











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El tizón tardío de la papa, causado por el oomiceto Phytophthora infestans Sulc., es uno de los principales problemas para la producción de papa en el mundo (Haverkort et al., 2009). En México, el centro y noreste son las regiones más afectadas por la enfermedad, en donde se hacen al menos dos aplicaciones de fungicidas por semana durante la época de mayor incidencia de la enfermedad (Rubio et al., 2000).
Desde la devastación de los campos de papa por el tizón tardío ocurrida en Irlanda en 1845, se intensificaron los esfuerzos por generar variedades resistentes. Inicialmente, los programas de mejoramiento genético de varios países utilizaron los genes mayores (genes R) provenientes de la especie silvestre Mexicana Solanum demissum como fuente de resistencia, sin embargo, muchas variedades cuya resistencia vertical se basaba en genes R se volvieron susceptibles en poco tiempo, por lo que se pensó que se deberían generar variedades con resistencia horizontal fundamentada en genes menores, con lo cual se esperaba obtener una resistencia duradera (Turkensteen, 1993; Landeo et al., 1995).
La resistencia vertical también es conocida como resistencia cualitativa, resistencia específica o resistencia total y la resistencia horizontal como resistencia cuantitativa, resistencia general, resistencia parcial o resistencia de campo. Sin embargo, en la actualidad se sabe que los genes R están involucrados en todos los tipos de resistencia por lo que se está tratando de generar variedades con una combinación de varios genes mayores y menores, a lo cual se le ha llamado construir pirámides de genes (Adillah et al., 2010; Kim et al., 2012; Zhu et al., 2013). Esta estrategia se desarrolla actualmente con la ayuda de la ingeniería genética, pero algunas variedades viejas que poseen una combinación de genes mayores y menores, las cuales fueron generadas por métodos tradicionales en México y otros países, han demostrado que tienen una resistencia duradera (Grünwald et al., 2002; Rietman et al., 2012).
Desde que Van der Plank (1963) describió los conceptos de resistencia horizontal y vertical, ha habido un interés constante en diferenciar la resistencia horizontal y vertical. La pendiente de la curva de infección, la etapa de desarrollo de las plantas en la que aparecen las primeras lesiones, la presencia de esporangios en las lesiones, y la reacción de hipersensibilidad en las hojas son algunos de los parámetros que se han utilizado para diferenciar los dos tipos de resistencia. De estos parámetros, los dos primeros han sido calculados a partir del área relativa bajo la curva de infección y han servido para clasificar un gran número de clones de acuerdo a su tipo de resistencia (Marhadour et al., 2013). En trabajos previos realizados bajo condiciones controladas en cámaras de crecimiento, se comparó la respuesta a la inoculación con un aislamiento de P. infestans en genotipos con y sin genes R (Rubio et al., 2005; Rubio et al., 2006). En estos trabajos se observaron los mismos mecanismos de resistencia en ambos grupos de clones, sin embargo, estos resultados deben validarse bajo condiciones naturales de infección de campo en un lugar en donde exista la mayor biodiversidad de razas de P. infestans, como es el Valle de Toluca, el cual es considerado el centro de origen de este oomiceto (Goodwin et al., 1992; Grünwald et al., 2001).
La mayor parte de las variedades de papa liberadas en México por el INIFAP poseen una combinación de genes mayores y menores que les confieren resistencia contra P. infestans (Flores y Cadena, 1996; Grünwald et al., 2002; Rubio et al., 2005). Su evaluación se ha realizado bajo condiciones de campo en el Valle de Toluca, lo cual ha permitido seleccionar algunos cultivares que tienen resistencia duradera, sin embargo, no se han comparado con genotipos que solamente tienen genes menores. El objetivo de este trabajo es realizar esta comparación bajo las condiciones de campo en el Valle de Toluca, con lo cual se espera contribuir al entendimiento de los dos tipos de resistencia y apoyar en la generación de variedades de papa que tengan resistencia duradera contra el tizón tardío. Este conocimiento es de gran importancia en la época actual, en la cual diversos investigadores han emitido opiniones controversiales que favorecen la utilización de pirámides de genes mayores, genes menores o la combinación de ambos (Darsow, 2014).
Materiales y métodos
Durante 3 años consecutivos (2008-2010) se establecieron experimentos de campo en el Sitio Experimental del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) en Metepec, localizado dentro del Valle de Toluca. En cada ciclo agrícola se compararon 10 clones de papa libres de genes R (resistencia horizontal) contra 10 genotipos con genes R (resistencia vertical). Los primeros fueron generados por el Centro Internacional de la Papa (CIP) y forman parte del grupo conocido como Población B, cuya resistencia horizontal contra P. infestans proviene de materiales avanzados libres de genes R, los cuales a su vez fueron derivados de genotipos con genes mayores (R) y menores (Landeo, 1995). Los clones de la población B utilizados en el presente estudio ya habían sido evaluados en años anteriores en el Sitio Experimental de Metepec del INIFAP y fueron seleccionados por su resistencia contra el tizón tardío, por su rendimiento y en general por su buena adaptación a las condiciones de campo en las que se sembraron (datos no publicados). En este proceso de selección se eliminaron de este estudio los clones que tienen un ciclo de madurez muy tardío, esto se hizo para evitar el sesgo que puede ocasionar la relación que existe entre el nivel de resistencia con la duración del ciclo vegetativo de los genotipos de papa (Bradshaw et al., 2004; Darsow, 2014).
Los genotipos de la población A fueron generados por el INIFAP y su resistencia se basa en la combinación de genes mayores y menores. Entre esta población existen 6 variedades que fueron liberadas previamente y 4 clones que fueron seleccionados por su buena adaptación a las condiciones en las que se cultiva la papa en el Valle de Toluca. En estos materiales, la resistencia contra el tizón tardío de la papa proviene de la especie silvestre Solanum demissum, la cual es originaria de México (Fores et al., 1996; Grunwald et al., 2002).
La época de desarrollo del cultivo fue durante el periodo de lluvia en los meses de junio a septiembre, por lo que no se aplicó ningún riego. El diseño experimental fue de bloques al azar con 10 repeticiones. En cada repetición se sembró una planta de cada uno de los 20 genotipos resistentes al tizón tardío (10 de la población A y 10 de la población B) con 1 planta de una variedad susceptible (Alpha) intercalada cada 3 plantas de los genotipos resistentes. Esto se hizo para asegurar una distribución uniforme del tizón tardío en el lote experimental. La distancia entre surcos fue de 92 cm y entre plantas de 30 cm. Durante el desarrollo del cultivo no se aplicó ningún fungicida ni se inocularon las plantas, por lo que la infección de las plantas con P. infestans ocurrió de manera natural. En cada planta se observó la fecha en la que apareció la primera lesión provocada por el tizón tardío y posteriormente se hicieron observaciones semanales sobre el progreso de la enfermedad en forma de porcentaje del área foliar infectada. El porcentaje de las lesiones cubiertas por esporas se estimó visualmente durante la época más propicia para la esporulación, la cual estuvo asociada principalmente con la época de mayor frecuencia y cantidad de lluvia. En algunos años se hicieron hasta 3 evaluaciones de esporulación debido a la incertidumbre sobre cuando ocurriría la máxima, pero solo se utilizaron los datos que reportaban los valores máximos.
Cada una de las 5 variables analizadas en este estudio [área relativa bajo la curva de infección (ARBCI), pendiente (pendiente de la regresión lineal: tiempo vs (%) de infección), (%) de las lesiones foliares cubiertas por esporangios, número de días después de la emergencia de las plantas cuando se observó la primera infección foliar y el rendimiento de tubérculos (g/planta)] se analizaron estadísticamente utilizando PROC GLM de SAS v9.2. En el análisis de varianza (ANVA) se probó si había diferencia entre genotipos (10 de la población A, 10 de la población B y la variedad susceptible Alpha), entre repeticiones en el espacio (10 repeticiones en cada experimento) y entre repeticiones en el tiempo (3 años). El análisis estadístico de este tipo de experimentos es descrito en Steel and Torrie (1980). El ANVA reportó diferencias significativas entre genotipos, por lo que se procedió a comparar las medias individuales por medio de una prueba de Tukey y las medias entre poblaciones utilizando una prueba de contrastes.
Resultados y discusión
El análisis de varianza demostró que casi todas las fuentes de variación (genotipo, repetición y año) fueron estadísticamente significativas (p≤ 0.05) en las variables dependientes ARBCI, pendiente, (%) de esporulación, primera infección foliar y rendimiento de tubérculos, con excepción del efecto de repeticiones y años en el rendimiento. Estos resultados indican que las diferencias climáticas entre años y las diferencias entre los genotipos tuvieron influencia sobre las variables que se utilizaron para evaluar la resistencia contra el tizón tardío en las plantas.
El progreso de la enfermedad durante el desarrollo de las plantas en los tres años de estudio se presenta en la Figura 1, en la cual se puede apreciar que en los años 2008 y 2010, el porcentaje del área foliar infectada fue mayor que en 2009. La cantidad total de precipitación ocurrida durante el periodo de desarrollo de las plantas (junio-septiembre) fue de 657, 684 y 669 mm en los años 2008, 2009 y 2010 respectivamente. Las diferencias en precipitación total durante los 3 años fueron pequeñas, sin embargo en el año 2009 hubo un periodo de baja precipitación entre los días 49 y 63 después de que las plantas emergieron, lo cual hizo que la infección fuera menor en este año. En la Figura 1 también se puede observar una gran diferencia de los genotipos que poseen algún tipo de resistencia (poblaciones A y B) respecto a la variedad Alpha, la cual es susceptible. Sin embargo, la diferencia entre las poblaciones A y B es muy pequeña pero significativa al final del ciclo de desarrollo de las plantas, sobre todo en los años con mayor severidad del tizón tardío (2008 y 2010).
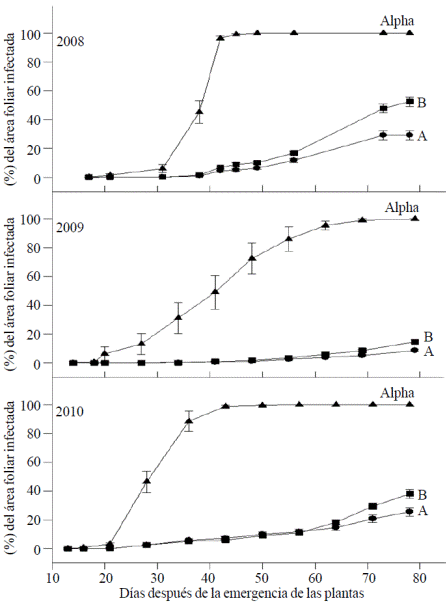
Figura. 1 Progreso de la infección foliar por P. infestans en genotipos de papa de las poblaciones A, B y en la variedad Alpha en pruebas de campo realizadas en el Valle de Toluca durante 3 años.
Estas curvas del avance de la enfermedad están relacionadas con la forma en que actúan los dos tipos de resistencia. De acuerdo a las observaciones de diversos investigadores (Van der Plank, 1963; Nelson, 1978; Marhadour et al., 2013), se ha establecido que la resistencia horizontal, a base de genes menores, se refleja en una baja pendiente de la curva de infección sin que se presente un incremento brusco que indique que la resistencia ha sido vencida, como ocurre en los genotipos que tienen únicamente genes R. La forma de las curvas de desarrollo de la enfermedad (Figura 1) sugieren que la resistencia de ambas poblaciones no es total y que los mecanismos de defensa en ambas poblaciones son parcialmente vencidos, pero los de la población A son más efectivos que los de la población B para disminuir el desarrollo de la enfermedad. Estas diferencias se deben a que la resistencia de la población B se basa en genes menores (Landeo, 1997) y en la población B existe una combinación de genes mayores y menores (Flores y Cadena, 1996; Grünwald et al., 2002; Rubio et al., 2005). Este hecho es apoyado por las diferencias significativas entre las medias de ARBCI, de la pendiente y del tiempo en que se aprecia la primer infección foliar en las poblaciones A y B (Cuadro 1).
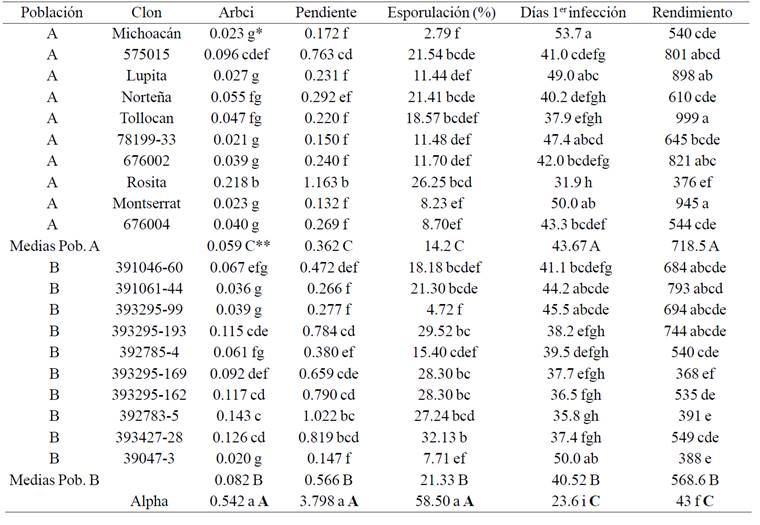
*Prueba deTukey para comparar las medias de los genotipos. Medias con diferentes letras minúsculas indican diferencias significativas (p≤ 0.05). **Prueba de contrastes para comparar las medias entre poblaciones y con la variedad Alpha. Medias con diferentes letras mayúsculas indican diferencias significativas (p≤ 0.05).
Cuadro 1 Área relativa bajo la curva de infección (ARBCI), pendiente (pendiente de la regresión lineal: tiempo vs % de infección), % de las lesiones foliares cubiertas por esporangios, número de días después de la emergencia de las plantas cuando se observó la primera infección foliar y rendimiento de tubérculos (g/planta) de 10 genotipos de la población A, 10 genotipos de la Población B y una variedad susceptible (Alpha). Los números son medias de 3 años y 10 repeticiones por año.
Desde otro punto de vista, la forma parecida en que se desarrolla la enfermedad de las poblaciones A y B (Figura 1), indica la dificultad de separar la resistencia horizontal y vertical debido a la asociación entre los factores que controlan la resistencia. En relación con esto, Stewart et al. (2003) demostraron que hay un efecto residual de la resistencia después de que los genes R han sido vencidos por razas compatibles de P. infestans. En base a la asociación entre genes R y QTLs, a los que se les atribuye la resistencia vertical y horizontal respectivamente, se ha propuesto que la resistencia horizontal puede ser una expresión de varios genes, incluyendo alelos de genes R, que tienen un efecto complementario sobre la activación de los mecanismos de defensa (Gerbhardt y Valkonene, 2001). En este esquema, la resistencia vertical es el resultado de una coordinación eficiente entre los genes Ry aquellos otros genes secundarios que intervienen en la resistencia. Esta propuesta es apoyada por la teoría sobre la cantidad de sensores que se necesitan para iniciar las reacciones de resistencia, la cual se basa en la diferente efectividad que han mostrado los genes R1 y R2 para activar las reacciones de hipersensibilidad (Vleeshouwers et al., 2000).
En este caso, un incremento en los receptores puede aumentar la sensibilidad para detectar y controlar el patógeno, por lo tanto, la variación alelica puede regular los receptores de la planta y consecuentemente afectar la respuesta de defensa. Por el lado del patógeno, Vleeshouwers et al. (2011) han descrito los mecanismos que P. infestans utiliza para que las substancias que secreta al invadir a su hospedero, no sean detectadas por las proteínas que son producidas en la planta por efecto de los genes R. En base a estos mecanismos de resistencia, se asume que los clones de la población A utilizados en el presente estudio, tienen varios genes mayores y menores de resistencia, cuyo efecto aditivo los hace más resistentes que los clones de la población B, en los que supuestamente no existen genes R, pero en los cuales pueden existir sus alelos que los hacen reaccionar de manera similar pero menos efectiva que los genotipos de la población A.
Puesto que el análisis de varianza indicó diferencias significativas entre genotipos, el siguiente paso fue la comparación de sus medias. En el cuadro 1 se presentan las medias de las 5 variables por genotipo y por población. En este cuadro se puede apreciar que de acuerdo al área relativa bajo la curva de infección (ARBCI) y de la pendiente, todos los clones de ambas poblaciones son más resistentes al tizón tardío que la variedad Alpha. En ambas poblaciones (A y B) existe un amplio rango de niveles de resistencia, siendo los genotipos 78199-33, Montserrat, Michoacán, Lupita, 676002, 676004, Tollocan y Norteña los más resistentes de la población A. En la población B los clones más resistentes son: 3947-3, 391061-44 y 393295-99. La comparación de las medias de las 5 variables por población (Cuadro 1) permite establecer una amplia superioridad de los genotipos que poseen algún tipo de resistencia (poblaciones A y B) sobre la variedad Alpha, la cual es susceptible. Sin embargo, las diferencias entre las poblaciones A y B son estrechas, pero significativas e indican mayor resistencia y rendimiento de los genotipos de la población A que los genotipos de la población B. El mayor rendimiento de la población A se asocia a la menor infección en el follaje, por lo que existe una mayor cantidad de fotosintatos que son producidos en la parte aérea de la planta y trasladados hacia los tubérculos.
La mayor resistencia de la población A no puede ser atribuida a la duración del ciclo vegetativo de las plantas, ya que en las dos poblaciones no hubo genotipos que se consideren muy tardíos. Es de conocimiento general que entre mayor es la duración del ciclo vegetativo de la planta mayor es la resistencia contra el tizón tardío de la papa (Bradshaw et al., 2004; Darsow, 2014). Esta relación es más clara en genotipos con resistencia horizontal, sin embargo, estudios moleculares recientes han demostrado que pueden existir algunos QTLs, que confieren resistencia, pero que no están ligados a los genes que confieren una madurez tardía de las plantas (Danan et al., 2011). Desafortunadamente no se han hecho estudios que permitan definir los genes R y QTLs que poseen los materiales de la población A utilizados en este estudio. Esta información, junto con técnicas modernas de ingeniería genética, podría servir para hacer más eficiente el proceso de generación y selección de genotipos con alto nivel de resistencia que no sea vencida en un corto plazo.
En el presente estudio se observó que la población B presentó mayor porcentaje de esporulación que la población A (Cuadro 1). La menor esporulación observada en la población A se puede explicar si se considera que la resistencia está basada en una combinación de genes mayores y menores; por lo tanto su efecto de resistencia es mayor que cuando solo se tienen genes menores. Esta explicación se apoya en los conceptos de Van der Plank (1963), quien mencionó que los genotipos con resistencia vertical muestran una reducción inicial de la esporulación debido a la acción de los genes R, pero una vez que la resistencia es vencida, la cantidad de inoculo aumenta rápidamente. Por lo contrario, los genotipos con resistencia horizontal se caracterizan por una disminución de la esporulación durante todo su ciclo debido a la acción conjunta de varios genes menores (Niederhauser, 1961; James and Fry, 1983). Por lo tanto, al combinar ambos tipos de resistencia es de esperarse una menor esporulación en la población A.
Observaciones adicionales hechas durante el desarrollo de las plantas indican que todos los genotipos en este estudio presentaron en alguna etapa de su desarrollo reacciones de hipersensibilidad (RH), la cual consiste en pequeñas aéreas necróticas que aparecen en las hojas como resultado de la muerte programada de las células para evitar la expansión de la infección. Estos resultados están de acuerdo con los trabajos de Gees and Hohl (1988), quienes demostraron que la RH puede presentarse indistintamente en plantas de papa con resistencia vertical y horizontal. Otros investigadores (Vleeshowers et al., 2000) observaron que la RH no es exclusiva de genotipos con genes R sino que también puede presentarse en especies silvestres de papa con varios niveles de resistencia contra el tizón tardío, sin embargo, en este caso, la velocidad y efectividad de la RH para controlar la infección fue asociada con los niveles de resistencia de los diferentes genotipos. Por lo tanto, la RH establecida inicialmente por Van der Plank (1963) para distinguir la resistencia horizontal y vertical no puede ser considerada válida en todos los casos.
Es importante señalar que algunos progenitores de la población B provienen de genotipos con genes R (Landeo, 1997). De acuerdo con este investigador, el proceso de mejoramiento genético para seleccionar genotipos libres de genes R se basó en la ausencia de la RH cuando los clones fueron inoculados con la raza 0 de P. infestans. Sin embargo, se ha demostrado que algunos genotipos sin genes R pueden mostrar la RH con la raza 0 (Gees and Hohl, 1988; Vleeshowers et al., 2000). Por otro lado, se desconoce cuál podría ser la reacción de las plantas que poseen genes R desconocidos. La incertidumbre de la prueba para separar genotipos con y sin genes R utilizando la reacción a la raza 0 de P. infestans, el comportamiento parecido de las curvas de desarrollo de la enfermedad de las poblaciones A y B en este estudio y la asociación entre genes R y QTLs, abren la posibilidad de que los clones de la población B tengan genes R.
Los resultados en el presente trabajo realizado bajo condiciones de campo, concuerdan con estudios previos realizados bajo condiciones controladas en cámaras de crecimiento (Rubio et al., 2005; Rubio et al., 2006). En dichos estudios se compararon dos variedades de la población A y dos clones de la B inoculados con una raza de P. infestans, y los resultados indicaron mayor resistencia en los genotipos de la población B. Es importante hacer notar que las pruebas realizadas en el presente trabajo con más genotipos y repetidas durante 3 años bajo condiciones de campo en el Valle de Toluca, dan mayor certeza a los resultados. La gran biodiversidad de razas de P. infestans que se encuentra en el Valle de Toluca da seguridad al proceso de evaluación de la resistencia. Los resultados de Blackburn et al. (2007) demuestran que el uso de razas complejas de P. infestans permiten distinguir genotipos con resistencia horizontal y vertical. En nuestro estudio, los materiales fueron sometidos a condiciones de campo en un lugar en el que existe una gran biodiversidad de razas de P. infestans (Goodwin et al., 1992; Grünwald et al., 2001).
La durabilidad de la resistencia solamente puede ser evaluada cuando una variedad es sembrada por varios años en un lugar en donde exista una gran biodiversidad de razas de P. infestans, como lo es el Valle de Toluca. La durabilidad de la resistencia de las variedades de la población A (Tollocan, Montserrat, Norteña, Michoacán y Rosita) utilizadas en el presente estudio, ha sido demostrada previamente bajo las condiciones del Valle de Toluca (Grunwald et al., 2002). Esto confirma que el proceso de mejoramiento genético del programa de papa del INIFAP ha sido, hasta el momento, un sistema adecuado para seleccionar genotipos resistentes y estables.
Conclusión
Los resultados de este estudio demuestran que los genotipos de papa de la población A (con resistencia vertical) tienen mayor resistencia que los de la población B (con resistencia horizontal). La población A mostró superioridad en la resistencia evaluada por 5 variables: área relativa bajo la curva de infección (ARBCI), pendiente (pendiente de la regresión lineal: tiempo vs % de infección), % de las lesiones foliares cubiertas por esporangios, número de días después de la emergencia de las plantas cuando se observó la primera infección foliar y el rendimiento de tubérculos. La combinación de genes mayores y menores de la población A, ha permitido crear genotipos con alto nivel de resistencia, que además puede ser duradera. La selección de materiales bajo las condiciones naturales de infección en el Valle de Toluca, en donde existe la mayor biodiversidad de razas de P. infestans, ha sido una estrategia adecuada para seleccionar variedades de papa resistentes al tizón tardío.

