











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El cultivo del maíz en condiciones de secano o temporal está expuesto a amplias variaciones en cantidad y distribución de la lluvia durante su crecimiento; estas variaciones en la disponibilidad de humedad frecuentemente causan estrés hídrico, que a su vez disminuye la acumulación de materia seca en los órganos aéreos de la planta, el desarrollo del área foliar y la formación de grano, factores que finalmente se reflejan en la pérdida de rendimiento. Cuando las deficiencias de humedad se presentan en diferentes periodos del ciclo del cultivo (sequía intermitente), la sequía puede causar estrés hídrico en las etapas de plántula y reducir el establecimiento del cultivo; en etapas fenológicas más avanzadas, el estrés puede afectar el desarrollo del área foliar y la fotosíntesis. Si la sequía ('sequía de medio verano' o 'canícula'), se presenta previo a la floración y/o en las etapas de floración y llenado del grano, el déficit hídrico reduce la producción de mazorcas y granos durante las dos semanas de la floración, debido a una disminución en la tasa de fotosíntesis durante el llenado del grano y una rápida inducción de la senescencia de las hojas; el rendimiento de grano también puede ser afectado por la competencia por foto-asimilados entre los órganos aéreos de la planta y las raíces como una respuesta de adaptación a la sequía (Bánziger et al., 2012).
En general, la elongación de las hojas depende dinámicamente de la disponibilidad de agua en el suelo; y en el maíz no es la excepción. Con una disponibilidad adecuada de agua en el suelo, la elongación se mantiene constante, sin embargo, cuando el potencial hídrico del suelo disminuye, la elongación también se reduce. Los cambios en el status hídrico apuntan al papel directo que el agua tiene en el crecimiento; su absorción provee la fuerza física para el alargamiento de las células (Acevedo et al, 1971).
Un fenómeno comúnmente observado bajo sequía impuesta en la floración es el alargamiento del intervalo antesis-floración femenina (Bolaños y Edmeades, 1990). Esto ocurre usualmente por un retraso en la emergencia del jilote en relación a la emergencia de las anteras, esta última siendo menos afectada por la sequía. En este contexto, el intervalo antesis-floración femenina es considerado más valioso como carácter de diagnóstico del comportamiento del genotipo que la fecha de floración femenina per se, debido a que el intervalo antesis-floración femenina es bastante independiente de las diferencias en madurez fisiológica entre variedades (Edmeades et al, 1989). La disponibilidad de nitrógeno constituye otro de los factores limitantes del crecimiento y rendimiento en condiciones de secano; si las plantas experimentan estrés por deficiencias de este elemento, se reduce la tasa fotosintética porque se disminuye el desarrollo del área foliar y se acelera la senescencia de las hojas (Bánziger et al, 2012).
En maíz, un nivel bajo de nitrógeno reduce la capacidad de crecimiento del grano durante su etapa de llenado (Bánziger et al, 2000), En condiciones de estrés hídrico y deficiencias de nitrógeno, en el mejoramiento genético podría, incluir características agronómicas relacionadas con la fenología, tasa fotosintética, acumulación de biomasa y partición de foto-asimilados a la mazorca (Bolaños y Edmeades, 1993). Por tanto, el objetivo del presente trabajo de investigación fue estudiar la respuesta de híbridos y variedades criollas de maíz, con riego y sequía y dos niveles de fertilización con nitrógeno, en el desarrollo del área foliar, la floración, el rendimiento de grano y sus componentes, en condiciones de invernadero.
Materiales y métodos
Sitio experimental y material vegetal
El experimento se realizó en invernaderos del área de resistencia a sequía, del Colegio de Postgraduados en Montecillo, Estado de México (19° 21' latitud norte, 98° 55' longitud oeste y 2 250 msnm). El invernadero empleado para este experimento tenía techo de material de plástico transparente. Se emplearon cuatro híbridos de maíz producidos por el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), estos híbridos se caracterizan por estar desarrollados para áreas de secano de los Valles Altos de la Mesa Central de México y dos materiales criollos colectados en el municipio de Españita, Tlaxcala. El material genético del INIFAP, representa a los híbridos comerciales producidos entre los años de 1960 y 2000. Los híbridos liberados entre 1961 y 1973 se catalogan como híbridos antiguos y los liberados en 1999 como híbridos modernos (Cuadro 1).
Cuadro 1 Híbridos y materiales criollos de maíz, evaluados en diferentes condiciones de sequía y fertilización nitrogenada.

†Híbrido de cruza doble, 'Híbrido trilineal; §Germoplasma recolectado en el Municipio de Españita, Tlaxcala, por el Dr. Cándido López-Castañeda, Programa de Postgrado en Recursos Genéticos y Productividad-Genética, Colegio de Postgraduados. Montecillo, Estado de México en el ciclo otoño-invierno, 2013.
Diseño experimental
Se utilizó un diseño de bloques completos al azar en arreglo factorial 4 x 2, con dos repeticiones, correspondientes a cuatro tratamientos de humedad edáfica y dos niveles de fertilización para todos los materiales. Las semillas se sembraron en un tubo de PVC de 4''de diámetro y 1 m de largo, con una bolsa cilíndrica de plástico de las mismas dimensiones en su interior, conteniendo suelo esterilizado; el suelo utilizado fue de textura migajón-limoso (34 % de arena, 56% de limo y 10% de arcilla) al que se le determinó su capacidad de campo (CC) de 33.9%, porcentaje de marchitamiento permanente (PMP) de 21.1 % y densidad aparente de 1.08 g cm3 (laboratorio de física de suelos, Colegio de Postgraduados, Montecillo, Texcoco, Estado de México). La siembra se realizó el cinco de mayo de 2013, se emplearon semillas con un peso de 350±5 mg en cada unidad experimental. Se evaluaron cuatro tratamientos de humedad edáfica (HE): T1= riego (R) que consistió en mantener la HE cercana a capacidad de campo (CC) hasta la madurez; T2= sequía 1 (S1) sin R por 10 días (d) cuando las plantas tuvieron 30 d después de la siembra (dds); T3= sequía 2 (S2) sin R por 20 d cuando las plantas tenían 50 dds, y T4= sequía 3 (S3) sin R por 30 d para plantas con 70 dds; al finalizar los periodos de sequía se aplicó un riego de recuperación (RR) en cada tratamiento de deficiencias hídricas edáficas con la aplicación de riegos subsecuentes hasta la madurez. También se evaluaron tratamientos de fertilización nitrogenada que se aplicaron al momento de la siembra; dosis alta de nitrógeno (AN) con 160-40-20 y dosis baja (BN) con 80-40-20, como fuente de nitrógeno, fosforo y potasio se usaron: urea, superfosfato de calcio triple y cloruro de potasio, respectivamente. Para el control de pulgón y mosquita blanca se aplicó 1 L ha-1 Engeo® a los 38 dds.
Variables cuantificadas y análisis estadístico
Se midió el potencial hídrico foliar (ψ, bares) con la bomba de presión o Scholander en la porción media de hojas completamente expandidas y expuestas a la radiación solar, entre las 11:00 y 13:00 h a los 30, 35 y 40 dds en T2= S1; 50, 55, 60, 65 y 70 dds en T3= S2, y 70, 75, 80, 85, 90, 95 y 100 dds en T4= S3 con AN y BN, y a los 41, 71 y 101 dds (un día después del RR) en todos los tratamientos.
El potencial osmótico (π, bares) se determinó en las muestras de hojas utilizadas para medir el ψ en todos los tratamientos de HE y N del suelo, y en las fechas antes indicadas; posteriormente estas muestras se colocaron en viales de plástico de 5 mL y se depositaron en un termo con N líquido (-195 °C) durante dos minutos, para romper la pared celular y extraer la savia; el π se midió con un osmómetro marca Wescor, modelo 5100C en el laboratorio (Barrios-Gómez et al, 2011).
El potencial de turgencia (P, bares) se calculó como la diferencia entre los potenciales ψ y π (p= ψ - π), para cada muestra analizada (Begg y Turner, 1976). El contendido relativo de agua (CRA, %) se determinó en todas las plantas, al extraer 10 discos de 5 mm de diámetro de la misma porción de la hoja utilizada para medir el ψ, π y p; los discos se pesaron inmediatamente para obtener el peso fresco (Pf, mg) y se colocaron en cajas petri, para quedar suspendidas en agua destilada durante 10 horas bajo iluminación, para obtener el peso a saturación completa del tejido (Psat, mg) y posteriormente, el peso de materia seca (Ps, mg), al secar las muestras de los discos de hoja en una estufa a 70 °C durante 48 h, [CRA=(Pf-Ps)/(Psat-Ps)100].
La expansión del área foliar (EAF) por planta (EAF=L*A*0.75, cm2), se determinó al medir el incremento en el área de las hojas en elongación cada cinco días, en todos los tratamientos. El número de días a antesis o floración masculina (FM); se determinó cuando 50% de las espiguillas presentaban anteras en dehiscencia y expuestas fuera de las glumas. El número de días a floración femenina (FF); se determinó cuando la planta presentó el jilote o inflorescencia femenina con los estigmas expuestos. Se determinó también el rendimiento de grano por planta (RG, g) y sus componentes número de granos normales (GN) por mazorca y el peso (g) de 100 granos.
El contenido hídrico edáfico (CHE) se determinó al pesar los tubos tres veces por semana en todos los tratamientos de HE y N; en R se agregó la cantidad de agua perdida por transpiración entre fechas sucesivas de pesaje, manteniendo la humedad (H) cercana a CC hasta la madurez fisiológica (MF). Se aplicó RR en S1, S2 y S3 al finalizar los tratamientos de sequía, y se aplicaron riegos subsecuentes después del RR hasta la MF. La cantidad total de agua transpirada se calculó, al sumar las pérdidas de agua estimadas en cada tubo durante el experimento.
Análisis estadístico
Se calculó el análisis de varianza para todas las variables con el programa SAS para Windows Versión 9.0 (SAS, 2002) en forma individual. La comparación de medias se realizó con la prueba de Tukey (p≤ 0.05).
Resultados y discusión
Contenido hídrico del suelo
En S1 se suspendió el R entre los 30 y 40 dds y la H aprovechable disminuyo 75% sin alcanzar el PMP (Figura 1a); en S2 el R se suspendió entre los 50 y 70 dds y la H aprovechable descendió por debajo de PMP entre los 60 y 70 dds (Figura 1b), y en S3 se dejó de aplicar R entre los 70 y 100 dds y la humedad del suelo disminuyó por debajo del PMP entre los 88 y 100 dds (Figura 1c).
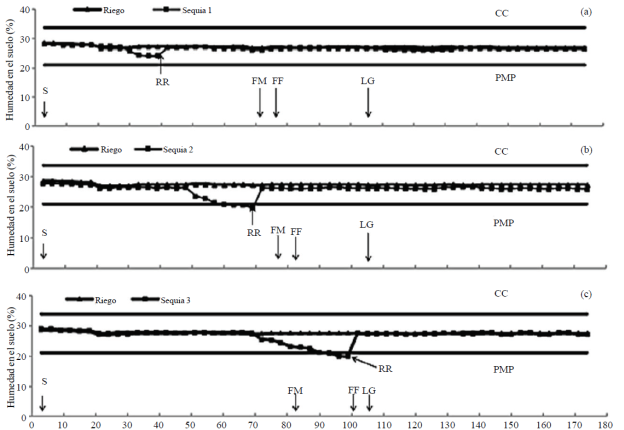
Figura 1 Contenido hídrico del suelo en riego y sequia 1 (a), riego y sequia; 2 (b) y riego y sequia 3 (c) en invernadero, ciclo verano-otoño 2013. Montecillo, Estado de México. CC= capacidad de campo; PMP= porcentaje de marchitamiento permanente; S= siembra; FM= floración masculina; FF= floración femenina; LG= llenado de grano; RR= riego de recuperación.
La reducción en el CHE se reflejó en una disminución considerable en el foliar en S1 (ψ < -12 bares), S2 (ψ < -25 bares) y S3 (ψ < -30 bares) con AN y BN con respecto a R (ψ > -5 bares), con valores más negativos para el π y niveles más bajos para el P y el CRA, al finalizar los tratamientos de deficiencias hídricas edáficas. En S2 las plantas mostraron síntomas severos de marchitamiento, pero fueron más acentuados en S3, donde la sequía coincidió con la FM, FF y el inicio del llenado del grano. Un estudio en frijol demostró que las deficiencias hídricas edáficas durante la floración, disminuyen severamente el contenido hídrico de la planta, reduciendo considerablemente el rendimiento de semilla, biomasa aérea, número de vainas (Barrios-Gómez et al, 2010).
Relaciones hídricas
Para el tratamiento de riego con los dos niveles de fertilización, se presentaron valores de ψ, π y p, y CRA entre -1 a -5, -10 a -13 y 8 a 11 bares, y 78 a 83%, respectivamente y no presentaron diferencias significativas entre los maíces evaluados, los materiales no presentaron estrés hídrico en ninguna fecha de medición. En el ambiente de sequía (S1) con AN (Figuras 2a, b, c y d) y BN Figuras 2e, f, g y h), se tuvieron los siguientes rangos de variación: ψ (-3 a -16 bares), π (-14 a -19 bares) y p (2 a 12 bares), y CRA (50 a 80%).

Figura 2 Potencial hídrico (a y e), osmótico (b y f) y de turgencia (c y g), y contenido relativo del agua (d y h) en sequía 1 con dosis alta (a, b, c, y d) y baja de nitrógeno (e, f, g y h) en condiciones de invernadero, ciclo de verano-otoño 2013. Montecillo, Estado de México. [* (p≤ 0.05); ns= no significativo].
En S2 el ψ, π y p, y CRA con AN (Figuras 3a, b, c y d) y BN (Figuras 3e, f, g y h) variaron de -4 a -29, -12 a -20 y 9 a -8 bares, 20 a 85%, respectivamente.
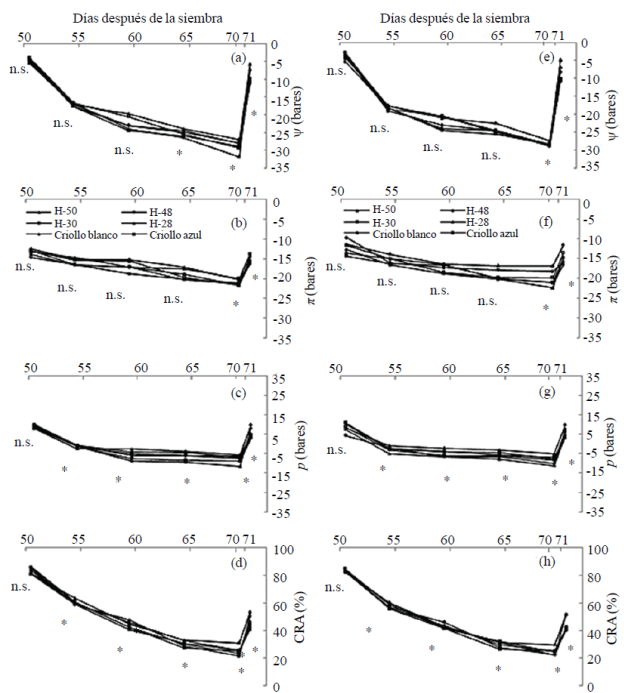
Figura 3 Potencial hídrico (a y e), osmótico (b y f) y de turgencia (c y g), y contenido relativo del agua (d y h) en sequía 2 con dosis alta (a, b, c, y d) y baja de nitrógeno (e, f, g, y h) en condiciones de invernadero, ciclo de verano-otoño 2013. Montecillo, Estado de México. [* (p≤ 0.05); ns= no significativo].
Para S3 con AN (Figura 4a, b, c y d) y BN (Figuras 4e, f, g y h), los rangos de variación fueron: ψ (-6 a -30 bares), π (-12 a -24) y p (6 a -5 bares), y CRA de 18 a 85%. Para todos los tratamientos de sequía los ψ, π y p, y CRA presentaron diferencias (p≤ 0.05) entre genotipos en algunas fechas de medición y se alcanzaron valores muy bajos conforme transcurrió el periodo de sequía.
La sequía disminuyó el ψ a -18, -22 y -15 bares en diferentes genotipos de maíz, sorgo y tabaco al medio día en las hojas superiores del dosel del cultivo; el π también disminuyó aunque no al mismo grado que lo hizo el ψ, de tal forma que el p tuvo valores de cero en algunas hojas; la disminución del π en las hojas más altas del dosel del cultivo, dio como resultado que estas hojas tuvieran más alto p a un ψ dado; la resistencia estomática de las hojas superiores aumentó a valores más negativos del y en los tres cultivos utilizados y se observó también que las hojas superiores en las plantas de tabaco, se marchitaron a valores más negativos del ψ que las hojas inferiores del dosel del cultivo (Turner, 1974).
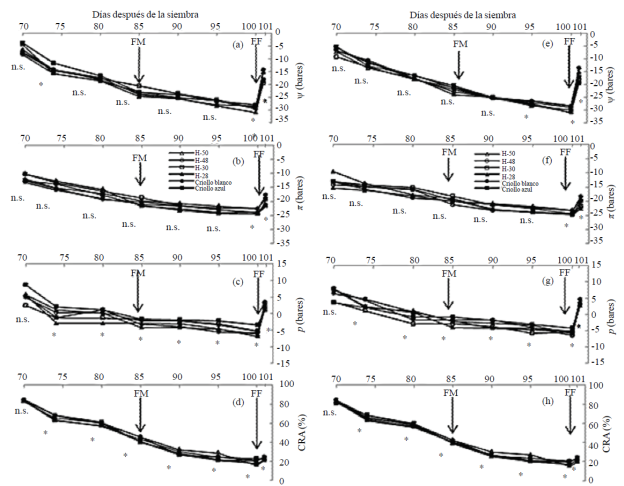
Figura 4 Potencial hídrico (a y e), osmótico (b y f) y de turgencia (c y g), y contenido relativo del agua (d y h) en sequía 3 con dosis alta (a, b, c, y d) y baja de nitrógeno (e, f, g y h) en condiciones de invernadero, ciclo de verano-otoño 2013. Montecillo, Estado de México. [* (p≤ 0.05); ns= no significativo; FM= floración masculina; FF= floración femenina].
Al comparar el comportamiento de un híbrido de maíz con Sorghum halapense, bajo riego (75% CC) y déficit hídrico (23.5-25% CC) durante 24 d, se determinó que el Sorghum halapense realizó una absorción continua de agua, lo que le permitió alcanzar un mayor y con respecto al híbrido de maíz y mantener el CRA por encima de 70% en condiciones de baja disponibilidad hídrica, lo que se reflejó en un intercambio gaseoso activo (Acciaresi et al., 2012).Avendaño et al. (2005) observaron que las variedades mejoradas de maíz Zacatecas-58 (ciclo 19 de selección masal) y Cafime (ciclo 16 de selección masal), presentaron menor ψ y π que las variedades originales (Zacatecas 58 y Cafime) sometidas a condiciones de sequía edáfica (10 d en PMP del suelo).
En el presente estudio se observó que el comportamiento fisiológico del germoplasma bajo condiciones de estrés hídrico y deficiencias de nitrógeno (DN) en el suelo fue similar en la etapa vegetativa (S1), antes del inicio de la floración (S2) y durante la floración y el inicio del llenado del grano (S3), sin que se hayan observado diferencias en el ψ, π y P, y CRA entre las plantas con AN y BN. También, se observó que los genotipos que mantuvieron valores de ψ, π y P, y CRA más altos al finalizar los tratamientos de sequía, reaccionaron más rápidamente al riego de recuperación.
Expansión del área foliar
La mayor reducción en la EAF de la planta se observó en los tratamientos de sequía (14.2 %) en comparación con los de riego (Cuadro 2). En la etapa vegetativa se observaron los valores más bajos de la expansión del área foliar (17.1%) (S1, 30- 40 dds), en comparación con las otras etapas fenológicas del cultivo: floración e inicio del crecimiento de grano. La mayor expansión de área foliar (EAF) la presentaron tres de los seis materiales evaluados: H-50, H-48 y criollo blanco, en los cuatro tratamientos evaluados.
Cuadro 2 Expansión del área foliar acumulada en seis genotipos de maíz bajo riego, sequia 1, sequia 2 y sequia 3 con dosis alta (AN) y baja de nitrógeno (BN) a los 40, 70 y 100 días después de la siembra, en condiciones de invernadero. Ciclo de verano-otoño 2013. Montecillo, Texcoco, Estado de México.
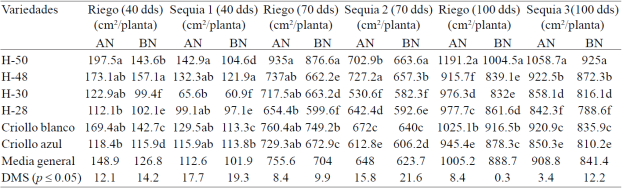
El mayor efecto del estrés hídrico en la expansión del área foliar en la etapa vegetativa (S1) se debió a que el área foliar de las plantas al inicio del ciclo, se encontraba en plena expansión y en esta etapa fenológica, la deficiencia de agua tiene mayor efecto al reducir la velocidad de elongación de las hojas que cuando las plantas han extendido completamente el área foliar (S2), incluyendo la hoja bandera (S3), donde el efecto de las deficiencias hídricas tiene menor efecto en la elongación de las hojas; Bánziger et al. (2012) determinaron que la sequía durante el periodo anterior a la floración en maíz disminuye el desarrollo del área foliar y la tasa fotosintética, y estas reducciones en el tamaño del área fotosintética y la actividad fotosintética de la planta, se reflejan en una drástica reducción de la producción de mazorcas y granos durante las dos semanas de la floración. Las diferencias en la EAF también se observaron entre los tratamientos de AN y BN; la EAF disminuyó 19.4, 3.8 y 7.4% por efecto del BN en Sb S2 y S3 (Cuadro 2), y 14.8, 6.8 y 11.6% en los respectivos tratamientos de R (Cuadro 2).
Vos et al. (2005), al someter plantas del hibrido Lincoln de maíz a dosis de N que variaron de 0.5 a 6 g por planta en condiciones de invernadero, observaron que el bajo N redujo hasta 29% el área foliar total y 16% el área final de la hoja más grande de la planta, lo que confirma la importancia del N en la EAF en maíz con deficiencias de N.
Floración
En R, S1 y S2 el H-48 y H-50 con AN y BN tuvieron menor número de días A y FF que los demás genotipos y en S3 el criollo azul tuvo menor número de días a FM y FF que los otros genotipos (Cuadro 3); diferencias similares entre híbridos y criollos fueron observadas en condiciones de campo con R, sequía y alto y bajo N, donde los criollos bajo sequía tuvieron menor número de días a A que los híbridos, sin diferencias entre criollos e híbridos para el número de días a FF y el intervalo antesis-floración femenina (IA-FF) (Serrem et al., 2009). Los efectos mayores de la deficiencia hídrica se manifiestan en el desarrollo floral y fallas en la fertilización y aborto de cigotos (Saini y Westgate, 2000), receptividad del estigma (Bassetti y Westgate, 1993) y el proceso de polinización (Sawhney y Shukla, 1994).
Cuadro 3 Días a antesis (A) y floración femenina (FF) en seis genotipos de maíz bajo riego, sequía 1, sequía 2 y sequía 3 con alto (AN) y bajo nitrógeno (BN), en condiciones de invernadero. Ciclo de verano-otoño 2013. Montecillo, Texcoco, Estado de México.
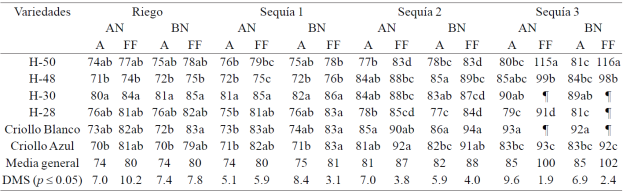
¶ Sin floración.
La sequía retrasó (p≤ 0.05) en promedio la A y la FF en 0.5, 7.5 y 11 d, y 0.5, 7.5 y 21 d con relación a riego en S1, S2 y S3, respectivamente. La deficiencia de nitrógeno retrasó la antesis un día en S1 y S2, la FF se retrasó un día en S1 y S2, y dos d en S3, sin presentar cambios en riego. Los efectos del contenido hídrico edáfico también se observaron en el IA-FF en S3; el IA-FF se alargó 10 d en S3 con respecto a R, sin presentar cambios en R, S1 y S2; este alargamiento en el IA-FF fue tan importante que evitó la formación de grano en todas las plantas de maíz en S3; las deficiencias de nitrógeno sólo tuvieron efecto en las plantas en S3, donde el IA-FF fue nueve días más largo que en R con AN y 11 días más largo que en R con BN. En S3 el jilote del H-30 y el criollo blanco con AN, y el H-30, H-28 y el criollo blanco con BN no emitieron los estigmas debido a estrés hídrico (Cuadro 3).
El número de días a A y FF se retrasó siete días en promedio en las plantas de maíz con BN y ocho d en las plantas con AN en S2, y 11 y 20 d con AN, y 11 y 22 d con BN en S3. En promedio el IA-FF en presencia de EH severo (S3) fue mayor con AN (15 d) y BN (17 d) que en R con AN y BN (seis d); S1 con AN y BN (seis d); y S2 con AN (seis d) y BN, donde la asincronía entre las floraciones fue mayor (15 d con AN y 17 d con BN) que en R (seis d con AN y BN), S1 (seis d con AN y BN) y S2 (seis d con AN y BN). Estudios en condiciones de campo muestran que la sequía retrasó 1.5 d la A y tres d la FF con respecto a R y el IA-FF aumentó tres d en líneas S1 de criollo de Ibarrilla, por efecto de la sequía (Reyes-Ramones, 2000) y en condiciones de sequía en invernadero, se determinó que la sequía retrasó 19 d la A y 18 d la FF en promedio de cuatro variedades de maíz (Cafime original, Zacatecas 58 original, Cafime SM16 y Zacatecas 58 SM19), sin cambios significativos en el IA-FF (Avendaño et al., 2008).
Rendimiento de grano y sus componentes
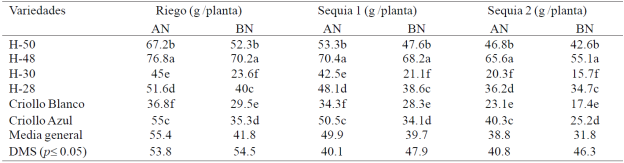
En S1 y S2, el EH redujo 8 y 27% el RG y las deficiencias de nitrógeno redujeron el RG 25, 20 y 18% en R, S1 y S2; el H-48 obtuvo mayor RG (p 0.05) que los demás genotipos en R y S1 con AN y BN; el H-48 y H-50 obtuvieron mayor RG (p 0.05) que el resto de los materiales genéticos en S2 con AN y BN (Cuadro 4). En S3 no se produjo grano en ninguno de los materiales evaluados.
Cuadro 4 Rendimiento de grano de seis genotipos de maíz evaluados bajo riego, sequía 1 y sequía 2 con alto (AN) y bajo nitrógeno (BN), en condiciones de invernadero. Ciclo verano-otoño 2013. Montecillo, Texcoco, Estado de México.
El número de granos normales mazorca-1 (NGNM) también se redujo por efecto de la sequía aunque esta reducción fue de menor magnitud que el RG; el NGNM disminuyó 8 y 15% en S1 y S2, y 2.3, 5.7 y 3.5% por efecto del BN en R, S1 y S2. Las deficiencias de nitrógeno aumentaron el número de granos abortados por mazorca (NGAM) 32% en S1 y 2% en S2. El peso de 100 granos (P100G) se redujo por la sequía en 13 y 40% en S1 y S2, y 29, 27 y 20% por efecto del bajo N en R, S1 y S2, respectivamente (Cuadro 5).
Cuadro 5 Número de granos normales mazorca-1 (GN) y peso de 100 granos (P100G) en seis genotipos de maíz evaluados bajo riego, sequía 1 y sequía 2 con alto (AN) y bajo nitrógeno (BN) en condiciones de invernadero, ciclo verano-otoño 2013. Montecillo, Texcoco, Estado de México.

La respuesta de los materiales de maíz a la sequía y las deficiencias de nitrógeno en el RG y sus componentes varió con la etapa fenológica, la duración y la intensidad del EH en los diferentes tratamientos de HE; en S1 (30-40 dds) la reducción en el RG y sus componentes NGNM y P100G, por efecto de la sequía (Grant et al, 1989) y las deficiencias de nitrógeno fue mucho menor que en S2 (50-70 dds) y sobre todo S3 (70-100 dds) donde las plantas de maíz no produjeron grano. El EH redujo el RG y sus componentes en mayor grado que las deficiencias de nitrógeno en todos los genotipos; el RG se reduce cuando las plantas sufren EH severo durante el periodo de floración (Nielsen, 2013); el aborto de granos y mazorcas al inicio del llenado de grano y la reducción de la fotosíntesis (Bánziger et al, 2012).
Conclusiones
Los híbridos mantuvieron mayor potencial hídrico y de turgencia, y contenido relativo de agua hasta el final de los tratamientos de sequía, tuvieron mayor capacidad de recuperación al estrés hídrico y mayor capacidad de expansión del área foliar en condiciones de alto y bajo nitrógeno. La sequía retrasó la floración masculina y femenina, y alargó el intervalo antesis-floración femenina, redujo el rendimiento de grano y sus componentes número de granos mazorca-1 y tamaño de grano, y este efecto fue mayor en sequía tres, cuando coincidió con la antesis y la floración femenina, causando que las plantas de maíz no produjeron rendimiento de grano en alto y bajo nitrógeno. Los híbridos superaron a los criollos en el rendimiento de grano y granos normales mazorca-1. Los híbridos superaron a los criollos en el P100G en condiciones de sequía con deficiencias de nitrógeno. Con alto y bajo N los criollos alcanzaron la antesis y floración femenina en menor tiempo que los híbridos y tuvieron un intervalo antesis-floración femenina más largo que los híbridos en la sequía de diez días. Los híbridos alcanzaron la antesis y floración femenina en menor tiempo que los criollos y tuvieron un intervalo antesis-floración femenina más corto que los criollos cuando se aplicó la sequía antes de la floración.

