










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.7 no.3 Texcoco abr./may. 2016
Articles
Prohexadione calcium (P-CA): a viable alternative hormonal habanero hot pepper
1 Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1923, Buenavista. Saltillo. Coahuila C. P. 25315. México. (joakinmzcs@gmail.com; marioe. vazquez@hotmail.com; azermenog@hotmail.com).
The habanero hot pepper planting in Mexico increased in recent years. The demand for this vegetable worldwide requires the application of new techniques to improve their performance and quality. One option is the use of bioregulators. The aim of this study was to determine the effect of retardant prohexadione calcium (P-Ca) on vegetative growth, yield and antioxidant levels in habanero hot pepper variety Jaguar. The experiment was a completely randomized design with 3 replicates per treatment during 2014 in UAAAN in Saltillo, Coahuila, under greenhouse conditions. Concentrations of P-Ca were: 0 (water control), 50, 100 and 150 mg L-1 in 1 and 2 applications. The first took place when the plants showed the first floral primordia and 15 days after the second was performed. The results indicate that P-Ca decreased to 52% and 30.5% in height and stem diameter respectively. Most P-Ca treatments increased the number of f lowers, fruits per plant and yield; the content of antioxidants in ripe fruits. P-Ca 150 mg L-1 was the treatment that caused the highest content of capsaicin, total carotenes and vitamin C in fruits. It is considered that P-Ca has potential for use as an alternative to improve performance and quality of habanero hot pepper.
Keywords: Capsicum chinense Jacq.; capsaicin; performance; total carotenoids; vitamin
La siembra de chile habanero en México aumentó en años recientes. La demanda de esta hortaliza a nivel mundial exige la aplicación de nuevas técnicas que permitan mejorar su rendimiento y calidad. Una opción es el uso de bioreguladores. El objetivo del presente estudio fue determinar el efecto del retardante prohexadiona de calcio (P-Ca) sobre el desarrollo vegetativo, rendimiento y niveles de antioxidantes en chile habanero variedad Jaguar. El experimento se estableció con un diseño completamente al azar con 3 repeticiones por tratamiento durante el año 2014 en la UAAAN en Saltillo, Coahuila, en condiciones de invernadero. Las concentraciones de P-Ca fueron: 0 (testigo agua), 50, 100 y 150 mg L-1 en 1 y 2 aplicaciones. La primera se realizó cuando las plantas mostraron el primer primordio floral y 15 días después se realizó la segunda. Los resultados indican que P-Ca redujo hasta un 52% y 30.5% en la altura y diámetro del tallo respectivamente. La mayoría de los tratamientos con P-Ca aumentaron el número de flores, frutos por planta y rendimiento; el contenido de antioxidantes en frutos maduros. P-Ca a 150 mg L-1 fue el tratamiento que provocó el mayor contenido de capsaicina, carotenos totales y vitamina C en los frutos. Se considera que P-Ca tiene potencial para utilizarse como alternativa para mejorar el rendimiento y calidad del chile habanero.
Palabras claves: Capsicum chinense Jacq.; capsaicina; carotenoides totales; rendimiento; vitamina C
Introduction
Chilies are known for their rich in micronutrients and bioactive compounds (phenolic compounds), so its consumption has been clearly recognized as an important factor for good health (Rodriguez et al., 2012). Antioxidants in peppers are numerous and include, ascorbic acid, flavonoids, capsaicinoids, and a wide variety of phenolic acids (Howard et al., 2000). Its pungency is related to the concentration of five compounds: capsaicin, nordihydrocapsaicin, dehydrocapsaicin, homocapsaicin and homodihydrocapsaicin, which are classified as capsaicinoids (Krajewska, 1988; Antonious et al., 2006, 2009). Vazquez-Flota et al. (2007) mention that there are 20 different types of these compounds in chiles. Dehydrocapsaicin capsaicin and capsaicinoids are the dominant detected in different types of hot peppers (Antonius et al., 2006).
The colors yellow, orange and red capsicum fruits are caused by carotenoids produced during ripening, which have immense nutritional value as provitamin A and antioxidants. The β-carotene are precursors of orange and red pigments in chili peppers. Chilies with high concentration of β-carotene proved to be rich in total carotenoids (Wall et al., 2001). Fresh chillies are excellent source of ascorbic acid, which is involved in various antioxidant processes of plants besides acting in the prevention of human chronic diseases, including certain cancers, coronary heart disease, arteriosclerosis and cataracts (Howard et al., 2000), making it a good alternative bioregulators use as stimulants to increase the antioxidant content in plants, which in turn, help reduce physiological stress and improve quality nutraceutical fruit.
In recent years the use of bio-oxidation promoters and tomato, broccoli and cabbage have proved to be an alternative to improve production and quality of the harvested product (Ramirez et al., 2009). The use of these substances has the advantage of producing effects are not permanent and therefore be modified according to the needs Horticulturist (Ramirez, 2003). P-Ca is a growth retardant currently used in fruit growing and is regarded as a promising alternative in modern horticulture (Ramirez et al., 2010c). P-Ca (3-oxide- 4-propionyl-5-oxo-3-cyclohexane-carboxylate) inhibits the biosynthesis of active gibberellins growth, reducing the longitudinal growth of shoots.
It has an effect in reducing vegetative growth in the plant, which stimulates a change in the translocation of assimilates and increased flower bud formation and therefore more fruit per plant (Ramirez et al., 2009). The use of P-Ca in fruit increases the levels of soluble solids, anthocyanins and aromas in the harvested product and substantially increases the total antioxidant capacity and concentration of lycopene in degrees of maturity; also it reduces levels of biologically active gibberellins and cytokinins increases in apical meristems in tomato fruit (Ramirez et al., 2010b; Ramirez et al., 2005). In jalapeno pepper capsaicin levels and increase yield per plant (Ramirez et al., 2009).
Therefore the aim of this research was to evaluate the effects of P-Ca on the phenotype, performance and antioxidant fruit quality of habanero hot pepper grow Jaguar and generate a technology for potential use prohexadione of Ca in the production of habanero hot pepper. The hypothesis was that P-Ca changes the phenotype of the plant and fruit quality in habanero hot pepper variety Jaguar.
Materials and methods
The experiment was established at the Universidad Autonoma Agraria Antonio Narro in Saltillo, Coahuila, Mexico, under greenhouse conditions, located at 25° 23' north latitude and 101° 01' latitude east, at an altitude of 1 743 meters. The habanero hot pepper seeds (Capsicum chinense Jacq.) Jaguar variety was used. The seeds were sown on May 30, 2014 in polystyrene trays of 200 cavities, using peat moss as a substrate mix premier. The transplant took place on July 11, 2014, when the plants had a height of 10 cm inside a greenhouse overhead type of metallic structure with side polycarbonate sheets and milky white plastic gauge (caliber 720) on the roof.
This was done in black plastic bags 20 L, substrate was used peat moss and perlite (70:30 v/v). The bags with vegetative material were arranged at a distance of 40 cm between plants and 70 cm between rows. When the plants had f lower primordia foliar application with P-Ca at doses of 0 (control- water), 50, 100 and 150 mg L-1 was performed. Asecond spray with P-Ca was carried out with the same concentrations 15 days after the first application. The variables evaluated were high growth rate and stem diameter, number of fruits, fruit length and yield per plant; capsaicin content, total carotenoids, vitamin C in ripe fruits. A completely randomized statistical design with 3 replicates per treatment was used. The results were analyzed with SAS statistical software version 9.1, for the analysis of variance and mean comparison by LSD (p≤ 0.05).
Horticultural assessment. The growth rate of the plant height and stem diameter was obtained when measured weekly during their life cycle. To measure the height, a tape was used with scale from 0 to 2 m, measuring from the stem base to the apex of the plant. The stem diameter was measured in its middle part with a vernier on a scale of 0 to 10 cm. The number of f lowers per plant was recorded in each bloom. The length and number of fruits and yield were determined in each of the four cuts and joined at the end. For heavy fruit Ohaus a scale model SP602, maximum capacity 600 grams with 0.1 grams approach was used.
Determination of antioxidants. The capsaicin content in fruits was determined by the method of Bennet and Kirby (1968). The 5 ripe fruits per repetition per treatment were made, lyophilized and macerated in mortar. A 1 g of sample was weighed and were added 10 ml of absolute ethanol and the mixture was stirred for 15 min. It was filtered on Whatman No. 1 and is gauged at 25 ml ethanol. The sample was transferred to a separatory funnel and 2.5 ml of buffer at pH 2.8 plus 0.5 ml of ethanol, 20.5 ml distilled water and 10 ml of solution adogen-toluene added. To 1 min the mixture was vigorously stirred. Subsequently, the absorbance of capsaicin was determined in the organic phase in a spectrophotometer (Thermo Electron Corporation Biomate 5) at a wavelength of 286 nm. A reading repetition was performed. To determine the concentration of capsaicin in the samples, a calibration curve was constructed with this antioxidant (Sigma, Co) in a range of 0 to 0.40 mg ml-1, dissolved in the solvents mentioned.
Determination of total carotenoid content in tissues was performed using the technique described by Thomas (1975), with some modifications. A 10 g of fresh fruit were weighed and 50 ml of acetone was added and 24 h was left cooling. In a mortar and triturated liquid was filtered in gauze through a separatory funnel, the sample through a cheesecloth with 20 ml acetone washed four times. Was added 20 ml of petroleum ether and 100 ml of distilled water and mixed gently by separating the upper container layer carotenoid turned add 20 ml of petroleum ether and 100 ml of distilled water eight to nine times and It was added 10 ml of 40% NaOH. Subsequently the sample with 50 ml of distilled water was washed to remove 40% NaOH used as indicator phenolphthalein and 20 ml of sodium sulfate was added to 10%. The mixture was filtered through cheesecloth containing anhydrous sodium sulfate and the final volume of the solution was taken. The carotenoid absorbance was determined in a spectrophotometer (Thermo Electron Corporation Biomate 5) at a wavelength of 454 nm using as blank petroleum ether. The total carotenoid content was determined by the following formula:
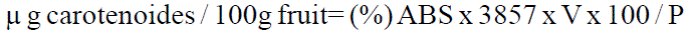
Where: (%) ABS= percent absorbance; 3 857= factor established; V= measured volume of the specimen, and P= sample weight in grams.
The vitamin C content in the fruits was determined with the methodology reported by Padayatt et al. (2001). A 10 g fresh weight of fruit were weighed and placed in a mortar, triturated with 10 ml of hydrochloric acid 2% (v/v). The mixture in 40 ml of distilled water was homogenised. It was filtered through cheesecloth and collected in an Erlenmeyer flask. The 10 ml of the supernatant were taken and titrated with 2,6-dichlorophenol (1 X 10-3 N), when the solution reached pink. The vitamin C content was determined using the following formula:
Vitamin C (mg 100 g PF)= (ml used 2.6 dichlorophenol x 0088 x total volume x 100) / (volume of the aliquot x weight of sample).
Results and discussion
Growth of stem
The plant height and stem diameter of habanero hot pepper were significantly reduced (p≤ 0.5) from the ninth and fourth week respectively with increasing doses of P-Ca when compared with the (Figures 1 and 2) control. This behavior was maintained in both tissues during the growth cycle. The maximum values observed reduction in growth were 52% in stem height and 30.5% in the diameter of that body when compared to the control. The prohexadione calcium is a growth retardant which inhibits the synthesis of biologically active gibberellins A1, A4, and A7 (Brown et al., 1997; Evans et al., 1999). This effect is directly related to the reduction in vegetative growth observed in Figures 1 and 2, which reflects a decrease in cell elongation (Evans et al., 1999; Rademacher y Kober et al., 2003).
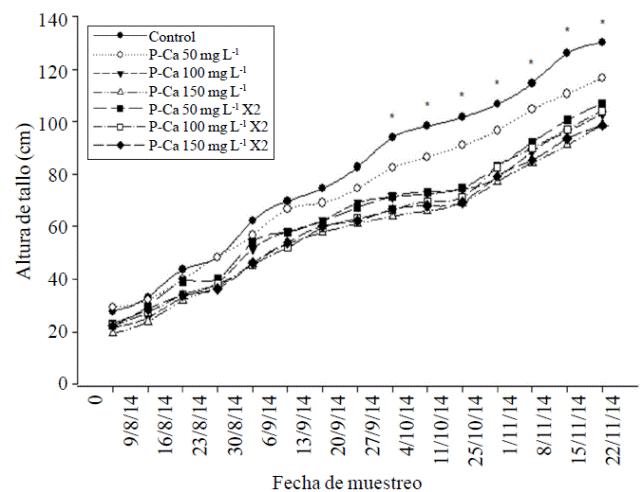
Figure 1 Effect of prohexadione calcium on the growth rate in stem height in habanero hot pepper var. Jaguar. Each point represents the average of nine plants. *indicates statistically significant difference according to the LSD test (p≤ 0.05).
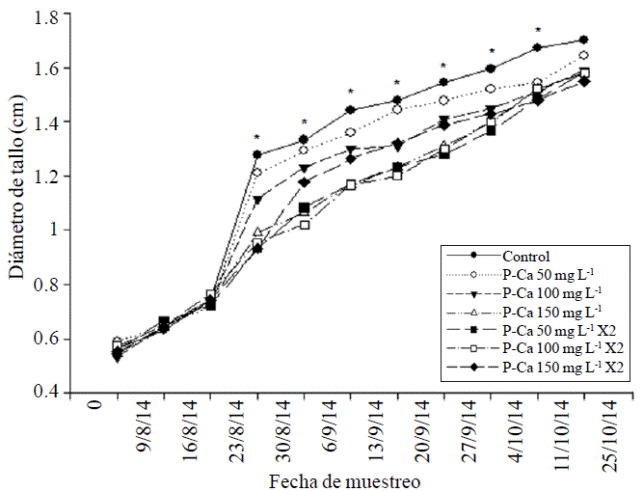
Figure 2 Effect of prohexadione calcium on the growth rate of stem diameter in habanero hot pepper var. Jaguar. Each point represents the average of nine plants. *indicates statistically significant difference according to the LSD test (p≤ 0.05).
Ramirez et al. (2008) identified in apexes of saladette tomato and chile pepper treated with P-Ca biologically inactive gibberellin GA20 and GA53, and demonstrated blocking the synthesis of GA1, GA4 and GA7 which are responsible for the growth of stem and diameter in both species. This effect of decreased growth has been observed in mirador chilli (Ramirez et al., 2010a), tomato peel (Ramirez et al., 2010c), jalapeno pepper (Ramirez et al., 2015) and fruits such as apple (Unrath 1999; Miller 2002; Basak 2007); and ornamental as petunia (Ilias and Rajapakse et al., 2005). The effect of P-Ca in stem diameter also previously observed in jalapeno (Ramirez et al., 2015).
At the end of the growth cycle, it was observed in most treatments P-Ca a tendency to restore growth, although not to the development of the control plants (Figures 1 and 2). Restoring growth of the main stem and diameter at the last stage of development has been observed in other horticultural species treated with P-Ca, and is related to the return of the synthesis of biologically active gibberellins at the apex of these plants (Rademacher, 2004).
Bloom, fruits and performance
Atrend to greater number off lowers per plant was observed as the concentration of P-Ca (Figure 3) increased. Bioregulator dose to 50 mg L-1 X 2 showed the highest flowering and significantly outperformed the control by 50%. Ramirez et al. (2009) state that is likely to be a synergism between P-Ca and increased bud cytokinins be stimulated floral, condition resulting in an increased number of flowers per plant. Ramirez et al. (2005) performed applying different concentrations of P-Ca in tomato hybrids and found that the growth retardant induced a significant increase in the content of cytokinins. This effect has also been reported in mirador chilli where there was an increase in the number of f lowers (Ramirez et al., 2010a); and fruits such as cherry (Elfving et al., 2003), apple, pear and plum (Basak and Rademacher et al., 1998).
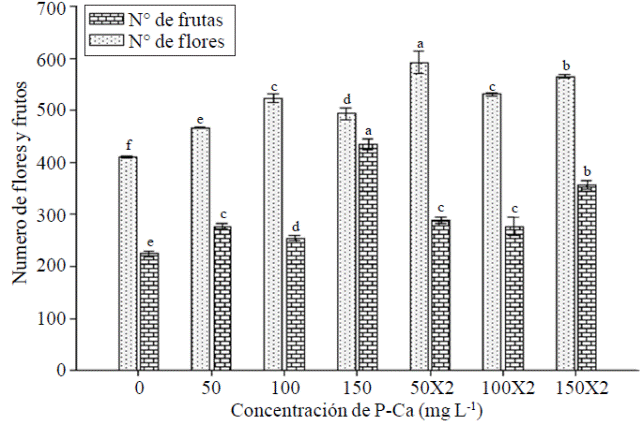
Figure 3 Effect of prohexadione calcium on number of f lowers and fruits habanero hot pepper var. Jaguar. Bars with the same letter are the same (LSD p≤ 0.05).
By adding the number of fruits per plant the four cuts made, it was observed that any treatment with P-Ca was significantly higher control (Figure 3). Treatment with P-Ca 150 mg L-1 caused the most outstanding to practically double the total number of fruits produced in the control increase. When the same dose of P-Ca was applied twice, an increase of 60% was achieved against control. The yield per plant also reflected the above effect. Production increased significantly in all treatments loa prohexadione calcium (Table 1); treatments protruding P-Ca 150 mg L-1 in one or two applications where increases of 43% and 36% respectively were observed when compared with the control again. The fruit size was reduced with most doses with P-Ca (Table 1).
Table 1 Effect of P-Ca on fruit length and yield habanero hot pepper var. Jaguar.
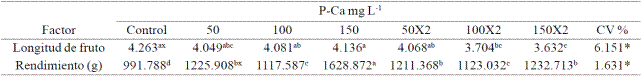
*,significativo a una p≤ (0.05); C.V. : coeficeinte de variación, x Valores con la misma letra en cada factor son iguales (LSD p≤ 0.05), cada factor representa el promedio de 9 plantas. X2= P-Ca aplicado dos veces.
Deka and Shadeque et al. (1996) mention that the performance depends on the accumulation of fotoasimilados and partitions in different parts of the plant. The P-Ca is a growth retardant capable of modifying the translocation of assimilates and redistribution of dry matter thereby causing an improvement in performance (Chetti, 1991); also stimulates flower induction and fruit set (Ramirez et al., 2005; Ramirez et al., 2010c). The increased number of flowers and observed in Figure 1 and the increase in reported in Table 1 yield fruits, the above support. Inhibition of vegetative growth possibly made available largest reserves of food for fruit set and fruit development (Ma and Smith, 1992). Have been reported jobs with similar results in jalapeno with P-Ca wherein an increase in performance was observed (Ramírez et al., 2009; Ramirez et al., 2010a; Ramirez et al., 2015) and tomato peel and saladette (Ramirez et al., 2005; Ramirez et al., 2010c). The reduction in fruit length caused by P-Ca may be linked to the significant increase in fruit set observed between treatments with growth retardant; however, this phenotype can be offset by the qualitative enrichment harvested fruit as has already been demonstrated in cherry and other fruit (Rademacher and Kober, 2003).
Antioxidants
Capsaicin
The Prohexadione increased Ca content in fruits capsaicin any dose tested (Figure 4). Treatment with P-Ca 150 mg L-1 caused the highest content of capsaicin. The dose 50 mg L-1 applied in one or two occasions also caused substantial increases in this antioxidant.
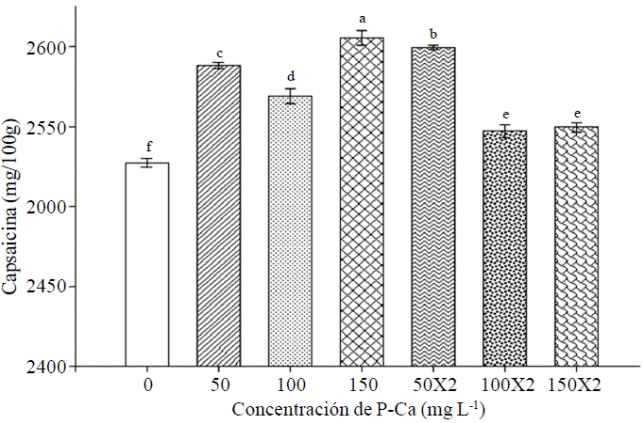
Figure 4 Effect of prohexadione calcium on levels of capsaicin in fruits habanero hot pepper var. Jaguar. Each bar represents the mean of three replicates. Bars with the same letter are the same (LSD p≤ 0.05).
Information on the effects of P-Ca in habanero hot pepper is very low; however, there are reports of increases in the level of capsaicin in jalapeno fruits treated with P-Ca (Ramirez et al., 2009; Ramirez et al., 2010a). May prohexadione calcium enzyme level modify the biosynthetic pathway of flavonoids, generating modified flavonoid linked to antioxidant activity in young tissues (Rademacher, 2004; Ramirez et al., 2010a). Capsaicin synthesis where its chemical structure a phenolic nucleus is linked via an amide linkage to a fatty acid is known; wherein the phenolic portion is vainillilamina, which is formed from phenylalanine via the route fenilporpanoides and fatty acid is formed from amino acids branched side chain, either valine or leucine (Vazquez et al., 2007). It suggested that P-Ca then could prolong this process resulting in increased production of capsaicna (Evans et al., 1999.)
Total carotenoid
Most P-Ca treatments modified the content of total carotenoids in fruits habanero hot pepper (Figure 5). This effect was greater when the P-Ca was applied to 50 mg L-1 in one or two applications causing increased total carotenoid 22% and 38% respectively when compared with control.
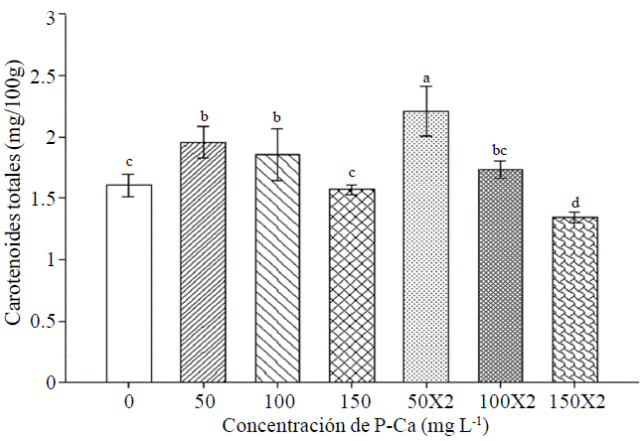
Figure 5 Effect of prohexadione calcium on the content of total carotenoids in fruits habanero hot pepper var. Jaguar. Each bar represents the mean of three replicates. Bars with the same letter are the same (LSD p≤ 0.05).
Information on the possible effects and prohexadione calcium metabolism and synthesis of carotenoids in fruits of habanero hot pepper is very limited. In jalapeno it has reported evidence of an increase in carotenes with the application of P-Ca. This effect is very strong with increased levels of luteolin in ripe fruit (Ramirez et al., 2015). Tomato Floradade P-Ca increased lycopene content in ripe fruit (Ramirez et al., 2010b). The growth retardant caused increases in carotenoid content, thereby improving the color of tangerine peel and orange (Barry and Roux, 2010). Therefore, it should further investigate the influence of P-Ca metabolism fruit ripening.
Vitamin C
The prohexadione calcium stimulated an increase in the content of vitamin C in fruits ripe habanero hot pepper var. Jaguar (Figure 6). The bioregulator treatments applied in concentrations of 100 and 150 mg L-1 in one or two asperciones caused on average 148% increase in vitamin C content when compared with control fruits. This effect was previously observed in jalapeno (Ramirez et al., 2009; Ramirez et al., 2010a) and tomato peel (Ramirez et al., 2010c). Rademacher and Kober (2003) report that ethylene is generated from aminocyclopropanecarboxylic acid (ACC) in a reaction catalyzed by ACC oxidase. This is a dioxygenase which requires ascorbic acid as a co-substrate and P-Ca is also inhibitory for this enzyme. Therefore, the hypothesis that P-Ca by inhibiting the synthesis of ethylene causes an accumulation of ascorbic acid as P-Ca compete for the active site in the ethylene synthesis.
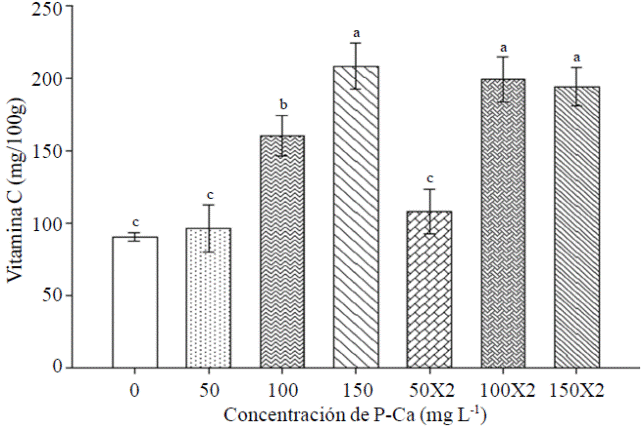
Conclusions
With the results obtained and the conditions under which it was conducted this investigation is concluded that P-Ca reduces the growth of stem height and diameter; increases flowering, fruit number and yield per plant; and it causes an increase in the capsaicin content, total carotenoids and vitamin C in ripe fruits habanero hot pepper of var. Jaguar.
Literatura citada
Antonious, G. F. and Jarret, R. L. 2006. Screening Capsicum accessions for capsaicinoids content. J. Environ. Sci. Health Part B. 41(5):717-729. [ Links ]
Antonious, G. F.; Kochhar, T. S. and Jarret R. L. 2009. Pungency in Capsicum chinense: variation among countries of origin. J. Environ. Sci. Health Part B. 44(2):179-184. [ Links ]
Barry, G. H. and Le Roux, S. 2010. Preharvest foliar sprays of prohexadione -calcium, a gibberellin-biosynthesis inhibitor, induce chlorophyll degradation and carotenoid synthesis in citrus rinds. HortSci. 45(2):242-247. [ Links ]
Basak, A. 2007. The effect of prohexadione-Ca on shoot growth and cropping of young apple trees of Jonagold cv. Roczniki Akademii Rolniczej w Poznaniu. Ogrodnictwo. 41: 261-268. [ Links ]
Basak, A. and Rademacher, W. 1998. Growth regulation of pome and stone fruit trees by use of prohexanedione-Ca. In: XXV International Horticultural Congress, Part 4: Culture Techniques with Special Emphasis on Environmental Implications. 514:41-50. [ Links ]
Bennett, D. J. and Kirby, G. W. 1968. Constitution and biosynthesis of capsaicin. J. Chem. Soc. C: Organic. 442-446 pp. [ Links ]
Brown, R. G.; Kawaide, H.; Yang, Y. Y.; Rademacher, W. and Kamiya, Y. 1997. Daminozide and prohexadione have similar modes of action as inhibitors of the late stages of gibberellin metabolism. Physiol. Plantarum. 101(2):309-313. [ Links ]
Chetti, M. B. 1991. Evaluation of Chamatkar on groundnut. Pestology. 15:43-50. [ Links ]
Deka, P. C. and Shadeque, A. 1996. Effect of foliar sprays of cycocel (CCC) on the growth and yield of bell pepper. Hort. J. 9(2):144-147. [ Links ]
Elfving, D. C.; Lang, G. A. and Visser, D. B. 2003. Prohexadione-Ca and ethephon reduce shoot growth and increase flowering in young, vigorous sweet cherry trees. HortSci. 38(2):293-298. [ Links ]
Evans, J. R.; Ishida, C. A.; Regusci, C. L. and Rademacher, W. 1999. Mode of action, metabolism and uptake of BAS-125W, Prohexadione- calcium. HortSci. 34(7):1200-1201. [ Links ]
Howard, L. R.; Talcott, S. T.; Brenes, C. H. and Villalon, B. 2000. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem. 48(5):1713-1720. [ Links ]
Ilias, I. F. and Rajapakse, N. 2005. Prohexadione-calcium affects growth and flowering of petunia and impatiens grown under photoselective films. Sci. Hortic. 106(2):90-202. [ Links ]
Krajewska, A. M. and Powers, J. J. 1988. Sensory properties of naturally occurring capsaicinoids. J. Food Sci. 53(3):902-905. [ Links ]
Ma, B. L. and Smith, D. L. 1992. Growth regulator effects on aboveground dry matter partitioning during grain fill of spring barley. Crop Sci. 32(3):741-746. [ Links ]
Miller, S. S. 2002. Prohexadione-calcium controls vegetative shoot growth in apple. Journal of tree fruit production. 3(1):11-28. [ Links ]
Padayatt, S. J.; Daruwala, R.; Wang, Y.; Eck, P. K.; Song, J.; Koh, W. S. and Levine, M. 2001. Vitamin C: from molecular actions to optimum intake. In: handbook of antioxidants. Cadenzas, E.; Packer, l. (Eds). Second edition. CRC press. Washington DC, USA. 117-145 pp. [ Links ]
Rademacher, W. 2004. Chemical regulation of shoot growth in fruit trees. Acta Hortic. 653: 9-15. [ Links ]
Rademacher, W. and Kober, R. 2003. Efficient use of prohexadione-Ca in pome fruits. Eur. J. Horticultural Sci. 68(3):101-107. [ Links ]
Ramírez, H. 2003. El uso de hormonas en la producción de cultivos hortícolas para exportación. In: Memoria del Tercer Simposio Nacional de Horticultura. Saltillo, Coahuila, México. 1-22 pp. [ Links ]
Ramírez, H.; Amado-Ramírez, C.; Benavides-Mendoza, A.; Robledo- Torres, V. y Martínez-Osorio, A. 2010a. Prohexadiona-Ca, AG3, ANOXA y BA modif ican indicadores fisiológicos y bioquímicos en chile Mirador. Rev. Chapingo Ser. Hortic. 16(2):83-89. [ Links ]
Ramírez, H.; Camacho-Chávez, V. M.; Ramírez-Pérez, L. J.; Rancaño- Arrioja, J. H.; Sepúlveda-Torre, L. y Robledo-Torres, V. 2015. La prohexadiona-Ca provoca cambios en el crecimiento vegetativo, giberelinas, rendimiento y luteolina en chile jalapeño. Ecosistemas y Recursos Agropecuarios. 2(4):13-22. [ Links ]
Ramírez, H.; Herrera-Gámez, B.; Benavides-Mendoza, A.; Rancaño- Arrioja, J. H.; Álvarez-Mares, V.; Amado-Ramírez, C. y Martínez-Osorio, A. 2010b. Prohexadiona de Calcio incrementa la capacidad antioxidante, el contenido de licopeno y la actividad enzimática en frutos de tomate Floradade. Rev. Chapingo Ser. Hortic. 16(3):155-160. [ Links ]
Ramírez, H.; Herrera-Gámez, B.; Méndez-Quiroa, Y. H.; Benavides- Mendoza, A.; Álvarez-Mares, V.; Rancaño-Arrioja, J. H. y Villareal-Quintanilla, J. A. 2008. Prohexadiona de calcio disminuye el contenido de giberelinas endógenas en ápices de tomate saladette y chile pimiento. Rev. Chapingo Ser. Hortic. 14(2):193-198. [ Links ]
Ramírez, H.; Méndez-Paredes, O.; Benavides-Mendoza, A. y Amado-Ramírez, C. 2009. Influencia de prohexadiona-Ca y promotores de oxidación sobre el rendimiento, capsaicina y vitamina C en chile jalapeño. Rev. Chapingo Ser. Hortic. 15(3):231-236. [ Links ]
Ramírez, H.; Peralta-Manjarrez, R. M.; Benavides-Mendoza, A.; Sánchez-López, A.; Robledo-Torres, V. y Hernández-Dávila, J. 2005. Efecto de prohexadiona-Ca en tomate y su relación con la variación de la concentración de giberelinas y citocininas. Rev. Chapingo Ser. Hortic. 11(2):283-290. [ Links ]
Ramírez, H.; Rivera-Cruz, C. E.; Benavides-Mendoza, A.; Robledo- Torres, V. y Reyna-Sustaita, G. 2010c. Prohexadiona-Ca, una alternativa en la producción de tomate de cáscara (Physalis ixocarpa Brot.). Rev. Chapingo Ser. Hortic. 16(2):139-146. [ Links ]
Rodríguez-Maturino, A.; Valenzuela-Solorio, A.; Troncoso-Rojas, R.; González-Mendoza, D.; Grimaldo-Juarez, O.; Aviles-Marin, M. and Cervantes-Diaz, L. 2012. Antioxidant activity and bioactive compounds of Chiltepin (Capsicum annuum var. glabriusculum) and Habanero (Capsicum chinense): a comparative study. J. Med. Plants Res. 6(9):1758-1763. [ Links ]
Tomas, P. 1975. Effect of post-harvest temperature on quality carotenoids and ascorbic content of Alphonso mangoes on ripening. J. Food Sci. 40(4):704-706. [ Links ]
Unrath, C. R. 1999. Prohexadione-Ca: a promising chemical for controlling vegetative growth of apples. HortSci. 34(7):1197- 1200. [ Links ]
Vázquez-Flota, F.; Miranda-Ham, M. L.; Monforte-González, M.; Gutiérrez-Carbajal, G.; Velázquez-García, C. y Nieto-Pelayo, Y. 2007. La biosíntesis de capsaicinoides, el principio picante del chile. Rev. Fitotec. Mex. 30(4): 353-360. [ Links ]
Wall, M. M.; Waddell, C. A. and Bosland, P. W. 2001. Variation in β-carotene and total carotenoid content in fruits of Capsicum. HortSci. 36(4):746-749. [ Links ]
Received: February 2016; Accepted: May 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
