











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cultivo de chile (Capsicum annuum L.) es uno de los cultivos más importantes en México, por su gran consumo en la población (Namesny, 2006) en México, la superficie cosechada es de 143 975 hectáreas con un rendimiento promedio de 16.2 t ha-1 (SIAP, 2015).
El Ca2+ es un elemento alcalino-térreo (Feyerabend et al., 2008), con radio iónico de 9.9 nm (Hu et al, 2004), que tiene alta correlación con el suministro de La (lantano), de tal manera que con el suministro de lantano se eleva la acumulación de Ca2+ en hojas por planta (Ramírez et al., 2012). Es abundante en la mayoría de los suelos y rara vez se comporta como un factor limitante, salvo en suelos ácidos donde puede ser necesario el aporte de sales cálcicas (Bonilla, 2008), para complementar el requerimiento nutrimental del cultivo de chile, consistente en: N de 2.4-4 kgt-1 de frutos, fósforo (P2O5) de 0.4-1, potasio (K2O) de 3.4-5.3, calcio (CaO) de 0.6-1.8 y magnesio (MgO) de 0.3- 0.5, aunque este requerimiento varía en los tipos, órganos y sistemas de producción de chile jalapeño, dulce y pimiento (Salazar y Juárez, 2013). Según Charlo et al. (2012)), el pimiento puede extraer 81.3 kgha-1 y en él se pueden encontrar 0.84 kg t-1 de fruto; Azofeifa y Moreira (2004) reportan 38 kgha-1 y 0.82 kg t-1, repectivamente; Fontes et al. (2005) encontraron que el pimiento extrae 114 kgha-1 y contiene 2.2 kg t-1.
El ión Ca2+ se difunde hacia el interior de la planta a favor de un gradiente de concentración y se transporta hacia fuera con la ayuda de bombas dependientes de ATP (Salisbury y Ross, 2000), y los síntomas de su deficiencia siempre son más pronunciados en los tejidos jóvenes (Kirkby y Pilbeam, 1984), de tal manera que las zonas meristemáticas de las raíces, tallos y hojas, donde existen divisiones celulares, son las más susceptibles, quizás porque se necesita calcio para que forme una nueva laminilla media en la placa celular que aparece entre las células hijas (Salisbury y Ross, 2000).
En la célula ocurren secuencias de reacciones que requieren Mn2+, Ca2+ y Cl- ligados a un conjunto de polipéptidos, y otros datos experimentales sugieren que respuestas inducidas del fitocromo requieren intermediarios como el Ca2+ y calmodulina y que estas sustancias juegan un papel en la fosforilación de proteínas nucleares (Lea y Leegood, 1993). El Ca2+ es un elemento esencial para las plantas, toda vez que forma parte del Fotosistema II (FS II) que está integrado por seis polipéptidos integrales (intrínsicos) que se encuentran conectados entre sí de manera no covalente, en donde el Ca2+ es esencial para la fotólisis del agua, y hasta donde se incorpora con ayuda de tres polipéptidos extrínsecos (periféricos) que son codificados por genes del núcleo (Salisbury y Ross, 2000).
Actualmente se reconoce que todos los organismos mantienen concentraciones inesperadamente bajas de Ca2+ libre en el citosol, habitualmente menores de 1 μM (Hepler y Wayne, 1985). Esto resulta cierto incluso cuando el calcio es tan abundante en muchas plantas, sobre todo leguminosas, como el fósforo, azufre y magnesio. La mayor parte del calcio que contienen las plantas se encuentra en las vacuolas centrales, y en las paredes celulares se encuentra unido a ciertos polisacáridos llamados pectatos (Kinzel, 1989).
En las vacuolas, el calcio suele precipitarse en forma de cristales de oxalato insolubles. En algunas especies, también se encuentra en forma de carbonato, fosfato o sulfato insoluble. Las concentraciones bajas de calcio, casi micromolares, deben mantenerse en parte para impedir la formación de sales de calcio insolubles, obtenidas a partir de ATP y de otros fosfatos orgánicos. Además, las concentraciones de Ca2+ por encima del margen micromolar inhiben la corriente citoplasmática (Williamson, 1984). Aunque se activan unas enzimas mediante Ca2+, muchas otras quedan inhibidas, lo que hace todavía más necesario que las células mantengan concentraciones muy bajas de Ca2+ en el citosol, donde existen muchas enzimas (Salisbury y Ross, 2000). Una parte importante del Ca2+ existente en el citosol se une de forma directa a varias enzimas, como la pequeña proteína llamada calmodulina, con la que se une en forma reversible (Robert et al, 1986), haciendo que la citada proteína se modifique en su estructura y entonces active a varias enzimas (Salisbury y Ross, 2000).
El Ca2+ también actúa en el huso acromático durante la división celular, el cual se requiere para el normal funcionamiento de las membranas celulares, y ha sido implicado como mensajero secundario en respuesta a condiciones ambientales y señales hormonales (Sanders et al. 1999). La aplicación de Ca2+ en la producción de repollo es indispensable (Amador et al, 2008), porque este elemento asegura procesos, tales como la síntesis de paredes celulares, en la lámina media, donde forma pectato de calcio que confiere estabilidad y mantiene la integridad de éstas (Gordillo et al. 2004).
El calcio es poco móvil y tiende a acumularse en los órganos más viejos, mientras que los de mayor actividad metabólica (hojas en crecimiento, flores, frutos y meristemos apicales) son los tejidos que necesitan un mayor aporte; por tanto la deficiencia de este macronutriente afecta en primer lugar a las partes en formación y meristemos en crecimiento, donde queda fijado y prácticamente inmóvil en sus paredes celulares. Debido a esta inmovilidad, las hojas viejas pueden tener concentraciones normales de calcio, mientras que las hojas jóvenes, frutos u otros órganos, pueden presentar niveles por debajo de la normalidad (Chiu y Bould, 1977).
El proceso de transpiración y contenidos altos de agua en el suelo favorecen el flujo de masa y a su vez la movilidad de Ca2+ (Ansorena, 1994), de tal manera que con altas dosis de Ca2+ las cantidades de este elemento que no logran entrar al citosol quedan en la pared celular, situación que genera células más pesadas (Marschner, 2002).
El objetivo de esta investigación fue determinar la eficacia de las formulaciones de (CaNO3)2 líq, (CaNO3)2 sol-1, (CaNO3)2 sol-2 y (CaNO3)2 sol-3, en el índice de verdor de hojas y rendimiento por unidad de superficie, así como la dosis más adecuada de cada solución, para inducir la producción de más materia seca en plantas y frutos de pimiento morrón.
Materiales y métodos
La presente investigación se realizó durante el ciclo agrícola otoño-invierno 2014 a 2015 en un invernadero tipo capilla, instalado en el campo experimental de la Universidad Tecnológica de Culiacán, ubicado en el km 2 de la carretera Culiacán-Imala, colonia Los Ángeles, Culiacán, Sinaloa, con coordenadas 24° 50' 30" latitud norte y 107° 50' 30" longitud oeste, en altura de 58 msnm. Según García (1988), el clima es B1S1, semiárido con lluvias en verano e inverno y 670 mm de precipitación anual. La temperatura media anual es de 24 °C, con máximas de 41 °C en verano y mínimas de 5 °C en invierno, y humedad relativa promedio de 66.6% anual.
El cultivar de pimiento morrón (Capsicum annuum L.) utilizado fue 'Capia Rojo' Syngenta®; el sustrato consistió en una mezcla de peat moss, perlita, vermiculita y arena en proporciones volumen: volumen (v:v) de 1:1:1:1; antes de depositar el sustrato en las macetas hechas con bolsas de plástico con capacidad de 12 L, éstas fueron tratadas por imbibición en solución con Trichoderma harzianum (2x1012 ufc g-1), Bacillus subtilis (2x1012 ufc mL-1) y Bacillus thuringiensis (2x1012 ufc mL1) a fin de favorecer el crecimiento radicular y evitar el estrés post-trasplante. El trasplante se realizó el 19 de octubre de 2014, con una planta por maceta. Las plantas se manejaron a dos tallos tutorados con rafia de plástico sostenida en hilos de alambre tendidos horizontalmente a lo largo del invernadero; el deshoje y podas se hicieron conforme las plantas lo requirieron.
Las fuentes de calcio (tratamientos) fueron: nitrato de calcio líquido [(CaNO3)2 líq] formulado a 18-0-0-16 (CaO), nitrato de calcio soluble [(CaNO3)2 sol-1] con formulación de 10.1 0-0-17.3 (CaO), nitrato de calcio soluble [ (CaNO3)2 sol-2] con fórmula 15.5-0-0-26.5 (CaO) y nitrato de calcio soluble [(CaNO3)2 sol-3] con fórmula 15-0-0-26 (CaO), a partir de las cuales se elaboraron las siete dosis o tratamientos siguientes: 21, 20, 19, 18, 17, 16 y 15 L ha-1, al considerar como base que son 18 L.ha-1 (testigo uno) lo que de cada formulación se recomienda comercialmente; cada dosis se aplicó tres veces por semana hasta el momento de la última cosecha de frutos mediante 1 200 mL por maceta; como testigo dos se aplicó la solución Steiner (1961) diariamente, excepto cuando se regó con solución de calcio, durante todo el periodo del cultivo (hasta el tercer corte) con la misma cantidad de mL por maceta. El diseño experimental fue bloques completos al azar, con tres repeticiones (tres macetas con una planta cada una) por tratamiento o dosis y 21 plantas por cada fuente de calcio.
Por medio de charolas colocadas por debajo de las macetas se capturó el agua drenada para analizar el pH y CE de la misma. Después de 18 días de haber aplicado los tratamientos, las muestras se tomaron cada tercer día, para hacer un total de 25 muestreos. El pH= 1.8 de entrada y pH= 4.5 de salida se determinaron con medidor portátil HANNA modelo HI-98128 y la CE= 1.2 de entrada y CE= 1.6 dS m-1 de salida se determinaron con medidor portátil HANNA modelo HI-98331.
Las variables de estudio fueron contenido de clorofila, altura de plantas, peso fresco y seco de plantas, peso fresco y seco de frutos, así como el rendimiento por hectárea. El índice de verdor fue cuantificado por medio de un medidor de clorofila Minolta Spad-502 previamente calibrado, la altura se midió con un flexómetro, el peso seco se obtuvo mediante secado en estufa a 85 °C hasta peso constante, de tal forma que el peso fresco y seco de plantas y frutos, así como el rendimiento se determinaron con báscula de precisión marca Ohaus. Los datos se analizaron estadísticamente con el procedimiento proc glm del SAS Institute (1985) versión 6.12, utilizando la prueba de comparación múltiple de medias Duncan, con α≤ 0.05, y la correlación entre variables se hizo con el procedimiento proc corr del mismo programa estadístico.
Resultados y discusión
El índice de verdor (Cuadro 1) en donde puede notarse que los promedios fueron estadísticamente similares; sin embargo, en las plantas cultivadas con (CaNO3)2 sol-3 se tuvo un incremento de 4.1% con respecto al promedio de las plantas que se manejaron con (CaNO3)2 líq y de 3.5% en relación a la media de las plantas cultivadas con (CaNO3)2 sol-1. Además, con la solución de (CaNO3)2 líq los respectivos incrementos fueron de 2.6 y 3.5% comparado con los promedios de las plantas en que se aplicaron (CaNO3)2 sol-1 y (CaNO3)2 sol-2.
Cuadro 1 Contenido de clorofila, altura de plantas, peso fresco y seco de plantas de pimiento morrón cultivado con cuatro formulaciones de calcio.
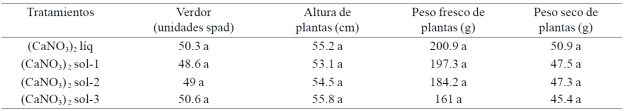
Medias con la misma letra en la columna son estadísticamente iguales Duncan, α≤ 0.05).
Los promedios de altura, peso fresco y seco de plantas tampoco fueron estadísticamente diferentes, pero con (CaNO3)2 sol-3 y (CaNO3)2 líq se obtuvieron incrementos de altura de 5.1 y 2.4% en comparación con la de aquéllas que fueron cultivadas con (CaNO3)2 sol-1 y (CaNO3)2 sol-2, respectivamente; el mayor incremento (24.8%) de peso fresco se obtuvo con (CaNO3)2 líq en comparación con el promedio de las plantas con (CaNO3)2 sol.3, pero con respecto a las que se cultivaron con (CaNO3)2 sol-1 fue de 1.8% y de 9.1% en relación a la que expresaron las que se manejaron con (CaNO3)2 sol-2. El peso seco de las plantas tuvo su mayor expresión con la solución de (CaNO3)2 líq, de tal manera que comparado con lo que lograron las que se cultivaron con (CaNO3)2 sol-1, (CaNO3)2 sol-2 y (CaNO3)2 sol-3, los respectivos incrementos fueron de 7.2, 7.6 y 12.1%.
La correlación entre clorofila y la altura de las plantas fue de r= 0.93* (p= 0.05); entre la altura y el peso fresco y seco de las plantas las respectivas relaciones fueron de r= 0.99** (p= 0.001) y r= 0.90* (p= 0.05). El valor de la relación entre clorofila y el peso fresco de las plantas fue de r=0.92* (p= 0.005) y con el peso seco alcanzó el valor de r= 0.83 ns (p=0. 09); mientras que la relación entre el peso fresco y el peso seco de las plantas fue r=0.83 ns (p= 0.16).
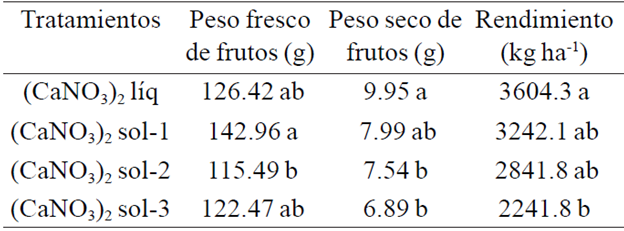
Con los valores del Cuadro 2 se pudo calcular que con la solución de (CaNO3)2 sol-1 el peso fresco de frutos fue 13.1% superior en relación a lo obtenido con la solución (CaNO3)2 líq; asimismo, 23.8 y 16.7% con respecto a lo que se logró con las respectivas soluciones (CaNO3)2 sol-2 y (CaNO3)2 sol-3; sin embargo, en peso seco la solución de (CaNO3)2 líq fue la que ocasionó incrementos de 24.5, 32 y 44.4% en relación a lo que se obtuvo con las soluciones (CaNO3)2 sol-1, (CaNO3)2 sol-2 y (CaNO3)2 sol-3, respectivamente. A su vez, el peso seco obtenido de los frutos cosechados de las plantas cultivadas con el (CaNO3)2 sol-1 superó en 6% al que se logró con (CaNO3)2 sol-2 y 16% en comparación al que se obtuvo con (CaNO3)2 sol-3; mientras que con (CaNO3)2 sol-2 el peso en cuestión superó en 9.4% al estimado a partir de los frutos cosechados de las pantas con (CaNO3)2 sol-3. De tal manera que el rendimiento de las plantas con (CaNO3)2 líq superó en 11.2, 26.8 y 60.8% a los promedios que se obtuvieron con (CaNO3)2 sol-1, (CaNO3)2 sol-2 y (CaNO3)2 sol-3, respectivamente. La correlación entre el peso seco de los frutos y el rendimiento por hectárea fue r=0.97 (p= 0.0001).
Cuadro 2 Promedios de peso fresco, seco y rendimiento de frutos de pimiento morrón cultivado con cuatro formulaciones de calcio.
Con los valores del Cuadro 3 se pudo calcular que con la dosis de 17 L.ha-1 de (CaNO3)2 líq, el promedio de peso fresco de frutos superó en 43.5% al promedio que se obtuvo con la solución Steiner y en 26.9% al que se logró con la dosis comercial (18 L ha-1) de dicha solución; con la dosis de 15 L ha-1 de (CaNO3)2 sol-1 el promedio superó en 77.1 y 51.5% a los respectivos promedios que se lograron con solución Steiner y la dosis comercial de dicha formulación, mientras que con la dosis de 16 L ha-1 los incrementos fueron de 82.6 y 56.3%; con (CaNO3)2 sol-2 la mejor dosis fue la comercial (18 L ha-1), ya que se tuvo un incremento de 52.2% en relación al promedio logrado con solución Steiner; mientras que con (CaNO3)2 sol-3 la mejor dosis fue la de 16 L ha-1, toda vez que con ella el incremento fue de 24.6% en comparación al promedio obtenido con solución Steiner y 23.4% al que se obtuvo con la dosis comercial de la misma formulación.
Cuadro 3 Peso fresco de pimiento morrón cultivado con diferentes dosis de soluciones de calcio.
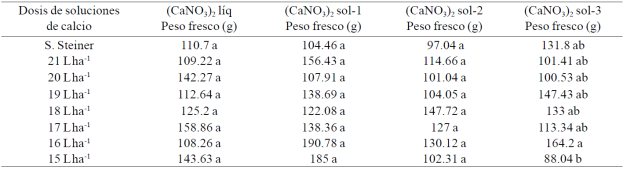
Medias con la misma letra en la columna son estadísticamente iguales Duncan, α≤ 0.05).
A partir de los promedios que se indican en el Cuadro 4 se pudo calcular que con la dosis de 17 L ha-1 de (CaNO3)2 líq, el peso seco de los frutos se incrementó 15.8% en relación al promedio que se logró con la solución Steiner y 33.2% con respecto a la dosis comercialmente recomendada; la mejor dosis de (CaNO3)2 sol-1 fue la 15 L ha-1, ya que el incremento fue de 2.5 veces más en comparación al promedio que logrado con la solución Steiner y de 61% con respecto a lo obtenido con la dosis comercial. A partir del (CaNO3)2 sol-2, la mejor respuesta se observó donde se aplicó la dosis comercial (18 L ha-1), misma que superó en 58.4% al promedio de peso seco obtenido con la solución Steiner; en tanto que del (CaNO3)2 sol-3, la dosis de 19 L ha-1 fue la que indujo mayor respuesta, toda vez que el incremento fue de 37.4% en relación al que se logró con la solución Steiner y 41.5% comparado con lo producido con la dosis comercial.
Cuadro 4 Peso seco de pimiento morrón cultivado con diferentes dosis de soluciones de calcio.
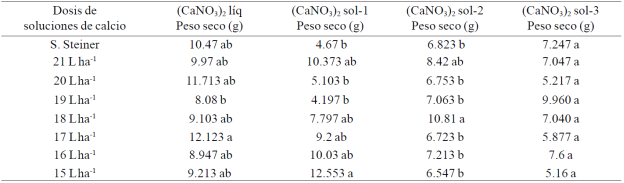
Medias con la misma letra en la columna son estadísticamente iguales Duncan, α≤ 0.05).
Los incrementos relativos del índice de verdor, altura, peso fresco y seco de las plantas cultivadas con la solución de (CaNO3)2 líq, indican que ésta es una práctica con la que se puede aplicar Ca2+ al suelo, y que con ella se facilita más su difusión hacía el interior de las plantas de pimiento, ya que según Salisbury y Ross (2000) el Ca2+ se difunde hacia el interior de la planta a favor de un gradiente de concentración y se transporta hacia fuera con la ayuda de bombas dependientes de ATP. De tal manera que con la solución de (CaNO3)2 líq las zonas meristemáticas de las raíces, tallos y hojas, donde ocurrieron divisiones celulares, fueron las más beneficiadas, quizás porque se necesitó calcio para que formaran nuevas laminillas medias en la placa celular que debieron aparecer entre las células hijas.
También tienen relación con lo reportado por Lea y Leegood (1993), quienes descubrieron que en la célula ocurren secuencias de reacciones que requieren Mn2+, Ca2+ y Cl-ligados a un conjunto de polipéptidos, lo que quizás ocurrió en las plantas de pimiento, toda vez que al activarse enzimas que favorecen la síntesis de clorofila, ésta se incrementó y, en consecuencia, el proceso de fotosíntesis para generar más materia fresca y seca en los frutos y plantas en general. Materia que quizás también se incrementó debido a la fosforilación de proteínas en el núcleo, con las consecuencias en la división celular y en el rendimiento por unidad de superficie.
Estos resultados tienen relación con lo que reportan Salisbury y Ross (2000), ya que al existir más calcio disponible éste puede ser absorbido e incorporado en las partes celulares e influir en la división de las células para inducir el crecimiento y peso de los frutos, que son dos componentes del rendimiento de pimiento por unidad de superficie. Además, permiten comprender de mejor manera las diferencias en extracción de Ca2+ que mencionan Charlo et al. (2012), ya que éstos reportan que el pimiento puede extraer 81.3 kg ha-1, mientras que Azofeifa y Moreira (2005) encontraron 38 kg ha-1 y Fontes et al. (2005) mencionan que el pimiento extrae 114 kg ha-1. Es decir, dependiendo del estado del ión Ca2+ y la dosis, en este caso cada una de las formulaciones de donde se elaboraron las dosis, será la facilidad con que éste pueda ser absorbido por las plantas.
Los resultados que se lograron con las formulaciones de (CaNO3)2 líq y (CaNO3)2 sol-1, en términos de materia seca y rendimiento por unidad de superficie, también tienen fuerte relación con lo reportado por Gordillo et al. (2004), toda vez que éstos descubrieron que el Ca2+ es el elemento que asegura procesos, tales como la síntesis de paredes celulares, en la
lámina media, donde forma pectato de calcio que confiere estabilidad y mantiene la integridad de éstas. Asimismo, los resultados de esta investigación se relacionan con los de Chiu y Bould (1977), ya que éstos refieren que el Ca2+ es poco móvil y tiende a acumularse en los órganos más viejos, mientras que en los de mayor actividad metabólica (hojas en crecimiento, flores, frutos y meristemos apicales) se necesita mayor aporte, donde queda fijado y prácticamente inmóvil en sus paredes celulares. La relación también es evidente con lo reportado por Marschner (2002) quien ha mencionado que con altas dosis de Ca2+ las cantidades de este elemento que no logran entrar al citosol quedan en la pared celular, situación que genera células más pesadas.
Los efectos ocasionados por las dosis más bajas de Ca2+ (15 y 17 L ha-1) de las soluciones hechas con las formulaciones (CaNO3)2 sol-1 y (CaNO3)2 líq, respectivamente, coinciden con lo reportado por Williamson (1984), quien menciona que las concentraciones bajas de Ca2+, casi micromolares, deben mantenerse en parte para impedir la formación de sales de calcio insolubles, obtenidas a partir de ATP y de otros fosfatos orgánicos. Además, las concentraciones de Ca2+por encima del margen micromolar inhiben la corriente citoplasmática. De tal manera que con las dosis aquí mencionadas (15 y 17 L ha-1) quizás se limitó más la formación de sales de calcio insolubles y se favoreció la corriente citoplasmática, para que en las plantas de pimiento ocurrieran las reacciones químicas necesarias y dichas plantas produjeran más materia seca.
Conclusiones
Con las soluciones de (CaNO3)2 líq y (CaNO3)2 sol-1 se cultivó pimiento morrón de manera más sustentable que con las soluciones de (CaNO3)2 sol-2 y (CaNO3)2 sol-3, ya que para mejor respuesta en rendimiento se necesitó de una dosis menor a la recomendada comercialmente y, en consecuencia, disminuyó la posibilidad de que a través del tiempo y por alta concentración el calcio se convirtiera en un agente contaminante en el sustrato utilizado.

