











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En Tenancingo y Villa Guerrero, Estado de México, México, la rosa para flor de corte, con más de 100 diferentes cultivares, se produce bajo cubiertas plásticas en 683 ha (SIAP, 2010) y sólo es superada por gladiolo (Gladiolus grandiflorus Hort.) y crisantemo (Dendranthema spp.). Tetranychus urticae Koch (Acari: Tetranychidae), conocido en la región como araña roja (Kheradpir et al., 2007), es una de las plagas más importantes de rosa (Nyalala et al., 2013) cuyo control en las últimas décadas se ha complicado y encarecido debido a la resistencia a acaricidas (Flores et al., 2007). Por lo tanto la identificación de los mecanismos naturales para su control es deseable. En Tenancingo y Villa Guerrero se ha observado variación natural en la tolerancia de los cultivares de rosa a plagas y enfermedades (Martin et al., 2001; Khatun et al., 2009; Coruh y Ercisli, 2010; Flores et al., 2011), por lo que la identificación de cultivares sobresalientes es indispensable para iniciar programas de mejoramiento genético o generación y/o aplicación de tecnología. El objetivo de esta investigación fue determinar la tolerancia al ataque de araña roja y su efecto en la calidad del tallo floral de 15 cultivares de rosa.
Materiales y métodos
La investigación se hizo en invernaderos del Centro Universitario UAEM Tenancingo de la Universidad Autónoma del Estado de México, localizada a 18º 97’ 03’’ latitud norte y 99º 61’ 17’’ longitud oeste y a 2 200 m de altitud. Se evaluaron 15 cultivares híbridos de tea de Rosa x hibrida (Cuadro 1; Anónimo 1, s.a; Anónimo 2, s.a) elegidos de acuerdo a un trabajo previo sobre percepción de productores en la tolerancia de rosa a plagas y enfermedades.

Los 15 cultivares fueron evaluados en primavera de 2010 y 2012 y otoño de 2010. Los tallos enraizados del patrón Manettii (Rosa sp.) con injerto de yema de los cultivares evaluados se establecieron individualmente en invernadero en macetas de plástico negro de 14 L de capacidad en tierra agrícola, tepojal y germinaza (3:1:1) en un diseño completamente al azar con ocho repeticiones. El pH, medido con Nie-co. nieuwkoop b.v. aalsmeer, Holland, se ajustó a 6.4 con la adición de 120 g m-2 de cal agrícola para el primer ciclo y 100 g m-2 para los dos siguientes. La conductividad eléctrica, medida con HI 98130, Hanna, se ajustó de 0.3 a 2.1 deciSiemens cm-1 con la adición del fertilizante. La fertilización de hizo con 26-12-12+2MgO y micro elementos (Polyfeed Haifa Chemicals, Israel) una semana después del pinzamiento en aplicaciones de 100 g m-2 cada 15 días. El riego se aplicó dos veces por semana en primavera y tres veces por semana en otoño (60% de humedad en el sustrato).
De cada planta se eliminó el primer botón floral para inducir brotación múltiple y posteriormente dejar dos tallos podados a 20 cm de altura en promedio. La infestación se hizo a los 20 días de la poda cuando los nuevos tallos florales tenían 10 cm y consistió en depositar 5 ácaros de T. urticae Koch con movilidad y sin sexar extraídos de cajas de incremento en plantas huésped sanas de frijol (Phaseolus vulgaris L.). Los ácaros se colocaron en el haz de la primera hoja basal y desde la infestación hasta el tercer muestreo (24 días) se registró temperatura (ºC) y humedad relativa (%) diaria con un higrotermógrafo (EL-USB-2 - RH/Temperature Data Logger, LASCAR electronics).
Se realizaron conteos semanales del ácaro por variedad en tres ocasiones, iniciando a la semana de la infestación, definido por el número de ácaros móviles por planta (NA). Con la finalidad de observar efectos de la infestación y daño de ácaros en la fenología de la planta, en la etapa de “punto de corte” se midieron longitud del tallo (LT, medido desde su base hasta el ápice del botón floral), diámetro de tallo (DT, promedio de los diámetros basal, medio y apical del tallo), número de foliolos (NF, de todas las hojas del tallo), pétalos por botón floral (NP), tallos productivos (TP, con presencia de botón floral) y tallos ciegos (TC, los que no lograron la diferenciación floral).
Adicionalmente se midió área foliar (AF, promedio de la superficie (cm2) de las hojas de tres tallos; Area Meter Portable, Model Ll-3000A, y accesorio L1-3050A/4, ambos Ll-COR, USA), y niveles de clorofila en hojas con daño (CCD) y sin daño (CSD) del ácaro, medidas en unidades SPAD (Chlorophyll Meter, Spad-502, Konica Minolta, Japón). Con los datos se realizó un análisis de varianza combinado y la comparación de medias con la prueba de Tukey (α= 0.05). En seis de los cultivares, seleccionados por sus valores medios y extremos en incidencia de ácaros se midió en tres etapas fenológicas el contenido de fenoles totales (mg fenoles totales g-1 peso seco) por el método de Folin y Ciocalteu (Makkar et al., 2007). También se hizo un análisis de componentes principales entre cultivares y variables evaluadas (González et al., 2010) y de conglomerados para definir la agrupación de los cultivares de acuerdo a sus variables. Las salidas se obtuvieron con el Sistema para Análisis Estadístico (Statistical Analysis System, SAS) versión 9 para Windows, y las gráficas del biplot y de conglomerados se obtuvieron con Info Stat Versión 2008.
Resultados y discusión
Grado de infestación
La infestación de ácaros por planta fue estadísticamente significativa (α≤ 0.05) y se identificaron dos grupos de cultivares contrastantes (Figura 1). En el primer grupo el cv. catalina fue el más susceptible al ácaro; el segundo grupo incluyó ambiance como la más resistente. Este hecho es contradictorio ya que en la rosa actual cultivada debería esperarse una reducción en su variación genética debido al avanzado proceso de domesticación y mejoramiento vinculado a sólo 7 de más de 100 especies utilizadas (Martin et al., 2001), pero sus múltiples y complejas hibridaciones posiblemente sean el principal factor de esta variabilidad.

Figura 1 Número de ácaros por planta registrados en 15 cultivares de rosa, desde la formación de brote floral hasta la etapa fenológica de “chícharo grande”. Sobre las barras se indica la desviación estándar de las medias. Barras con la misma letra son iguales significativamente (Tukey, α= 0.05).
En México las empresas dedicadas a la producción de rosa no hacen trabajos de investigación que estén relacionados con el desarrollo de nuevos cultivares a partir de la explotación de la variación genética existente; tampoco se conocen estudios sobre autodefensa de la planta ya sea directa, a través de la síntesis de productos, o indirecta vía atracción de enemigos naturales, contra herbívoros (Agrawal et al., 2002; Boege y Marquis, 2005). Todos los cultivares son importados, lo que incrementa los costos de producción.
Variables ambientales asociadas al desarrollo poblacional del ácaro
Las variaciones en temperatura, humedad relativa y su interacción afectaron el crecimiento poblacional de ácaros por planta y por temporada evaluada (Figura 2). En primavera de 2012 hubo una temperatura de 26 ºC y una humedad relativa de 60%; ambas favorecieron el incremento en la población de T. urticae. Estos resultados coinciden con los de otros estudios donde los intervalos de 25-28 ºC y 60-70% representaron las condiciones óptimas para el desarrollo de esta plaga (Kheradpir et al., 2007). En primavera de 2010 la temperatura promedio fue muy similar a la de 2012, pero la humedad relativa fue del 56.5%. En otoño de 2010 la temperatura y la humedad relativa fueron de 20.7 ºC y 58% por lo que el número de ácaros por planta estuvo por debajo de los registrados en los otros dos ciclos de producción (Figura 2).

Figura 2 Relaciones entre número de ácaros por planta, temperatura y humedad relativa en la variedad catalina registrados en los tres experimentos.
Las temperaturas óptimas para el desarrollo del ácaro también promueven el incremento de hembras (Rasmy et al., 2011); quizás las diferencias que se observaron entre primavera de 2012 con los otros dos ciclos de producción se atribuyan a un incremento en éstas. Otro factor ambiental que también puede afectar el crecimiento poblacional del ácaro es la fertilización de la planta (Chow et al., 2009; Flores et al., 2011).
La respuesta diferencial en el desarrollo poblacional del ácaro, también pudo estar afectada por mecanismos de defensa de la planta como barreras físicas o productos metabólicos. El contenido de fenoles totales medido en algunos de los cultivares (Figura 3), elegidos por sus valores medios y extremos en incidencia de ácaros (Figura 1) también mostraron variación natural; se pudo detectar la tendencia a ser mayores en cultivares con menor incidencia de araña roja.
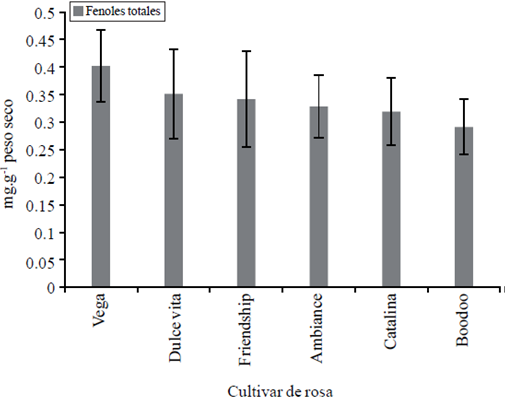
Figura 3 Contenido de fenoles totales en hojas fotosintéticamente activas de tallos florales de rosa determinado en las etapas fenológicas arrocillo, canica y punto de corte. Sobre las barras se indica la desviación estándar del error.
Otros autores han observado que los compuestos fenólicos pueden estar de forma constitutiva en las plantas o sintetizados en respuesta a la infección de la plaga (Gardner et al., 1999), donde se acumulan en mayor cantidad y más rápidamente en especies resistentes al ácaro, con la presencia de algún patógeno (Khatun et al., 2009), en hojas sanas (Coruh y Ercisli, 2010), o incluso por el manejo agronómico y variables ambientales presentes; su presencia también representan una ganancia evolutiva para retrasar el daño en condiciones ambientales adversas a la planta (Gardner et al., 1999).
Otros factores bioquímicos del metabolismo primario (moléculas antiestrés), y del secundario (terpenos, taninos y aceites esenciales) de carácter constitutivo e inducido, así como estructuras morfológicas (estomas, ceras, glándulas, grosor de hoja, etc.) también pueden influir en la atracción o repelencia del ácaro (Graham et al., 2008; Mumm y Dicke, 2010; Flores et al., 2011), todos dentro de una compleja relación entre parasito y huésped que debe explorarse individualmente (Hernández et al., 2002; Vivanco et al., 2005). Todos estos elementos pueden ser criterios complementarios en el manejo integral del cultivo y en mejoramiento genético (Chow et al., 2009). Recientes estudios reportan, además del manejo ambiental, el uso de compuestos volátiles de hojas de plantas como Cleome gynandra L./Gynandropsis gynandra (L.) Briq.), la cual en co-cultivo con Rosa x hybrida para producción de flor de corte, sin afectación en su rendimiento y calidad, ha reducido sustancialmente las poblaciones de T. urticae (Nyalala et al., 2013).
Variables asociadas a calidad del tallo floral
Entre cultivares se observaron diferencias altamente significativas (α= 0.01) en todas las variables asociadas a calidad del tallo floral, las cuales reflejan directamente la variabilidad fenotípica existente. Las características con mayor amplitud de variación entre cultivares fueron número de pétalos, área foliar, longitud de tallo, tallos productivos, tallos ciegos, número de foliolos y diámetro de tallo (NP, AF, LT, TP, TC, NF, DT; Cuadro 2). El efecto ambiental asociado a los tres ciclos de producción también repercutió en el desarrollo de los cultivares al observarse diferencias altamente significativas en todas las variables evaluadas, excepto en AF. La interacción significativa C x CU que se observó en todas las variables, excepto en las de contenido de clorofila, sugieren una respuesta diferencial de los cultivares a los factores ambientales predominantes en los tres ciclos de crecimiento y desarrollo; en el cv. bella perla, las temperaturas más bajas que se observaron en otoño de 2010 en relación a los otros dos ciclos, originaron un lento crecimiento del tallo floral y hubo un mayor número de tallos cortos y sin flor (tallos ciegos), también probablemente afectado por el fotoperiodo. El ciclo fenológico promedio a punto de corte en todos los cultivares también se extendió en 20 días.
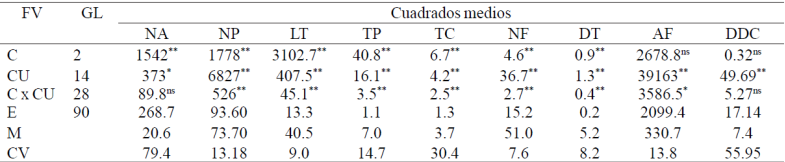
FV= fuente de variación; GL= grados de libertad; C= ciclos; CU= cultivares; E= error; M= media; C.V= coeficiente de variación (%). NA= número de ácaros; NP= número de pétalos; LT= longitud de tallo; TP= tallos productivos; TC= tallos ciegos; NF= número de foliolos; DT= diámetro de tallo; AF= área foliar; DDC=diferencia de clorofila. *= significativo; **= altamente significativo; ns= no significativo.
Cuadro 2 Cuadrados medios y significancia estadística de los valores de F de tres experimentos establecidos para la evaluación de cultivares de rosa.
Las variaciones en los contenidos de clorofila entre cultivares no fueron significativas; este hecho podría considerarse como el efecto varietal menos variable que influye en la tonalidad del color de las hojas, pero se observó una mayor dispersión en tejido con daño por ácaros, lo que remarca la disminución de la actividad fotosintética en determinados cultivares. Resultados similares también se observaron en hojas de café (coffea sp; Neves et al., 2006), también con reportes de síntesis de productos metabólicos.
Ácaros vs variables fisiológicas
El crecimiento poblacional del ácaro (NA) en todos los cultivares de rosa estuvo correlacionado negativa y significativamente con algunas variables de calidad del tallo floral, como longitud de tallo, diámetro de tallo y área foliar (LT, DT y AF; Cuadro 3). El NA con los contenidos de clorofila en las hojas también mostraron una tendencia similar.

NA= número de ácaros por planta; NP= número de pétalos; LT= longitud del tallo floral; TP= tallos productivos; TC= tallos ciegos; NF= número de foliolos; DT= diámetro de tallo floral; AF= área foliar; CSD= clorofila sin daño; CCD= clorofila con daño; * =significativo (p= 0.05); **= altamente significativo (p= 0.01).
Cuadro 3 Coeficientes de correlación entre las variables evaluadas.
Golawska et al. (2010), Calatayud et al. (2008), Neves et al. (2006) y Stone et al. (2001) han observado una disminución significativa en los valores de las variables fisiológicas de la planta al incrementarse poblacionalmente la plaga, con efectos directos en la calidad comercial del tallo floral (Tjosvold y Chaney, 2001). La interrupción en el desarrollo de la estructura floral aumenta con el incremento poblacional de T. urticae, aunque no significativamente; probablemente los factores edafoclimáticos y su interacción con el tipo de cultivar tengan un mayor efecto sobre este tipo de respuestas.
Análisis de conglomerados
Los datos para los 15 cultivares de rosa y para las variables evaluadas fueron sometidos a un análisis multivariado con el método de media aritmética no ponderado (UPGMA Method). En éste, al considerar una distancia promedio de ligamiento de 4.17 se identificaron cinco grupos (Cuadro 4): en el grupo 1 sólo se observó al cv. Friendship (FS), mejorado en Francia, de flor bicolor en rojo y crema, y con nivel de tolerancia a araña roja estadísticamente igual al mostrado por los mejores cultivares, como catalina, voodoo y freedom (Figuras 1 y 3). El segundo grupo contiene a los cv. catalina y voodoo los cuales se caracterizaron por hospedar el mayor número de ácaros (NA, Figura 1). El cv. catalina fue obtenido por hibridadores en Nueva Zelanda y en términos de distancia geográfica representa al lugar de mayor aislamiento con respecto a los lugares donde se desarrollaron los demás cultivares estudiados, que fue en países de Europa (Cuadro 1). El cv. voodoo, desarrollado en Holanda, está reportado en California, Estados Unidos de Norteamérica como resistente a enfermedades (Anónimo 3, 1978). El tercer grupo contiene a los cv. poison, freedom y grand gala, originarios de Holanda, Alemania y Francia, respectivamente; en los dos últimos, de flor roja, no se encontró información sobre sus progenitores, por lo que se desconoce si ambos y poison, de flor púrpura, están emparentados.
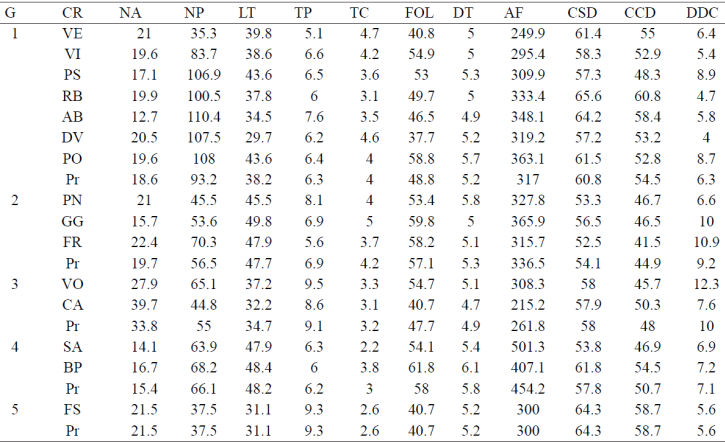
Cuadro 4 Medias de variables evaluadas de 15 cultivares de rosa, agrupadas por análisis de conglomerados.
El cuarto grupo se integró con samourai y bella perla, desarrollados en Francia por diferentes empresas y contrastantes en progenitores y color de flor. Ambos conformaron el grupo de cultivares con la menor tolerancia al daño causado por ácaros (Figura 1). El quinto grupo representa casi al 50% de los cultivares evaluados; vega, dulce vita, visión, polar star, polo, royal baccara y ambiance, son contrastantes en cinco tonos de color de flor. Los cv. polar star y polo, de flor blanca, conformaron un subgrupo pero como se desconocen sus progenitores, no es posible inferir si hay parentesco entre ellos. Con base en la percepción de los productores en el área de estudio el cultivar polo es uno de los más susceptibles a plagas y enfermedades y éste y polar star fueron desarrollados por la misma empresa en Alemania.
Los 15 cultivares analizados en el dendograma de la Figura 4 pertenecen al grupo de híbridos de tea, considerados como los más representativos dentro de la categoría de las rosas modernas, obtenidos por hibridación de rosas de los grupos tea e híbridos perpetuos de la categoría de rosas viejas. Las rosas del presente estudio, estrictamente clasificadas como híbridos de tea, representan una mezcla compleja reducidamente ubicada en un margen estrecho de una amplia variación genética de más de 100 cultivares distribuidos en 13 grupos de las categorías de rosas modernas y viejas (Martin et al., 2001). Los componentes principales (CP) 1, 2 y 3 explicaron el 72.5% de la variación total original; casi un 50% fue representada en el biplot usando los CP 1 y 2 (Figura 5).
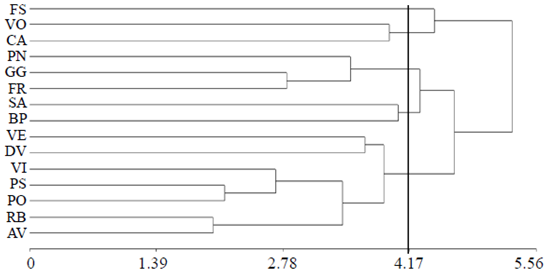
Figura 4 Dendograma para 15 cultivares de rosa obtenido por el método de ligamiento promedio (Método UPGMA). FS= friendship; VO= voodoo; CA= catalina; PN= poison; GG= grand gala; FR= freedom; SA= samourai; BP= bella perla; VE= vega; DV= dulce vita; VI= visión; PS= polo star; PO= polo; RB= royal baccara; AB= ambiance.

Según Sánchez (1995) y González et al. (2010) estos porcentajes sugieren confiabilidad en la interpretación de las correlaciones aproximadas que se pueden detectar entre los cultivares y las variables evaluadas en el presente estudio. En el biplot se observaron tres agrupaciones de cultivares por valores similares identificados por su proximidad dentro de un mismo cuadrante. El primer grupo incluyó a los cultivares freedom, poison y grand gala (FR, PN y GG); el segundo grupo lo constituyeron los cultivares polar star, samourai y bella perla (PS, SA, y BP); el grupo tres incluyó a vega, visión, dulce vita, ambiance y royal baccara (VE. VI, DV, AB y RB). Los cultivares voodoo, catalina y friendship (VO, CA y FS) disimilares en sus puntuaciones en ambos CP 1 y 2, no se incluyeron en los grupos anteriores.
Análisis de componentes principales
El CP1 se explicó principalmente por la longitud del tallo (lt) y por el número de foliolos (fol); el CP2 se relacionó esencialmente con número de ácaros (na) y tallos productivos (tp); y en el CP3 las mayores puntuaciones se registraron en número de pétalos (np) y tallos ciegos (TC). Los cultivares CA, AM, SA, FR y GG tuvieron las mayores puntuaciones en la CP1; en la CP2 se observó que CA y AB tuvieron la mayor dispersión en la gráfica del biplot y en el CP3 esta misma situación de detectó con BO y PO (Figura 5).
En este biplot también se puede observar que CA, BO, FS y VE tuvieron el mayor número de ácaros (NA) y tallos productivos. SA, GG, FR, BP, PN, PO y PS fueron los más sobresalientes en longitud de tallo (lt), número de foliolos (fo), área foliar (af), diámetro de tallo (dt), diferencia de clorofila (ddc), número de pétalos (np) y tallos ciegos (tc). RB, AB, DV y VI se agruparon principalmente por sus contenidos de clorofila con daño (ccd) y sin daño (csd).
Conclusión
El desarrollo poblacional de T. urtucae en rosa, además de ser afectado por factores ambientales como humedad y temperatura, también lo es por la respuesta diferencial de cultivares en sus mecanismos naturales complejos de defensa que incluyen productos metabólicos de la planta, los cuales se reflejaron en la calidad del tallo floral medido en diez diferentes parámetros. Además, la variación en el crecimiento poblacional del ácaro entre cultivares denota que aunque las rosas modernas son producto de un margen estrecho de más de 100 especies, éstas conservan variabilidad genética, posiblemente como resultado de las cruzas complejas que les han dado origen y que pudiera ser explotada en programas de mejoramiento. La distancia geográfica entre los orígenes de algunos cultivares coincidió con la respuesta diferencial más extrema de cultivares al desarrollo del ácaro.

