | Cucurbitaceae | Cucurbita sp., y Cucumis sp. y Sechium sp. | Agrícola |
| Anomoea rufifrons Lacordaire (1848) | Fabaceae | Acacia farnesiana, Prosopis laevigata | Maderable |
| Charidotis auroguttata Boheman (1855) | Bignoniaceae | Macfadyena unguis-cati | Ornamental y medicinal |
| Chelymorphapubescens Boheman (1854) | Convolvulaceae | Jacquemontia pentantha | Medicinal y melífera |
| Microctenochirapunicea Boheman (1855) | Convolvulaceae | Melífera | |
| Plagiodera thymaloides Stål (1860) | Flacourtiaceae | Neopringlea integrifolia | Melífera |
| Especies encontradas en bosque de pino-encino del Cañón de La Libertad | |||
| Chalepus digressus Baly (1885) | Poaceae | Panicum sp., y Zea mays | Agrícola y forrajera |
| Cryptocephalus umbonatus Schaeffer (1906) | Asteraceae | Conyza sp. | Medicinal |
| Pinaceae | Pinus pseudostrobus | Maderable | |
| Fidia albovittata Lefévre (1877) | Vitaceae | Vitis sp. y Vitis tiliifolia | Medicinal y Melífera |
| Especies compartiendo selva baja subcaducifolia y bosque pino-encino del Cañón de | |||
| Sumitrosis rosea Weber (1801) | Fabaceae | Desmodium sp. | Medicinal |
| Chalepus amabilis Baly (1855) | Poaceae | Panicum sp. y Zea mays | Ganadera |
| Especies encontradas en bosque pino-encino del Cañón Del Novillo | |||
| Babia tetraspilota LeConte (1858) | Fabaceae | Prosopis laevigata, Acacia farnesiana | Maderable y comestible |
| Desmodium sp. | Medicinal | ||
| Especies encontrados en selva baja subcaducifolia del Cañón Del Novillo | |||
| Coptocycla texana Schaeffer (1933) | Boraginaceae | Ehretia anacua | Ornamental |
| Especies encontradas en ambos Cañones en selvas bajas subcaducifolias | |||
| Charidotella sexpunctata Fabricius (1781) | Convolvulaceae | Convolvulus arvensis | Maleza |
| Bignoniaceae | Macfadyena unguis-cati | Ornamental y medicinal | |
| Convolvulaceae | Jacquemontia pentantha | Medicinal y melífera | |
| Labidomera suturella Guérin-Méneville (1838) | Asclepiadaceae | Asclepias curassavica | Medicinal |
| Plagiometriona clavata Boheman (1855) | Solanaceae | Capsicum annuum var. aviculare | Agrícola |
| Especies encontradas en ambos Cañones en bosques de pino-encino | |||
| Neobrotica sexmaculata Jacoby (1887) | Verbenaceae | Lantana sp. | Medicinal |
| Bignoniaceae | Chilopsis linearis | Agroforestal y ornamental, | |
| Plagiodera semivittata Stál (1860) | Salicaceae | Salix bomplandiana, Populus mexicanus | Reforestación, Restauración y comercial |
| Systena contigua Jacoby (1889) | Fabaceae | Prosopis juliflora | Forestal |
| Solanaceae | Solanum sp. Lycopersicon sp. Capsicum sp. | Agrícola | |
| Verbenaceae | Lippia graveolens. | Agrícola | |
| Xenochapelus omoger Crotch (1873) | Fabaceae | Centrosema sagittatum, Desmodium sp. Ebenopsis ebano, Leucaena pulverulenta, Bauhinia divaricata, Erythrina anacua. | Ornamental, medicinal, forestal, industrial y Medicinal |
| Especies compartiendo selva baja subcaducifolia y bosque pino-encino en ambos cañones | |||
| Alagoasa bipunctata Chevrolat (1834) | Acanthaceae | Yeatesia mabryi | Melífera, agrícola, química y medicinal |
| Verbenaceae | Lippia graveolens | ||
| Fabaceae | Desmodium sp. | ||
| Gynandobrotica lepida Say (1835) | Fabaceae | Phaseolus vulgaris | Agrícola |
| Heterispa vinula Erichson (1847) | Poaceae | Panicum sp. | Ganadera |
| Malvaceae | Pavonia uniflora, Sida acuta | Farmacológica Medicinal | |
| Brachycoryna pumila Guérin-Méneville (1944) | Asteraceae | Aphanostephus ramosissimus var. humilis, Baccharis salicifolia | Artesanal y medicinal |
| Verbenaceae | Phyla incisa | Ornamental | |
| Centralaphthona fulvipennis Jacoby (1885) | Euphorbiaceae | Croton ciliato- glandulifer, Euphorbia graminea. | Química, medicinal y forraje |
| Fabaceae | Desmodium sp., Bauhinia divaricata | Medicinal, ornamental y melífera | |
| Poaceae | Panicum sp., y Zea mays | Ganadero y agrícola | |
| Xantonia sp. | Fagaceae | Quercus polymorpha | Forestal |
La diferencia de resultados en especies encontradas en estos trabajos puede deberse a factores ambientales como las condiciones climáticas favorables en el momento de la colecta en determinado año y al trabajo físico, número de redazos sobre la vegetación arbustiva, 200 golpes de red para Sánchez-Reyes sobre 50 para este trabajo donde en su mayoría de colectas se usó la técnica de colecta directa (manual) sobre las plantas. En otro trabajo en la Sierra de San Carlos dentro del mismo estado de Tamaulipas, Sánchez-Reyes et al. 2014, capturaron un total de 290 ejemplares de Chrysomelidae, nuevamente ellos observaron que la mayor riqueza total corresponde a la subfamilia Galerucinae (43.24%), de nuevo existe diferencia con la diversidad de especies para este trabajo, con respecto a la vegetación en este trabajo se registra una mayor diversidad de especies (39) en la comunidad vegetal bosque de encino, y (5) especies para la selva. En comparación a nuestro trabajo el bosque de pino encino registro (18) especies y (17) especies de crisomélidos asociados con selva baja subcaducifolia.
La familia Fabaceae registra la mayor cantidad de insectos asociados con ocho especies de crisomélidos, contrario al trabajo de Gavrilović y Ćurčić (2013) quienes registraron que Asteraceae fue una de las familias mejor representadas con especies de plantas hospederas con 11 especies de crisomélidos en la Provincia de Vojvodina, Serbia; por otro lado Aslan y Gök (2006) reportan que Asteraceae fue de las cinco familias con mayor número de hospederos de 65 especies de crisomélidos registrados en Turquía. Aslan et al. (2009), encontraron en Antalya, Turquía que la familia Asteraceae dominó en 5% de plantas hospederas para crisomélidos del estrato arbustivo, muy similar a lo encontrado en este trabajo con 5.4%.
Las familias Poaceae y Verbenaceae presentaron un número menor de crisomélidos, en este caso se registraron cuatro especies asociadas; en tanto para las familias Bignoniaceae y Convolvulaceae se encontraron tres especies. Asteraceae y Solanaceae presentan dos especies. Finalmente con una sola especie se registró a las familias Acanthaceae, Asclepidaceae, Boraginaceae, Cucurbitaceae, Euphorbiaceae, Fagaceae, Flacourtiaceae, Malvaceae, Pinaceae, Salicaceae y Vitaceae (Figura1).
La subfamilia Cassidinae registró la mayor cantidad de familias (8) de plantas hospederas siendo está también la de mayor diversidad en especies dentro del área de estudio, en donde Poaceae, Fabaceae, Convolvulaceae y Bignoniaceae presentaron la mayor frecuencia de asociación de especies de casidinos; tal como lo menciona Jolivet (1988), estas familias vegetales son las de mayor número de hospederos para crisomélidos adultos. En el Cuadro 2 se da un listado de especies de Chrysomelidae asociados a especies vegetales maderables y no maderables en los dos tipos de vegetación; asimismo las especies colectadas por cada cañón y especies compartidas por ambos cañones.
Todas las especies fueron colectadas en zonas conservadas, pero no se elimina la posibilidad que cualquiera de ellas pueda ser una plaga potencial si hubiese un cultivo cercano, puesto que tiene la capacidad de volar en busca del alimento.
De las especies de Chrysomelidae registradas, Plagiometriona clavata resultó ser la de mayor importancia agrícola para el chile piquín de la región (Capsicum annuum var. aviculare Dierb) (Figura 2). De acuerdo a la herbivoría causada por larvas y adultos encontrados sobre la planta, este podría ser un hospedero original para el ciclo de vida del escarabajo, esta especie puede llegar a ser una plaga de importancia para las poblaciones silvestres de chile piquín, como lo reporta Tejas et al. (2011) para Baja California Sur.
Es importante indicar que las densidades de crisomélidos fueron influidas por el clima prevaleciente en el periodo de colecta, ya que el año 2012 fue un año atípico con respectos a años anteriores; la temperatura media de la zona de estudio fluctúa entre 18 a 24.3 °C y la media de precipitación anual es de 717.3 mm a 1 058.8 mm con un clima Semicálido Subhúmedo (A)C(w0) con lluvias en verano (AlmaguerSierra 2005), pero en estos cañones de acuerdo a sus relieve y vegetación tienen un microclima propio de hasta 2 °C menos con respecto a la temperatura fuera de los cañones, esto hace que cualquier aumento de temperatura por arriba del promedio producirá desestabilidad, estrés hídrico y en ocasiones la muerte a especies animales y vegetales que son sensibles a cambios significativos de temperatura.
En este año (2012) las temperaturas altas de hasta 40 °C en los meses de abril y mayo en el Cañón de La Libertad y de 44 y 42 °C para mayo y junio, respectivamente en el Cañón del Novillo. Otro factor ambiental que hizo efecto en la baja densidad poblacional no solo de crisomélidos sino de otros artrópodos fue la falta de lluvias durante casi todas las estaciones del año que a su vez retardaron el crecimiento y desarrollo de la vegetación en la temporada de lluvias, la precipitación promedio anual (2012) en el Cañón del Novillo fue de 104 mm y 660 mm para el Cañón de La Libertad, ni siquiera alcanzo los 717.3 mm que es la precipitación anual más baja de la zona.
Conclusiones
En el Cañón de la Libertad y el Cañón del Novillo, Tamaulipas se registraron 24 géneros y 26 especies de Chrysomelidae, siendo la subfamilia Galerucinae la más abundante en individuos, estos datos concuerdan con los trabajos de Sánchez-Reyes et al. (2014; 2015); Niño (2005) realizados en Tamaulipas, donde la subfamilia Galerucinae fue la más abundante en la colecta total de crisomélidos.
Gynandobrotica lepida fue la más representativa con 17.6% (21) ejemplares abundancia, seguida de Xanthonia sp. con 15.1% (18) ejemplares ambas especies encontradas en los dos cañones.
De las 26 especies encontradas en los dos cañones, (8) se encontraron exclusivamente en la vegetación de selva baja subcaducifolia, mientras para el bosque de pino-encino se registraron (9) especies, así también son (9) las especies que comparten ambas comunidades vegetales. Puede que estas especies debido a su capacidad de volar y desplazarse a grandes distancia encontremos a especies en diferentes tipos de vegetación donde no las habíamos registrado antes, sobre todo las especies polífagas.
Como ya se ha dicho, uno de los principales factores que influyen en la distribución de Chrysomelidae es la presencia de la planta hospedera, ya que su diversidad está determinada de manera directa por la diversidad de plantas presentes en una zona (Baselga y Jiménez-Valverde, 2007).
La familia Fabaceae registró la mayor cantidad de crisomélidos asociados con ocho especies, siguiéndole en orden de importancia Poaceae y Verbenaceae (4); Bignonaceae y Convolvulaceae (3); Asteraceae y Solanaceae (2); y las familias con una sola especie Acanthaceae, Asclepiadaceae, Boraginaceae, Cucurbitaceae, Euphorbiaceae, Fagaceae, Flacourtiaceae, Malvaceae, Pinaceae, Salicaceae y Vitaceae. Alagoasa bipunctata, Gynandobrotica lepida, Heterispa vinula, Brachycoryna pumila, Centralaphthona fulvipennis y Xantonia sp. tienen un rango de distribución más amplio sobre los dos tipos de vegetación al tener disponibles plantas hospederas donde pueden completar su ciclo de vida.
Se registra por primera vez en Tamaulipas a Plagiometriona clavata asociada a chile piquín silvestre (Capsicum annuum var. aviculare).

