











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las variedades de importancia comercial para producción de nopal verdura en México son Milpa Alta, Atlixco, COPENAS y Valtierrilla (Flores et al., 1995). La norma internacional de calidad de nopal, CODEX STAN 185-1993, establece que los cladodios tiernos pueden comercializarse cuando estos presentan desde 9 cm hasta 30 cm de longitud (FAO- WHO, 1993). Los cladodios de 20 cm de longitud y peso de 90-100 g, se comercializan como penca entera (Cantwell, 1999); mientras que los cladodios más pequeños (12 cm) o más largos (casi 30 cm), se utilizan para la elaboración de productos frescos-cortados (Rodríguez, 2002).
El oscurecimiento enzimático es uno de los principales problemas que limita la comercialización de nopal verdura, tanto en su forma entera como fresca-cortada (Rodríguez, 2002). Este deterioro tiene gran importancia por su impacto visual que afecta la aceptación organoléptica y la calidad comercial, y por reducir el valor nutricio de frutas y hortalizas (Artés et al., 2007). El oscurecimiento enzimático se presenta en la superficie de cortes y heridas y es causado por la enzima polifenoloxidasa (PPO). Esta enzima en presencia de oxígeno actúa sobre los sustratos fenólicos, formando las quinonas. Estos compuestos reaccionan entre sí o con otros compuestos (aminoácidos, péptidos y azúcares reductores) formando melaninas, compuestos de color rojo, café o negro (Artés et al., 2007). Sin embargo, la formación de quinonas es reversible en presencia de agentes reductores como el ácido ascórbico, los cuales son capaces de reducir las quinonas a los compuestos fenólicos nativos; mientras que la formación de melaninas es irreversible (Barbagallo et al., 2009). El oscurecimiento enzimático ocurre cuando los sustratos fenólicos, la PPO y el oxígeno molecular se unen bajo condiciones apropiadas de pH, temperatura y actividad de agua (Artés et al., 1998).
Existen diferencias en el grado de oscurecimiento, en el contenido y tipo de compuestos fenólicos, en la actividad de PPO y en el contenido de ácido ascórbico entre variedades de hortalizas como papa, alcachofa y otras (Barbagallo et al., 2009; Cabezas et al., 2009a b). Adicionalmente, se ha observado que existen cambios en la actividad de PPO, contenido de compuestos fenólicos, oscurecimiento y contenido de ácido ascórbico durante el desarrollo de hortalizas de hoja como col rizada (Brassica oleracea var. Acephala) (Korus, 2011) y lechuga (Chutichudet et al., 2011). Aguilar et al. (2007) evaluaron la actividad de PPO y el potencial de oscurecimiento en cladodios de 21 variedades de Opuntia, encontrando diferencias significativas entre las variedades, presentando ‘Chicomostoc’, ‘Negrito’y ‘Jade’, los menores valores de oscurecimiento.
Sin embargo, los valores de actividad de PPO de estas tres variedades fue similar al de otras variedades que presentaron un mayor potencial de oscurecimiento. Estos autores no evaluaron otras variables como el contenido de ácido ascórbico (reductor de quinonas), fenoles totales, ni el contenido de sustratos de PPO, que pueden influir en el desarrollo del oscurecimiento. Se ha observado que el oscurecimiento del tejido ocurre cuando el contenido de ácido ascórbico se ha agotado (Adams y Brown, 2007). Adicionalmente, se ha indicado que el ácido ascórbico puede inhibir a PPO al disminuir el pH del citosol (Yoruk y Marshall, 2003). Degl’Innocenti et al. (2007) concluyeron que el alto contenido de ácido ascórbico de rúcula (Eruca sativa) está altamente asociado con la resistencia al oscurecimiento mostrada durante el almacenamiento de esta hortaliza en su forma fresca-cortada, mientras que Bottino et al. (2009) observaron un comportamiento similar en espinaca.
Aunque se ha evaluado la actividad de PPO y el oscurecimiento de diferentes variedades de nopal, se desconoce si hay cambios en el contenido de compuestos fenólicos y ácido ascórbico durante el desarrollo de los cladodios de los tamaños comerciales establecidos por la norma de la Comisión del Codex Alimentarius (CODEX STAN 185-1993). Por lo anterior, el objetivo de este trabajo fue estudiar la actividad de polifenoloxidasa (PPO), el contenido de compuestos fenólicos y de ácido ascórbico, y el grado de oscurecimiento durante el desarrollo de nopal verdura de variedades comerciales.
Materiales y métodos
Material experimental
Se utilizó nopal verdura (Opuntia ficus indica) de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco, de una plantación comercial ubicada en Hermosillo, Sonora, México (29° 11' 27.53" latitud norte, 110° 51' 17.13" longitud oeste). Las cuatro variedades de nopal estuvieron bajo las mismas condiciones de cultivo comercial, mismo tipo de suelo, clima y manejo agronómico. La cosecha de los cladodios de las cuatro variedades y tamaños se realizó manualmente el mismo día y a la misma hora, en el mes de julio. Se cosecharon cladodios tiernos, con presencia de hojas, y se transportaron inmediatamente hacia los laboratorios de la Institución, donde se seleccionaron cladodios firmes y libres de defectos. Se clasificaron en seis estados de desarrollo con base a longitud, incluyendo cladodios de 9 a 30 cm de longitud, tamaños considerados comerciales por la norma internacional de calidad de nopal, CODEX STAN 185-1993 (FAO-WHO, 1993). A continuación se les midió longitud, peso y grosor (Cuadro 1), y se almacenaron a -20°C.
Cuadro 1 Longitud, peso y grosor durante el desarrollo de los cladodios de nopal verdura de las variedades COPENA F-1. COPENA V-1, Milpa Alta y Atlixco.

*Los datos representan el promedio y su desviación estándar, n= 30.
Posteriormente fueron troceados y almacenados de nuevo a -20°C. En el tejido congelado se realizaron los análisis de grado de oscurecimiento y ácido ascórbico; y se prepararon polvos de acetona para análisis de actividad de PPO. Adicionalmente, se tomaron por triplicado, 150 g de nopal congelado, se liofilizaron en un liofilizador de charolas Freezemobile UNITOP 600L. El nopal liofilizado se colocó en frascos de plástico herméticamente sellados, los cuales se almacenaron a -20°C, estas muestras liofilizadas se utilizaron en la determinación de fenoles totales. Todas las determinaciones se realizaron por triplicado.
Metodología
Extracción y actividad específica de PPO. La preparación del polvo de acetona y la extracción de PPO se realizaron de acuerdo al procedimiento indicado por Sciancalepore y Longone (1984). Se pesaron 200 g de nopal finamente picado, se les agregaron 400 mL de acetona fría (-20°C) y 1.4 g de polietilénglicol, se homogeneizaron en una licuadora. El homogeneizado obtenido se filtró con vacío utilizando papel Whatman No.1. Al residuo se le realizaron de 2 a 3 lavados con 200 mL de acetona fría, hasta que se obtuvo un polvo de color blanco (polvo de acetona), éste se dejó secar a temperatura ambiente por 24 h, se colocó en frascos de plástico sellados y se almacenaron a -20°C. Para la extracción de PPO se pesaron 0.3 g de polvo de acetona y se les agregó 40 mL de fosfato de potasio (K3PO4) 0.05 M que contenía KCl 1 M a un pH de 8.5. Esta mezcla se sometió a agitación constante por 1 hora a 5 °C. Posteriormente, la muestra se centrifugó a 4101 x g a 4°C por un tiempo de 45 min. El sobrenadante se filtró con una tela de organza. El filtrado obtenido representó el extracto enzimático, que se dividió en dos partes que se utilizaron para análisis de proteína y medición de actividad específica de PPO.
La actividad específica de PPO se determinó midiendo la velocidad inicial de formación de quinona siguiendo la metodología propuesta por Sciancalepore y Longone (1984). A un mL del extracto enzimático se le agregó 1 mL de 4-metilcatecol 0.02 M en buffer de citratos 0.1 M a pH 3.5 (definido en pruebas preliminares como el pH óptimo para la actividad de PPO de nopal). Se midió la velocidad inicial de formación de quinona indicada por el incremento en absorbancia a 395 nm. Se calculó la pendiente de la parte inicial de la curva, expresando los resultados como unidades de actividad por miligramo de proteína. Una unidad de actividad de la enzima se define como la cantidad de enzima que causa un cambio en absorbancia de 0.01/ min.
El contenido de proteína soluble en los extractos enzimáticos se determinó por el método de Bradford, utilizando albúmina de suero de bovino como estándar (Bradford, 1976).
Fenoles totales. En 500 mg de nopal liofilizado se efectuaron tres extracciones sucesivas sonicando con metanol al 80% de acuerdo a lo propuesto por Escarpa y González (2000) y Kähkönen et al. (1999). La determinación de los fenoles totales se efectuó de acuerdo al procedimiento de Singleton y Rossi, (1965). El incremento en absorbancia a 765 nm se relaciona con la concentración de fenoles totales con una curva estándar preparada con ácido gálico. Los resultados se expresaron como mg de equivalentes de ácido gálico (EAG)/ g de peso seco.
Ácido ascórbico total. La extracción se realizó en 30 g de muestra congelada que se homogeneizó con 100 mL de solución extractora (30 g de ácido fosfórico, 80 mL de ácido acético en un litro de agua), se filtró en tela de organza y posteriormente en filtros Millipore de 0.22 μm. La determinación se realizó de acuerdo a la técnica de Doner y Hicks, (1981) en un HPLC Varian 9012, con una columna μBondapak NH2, a una longitud de onda de 268 nm. Se utilizó un estándar externo de ácido L-ascórbico a 0.05 mg/ mL. Se empleó ditiotreitol (DTT) para reducir el ácido dehidroascórbico a ácido L-ascórbico. Los resultados se expresaron como mg de ácido ascórbico total/ 100 g de peso fresco.
Grado de oscurecimiento. Este método se basa en la medición de la absorbancia de las quinonas formadas debido a la oxidación de los compuestos fenólicos por PPO en presencia de oxígeno. La longitud de onda de máxima absorbancia de la quinona formada a partir de pirocatecol y L-DOPA es de 390 y 480 nm, respectivamente (Nicolas et al., 1994). 20 g de muestra se homogeneizaron con 100 ml de buffer de citratos pH 4.46 (pH promedio del nopal verdura), se incubaron en un baño de agua a 35°C y agitación constante por 48 h. Posteriormente, las muestras se filtraron con tela de organza y el filtrado se centrifugó por 10 min a 18 500 rpm en una centrífuga refrigerada Beckman Model J2-21. En pruebas preliminares se midió la absorbancia del extracto de 380 a 510 nm y se encontró que la mayor absorbancia se obtuvo a 420 nm. Se leyó la absorbancia del sobrenadante a 420 nm en un espectrofotómetro Perkin Elmer Lambda 3A (UV-VIS). Los resultados se expresaron como valores de absorbancia a 420 nm (Sapis et al., 1983).
Diseño experimental y análisis estadístico. Se empleó un diseño experimental en bloques con los factores variedad y estado de desarrollo, en el que el factor variedad fue utilizado como bloque. Se realizó un análisis de varianza por modelos lineales generales (GLM) a un nivel de significancia de 0.05. Cuando hubo significancia se efectuaron comparaciones de medias por la prueba de Tukey. Todos los datos fueron procesados en el paquete estadístico computacional NCSS (Hintze, 1997).
Resultados y discusión
Las características morfológicas de las cuatro variedades de nopal estudiadas son diferentes. Los cladodios de nopal COPENA F-1 se caracterizan por ser largos y delgados con muchas espinas; mientras que los de COPENA V-1 son discoides y con pocas espinas. Los cladodios de COPENA F-1 presentaron la mayor longitud y los de COPENA V-1 la menor, los de Milpa Alta y Atlixco presentaron longitudes similares (Cuadro 1). La variedad Atlixco presentó el mayor peso y la variedad COPENA V-1 el menor; mientras que COPENA F-1 y Milpa Alta presentaron un peso similar. El grosor de los cladodios de COPENA V-1 fue mayor y el de los de Milpa Alta fue el menor.
Actividad específica de PPO. En las cuatro variedades de nopal verdura estudiadas, la actividad específica de PPO mostró una tendencia a aumentar significativamente (p< 0.05) a medida que avanzó el estado de desarrollo del cladodio, con valores de 36.83 unidades de actividad por miligramo de proteína en el estado I hasta alcanzar valores de 62.08 unidades de actividad por miligramo de proteína en el estado VI; lo cual representa un incremento de 168% (Figura 1). Comparando la actividad específica de PPO de las diferentes variedades no se encontró efecto (p> 0.05) de la variedad en la actividad de esta enzima, mostrando estas valores promedio similares de 48.71, 46.81, 49.15 y 47.43 unidades de actividad por miligramo de proteína, para COPENA F-1, COPENA V-1, Milpa Alta y Atlixco, respectivamente.

Figura 1 Actividad específica de polifenoloxidasa durante el desarrollo de cladodios de nopal verdura de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco. Una unidad de actividad (U) se definió como la cantidad de enzima que causa un incremento en absorbancia de 0.01/min. Las barras verticales indican la desviación estándar para cada estado de desarrollo, n= 3.
Estos resultados concuerdan con lo publicado por Aguilar et al. (2007) quienes encontraron diferencias no significativas en la actividad de PPO de los cladodios de aproximadamente 20 cm de las variedades COPENA F-1, Atlixco y Milpa Alta. Se ha mostrado que el pH causa cambios conformacionales en la enzima y afecta la cinética de actividad de PPO. El pH óptimo de PPO varía dependiendo del vegetal, con valores de pH óptimos entre 4 y 8. Se ha estimado que el pH óptimo de PPO de manzana y uva es de 3.5 a 4.0 (Yoruk y Marshall, 2003). En este estudio se determinó que 3.5 es el pH óptimo para la actividad de PPO de nopal, este valor es similar al pH óptimo de actividad de PPO de manzana y uva. En el presente estudio los resultados de actividad específica de PPO se expresaron como unidades de actividad por mg proteína, mientras que Aguilar et al. (2007) expresaron sus resultados como unidades de actividad por g de peso fresco, por lo anterior no es posible comparar los valores de actividad del presente estudio con los de estos autores. El incremento en la actividad de PPO observado durante el desarrollo de nopal verdura concuerda con lo observado por Korus (2011), quien mostró que la actividad de PPO se incrementó a medida que avanzó el estado de madurez de la hoja de col rizada.
Fenoles totales. El estado de desarrollo tuvo un efecto significativo en el contenido de fenoles totales de los cladodios de nopal de las cuatro variedades estudiadas (p< 0.05), encontrando que los cladodios en estado de desarrollo II tuvieron un mayor contenido que los cladodios de los estados V y VI (Figura 2). Asimismo, se encontraron diferencias significativas (p< 0.05) en el contenido de estos compuestos en las distintas variedades, observándose que COPENA F-1 presentó el mayor contenido (19.89 mg EAG/ g peso seco), seguido de Milpa Alta (16.06 mg EAG/ g peso seco) que presentó valores similares a COPENA V-1 (13.85 mg EAG/ g peso seco) y por último Atlixco presentó el menor contenido (12.12mgEAG/g peso seco), aunque similar a COPENAV-1.

Figura 2 Contenido de fenoles totales durante el desarrollo de cladodios de nopal verdura de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco. EAG: equivalentes de ácido gálico. Las barras verticales indican la desviación estándar para cada estado de desarrollo, n= 3.
Al igual que en el presente estudio, Guevara et al. (2010) encontraron diferencias significativas en el contenido de fenoles totales de 10 variedades de nopal (8 silvestres y 2 cultivadas) colectadas en diferentes regiones de México, observándose los valores más altos (19.9 mg EAG/ g peso seco) en la variedad Morado y los menores en los cladodios de la variedad Tapón (menores a 2 mg EAG/g peso seco). Estos autores atribuyen las diferencias encontradas entre las variedades a diferencias en las condiciones climáticas. Sin embargo, en el presente estudio las cuatro variedades evaluadas fueron cultivadas en las mismas condiciones y fueron muestreadas el mismo día, lo cual sugiere que las diferencias son intrínsecas a la variedad. El contenido de fenoles totales de los cladodios de COPENA F-1 es similar al encontrado por Guevara et al. (2010) en la variedad silvestre Morado (19.9 mg EAG/ g peso seco). El contenido de fenoles totales de las cuatro variedades analizadas en este estudio es mayor al de la variedad comercial de O. ficus indica, Blanco que tuvo un contenido de 5.25 mg EAG/ g peso seco (Guevara et al., 2010). Sin embargo, los valores encontrados en los cladodios de Milpa Alta del presente estudio son inferiores al de 32.20 mg EAG/ g peso seco mostrado por Corral et al. (2008), lo cual pudiera atribuirse a diferencias en las condiciones climáticas de las regiones de cultivo.
El contenido de fenoles totales de nopal verdura varió de 10.63-24.23 mg de equivalentes de ácido gálico/ g de peso seco en los distintos estados de desarrollo de las variedades estudiadas. Estos valores son mayores a los de otros productos hortícolas como cebolla, pepino, zanahoria, tomate y papa, que mostraron valores de 0.4-6.6 mg de EAG/ g de peso seco (Kähkönen et al., 1999).
Ácido ascórbico total. El contenido de AA total en las cuatro variedades estudiadas disminuyó significativamente (p< 0.05) conforme avanzó el desarrollo del cladodio, con valores de 13.50 a 7.41 mg de AA/100 g de peso fresco en los cladodios de los estados I y VI, respectivamente (Figura 3). Lo anterior indica una disminución de casi el 50% del contenido. Se encontraron diferencias significativas (p< 0.05) en el contenido de AA entre las variedades estudiadas, se observó que COPENAF-1 presentó un contenido mayor (11.90 mg de AA/ 100 g de peso fresco), seguido de COPENA V-1 (10.72 mg de AA/ 100 g de peso fresco); mientras que Milpa Alta y Atlixco presentaron el menor contenido (8.79 y 8.19 mg de AA/ 100 g de peso fresco, respectivamente). Estos resultados muestran que los cladodios de COPENA F-1 presentaron un contenido de AA40% mayor que los de Milpa Alta y Atlixco.
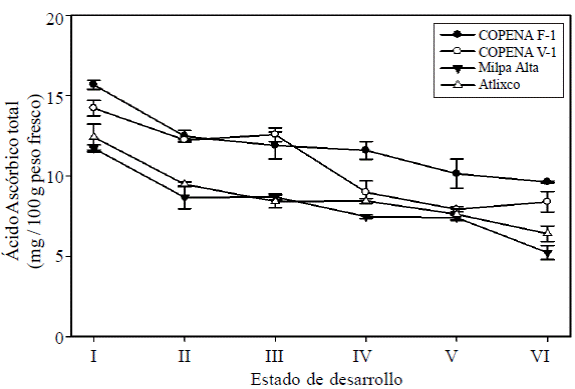
Figura 3 Contenido de ácido ascórbico total durante el desarrollo de cladodios de nopal verdura de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco. Las barras verticales indican la desviación estándar para cada estado de desarrollo, n= 3.
Rodriguez y Cantwell (1988), no encontraron un patrón consistente de cambio en el contenido de ácido ascórbico durante el desarrollo de cladodios de tres especies de Opuntia, con valores de 7 a 18 mg de AA/ 100 g de peso fresco, aunque cabe resaltar que estos autores emplearon un método colorimétrico a diferencia del presente estudio que utilizó la técnica de HPLC. Estos resultados concuerdan con los de Rodriguez y Villegas (1997) quienes reportaron valores de 12 mg de AA/ 100 g de peso fresco en cladodios de 20 cm de longitud de dos variedades de nopal verdura (COPENA F-1 y COPENAV-1). Corral et al. (2008) mostraron un contenido de 22.2 mg de AA/ 100 g de peso fresco en cladodios de Milpa Alta cultivados en el Estado de México, México; valores superiores a los del presente estudio. Estas diferencias pudieran atribuirse a diferencias en la metodología empleada en la determinación o al efecto de las condiciones climáticas de los sitios de cultivo, ya que se ha mostrado que tanto la intensidad de luz como la temperatura ambiental tienen influencia en el contenido de AA (Lee y Kader, 2000).
El mayor contenido de AA en los primeros estados de desarrollo de los cladodios de nopal concuerda con lo mostrado por Parsons y Fry (2012), quienes indicaron que las altas concentraciones de ácido ascórbico son características de tejidos en rápido crecimiento, a pesar de que estos también presentan altas actividades de ascorbato oxidasa, enzima que inicia el catabolismo del ácido ascórbico.
El contenido de AA total de nopal verdura obtenido en este estudio es similar al reportado para plátano, papas frescas y tomates frescos; aunque es menor al reportado para zarzamora, melón, uvas, kiwi, mandarina, naranja, frambuesa, brócoli, berza, coliflor y espinacas (Lee y Kader, 2000). Adicionalmente se observó que en nopal, el ácido L-ascórbico es el principal componente del ácido ascórbico total, con aproximadamente 80% en promedio para los diferentes estados de desarrollo y variedades; este comportamiento es similar al mostrado por Lee y Kader (2000) en diferentes frutas y hortalizas.
Se encontró una relación inversa entre ácido ascórbico y actividad específica de PPO con coeficientes de correlación de -0.80, -0.93, -0.88 y -0.87 en las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco, respectivamente. Este comportamiento muestra que al disminuir el contenido de ácido ascórbico total aumenta la actividad específica de PPO.
Oscurecimiento. Los valores de oscurecimiento mostrados por los cladodios de nopal verdura de las distintas variedades y estados de desarrollo fueron bajos, aún después de 48 h con valores de absorbancia a 420 nm de 0 a 0.2 (Figura 4), encontrando que no hay cambios significativos (p> 0.05) en el grado de oscurecimiento a 420 nm durante el desarrollo de los cladodios de las cuatro variedades evaluadas. Sin embargo, se encontraron diferencias significativas (p< 0.05) en el oscurecimiento entre las distintas variedades, observándose que COPENA F-1 y Milpa Alta presentaron los valores más altos, seguido de las variedades COPENA V-1 y Atlixco que presentaron valores similares.

Figura 4 Grado de oscurecimiento (Abs 420 nm) durante el desarrollo de cladodios de nopal verdura de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco. Las barras verticales indican la desviación estándar para cada estado de desarrollo, n= 3.
Los resultados del presente estudio concuerdan con los mostrados por Aguilar et al. (2007), quienes evaluaron la actividad de PPO y el potencial de oscurecimiento en cladodios de 30 días de edad de 21 variedades de Opuntia, encontrando diferencias significativas entre las variedades analizadas, presentando ‘Chicomostoc’, ‘Negrito’y ‘Jade’, los menores valores de oscurecimiento. Sin embargo, los valores de actividad de PPO de estas variedades fueron similares al de otras variedades que presentaron un mayor potencial de oscurecimiento. Artés et al. (1998) indicaron que la especie y la variedad se encuentran entre los factores más importantes que influyen en la susceptibilidad al oscurecimiento de las frutas y hortalizas, resaltando que en papa y manzana la susceptibilidad al oscurecimiento depende de la variedad.
Cabezas et al. (2009a) evaluaron cinco variedades de papa (Agata, Agria, Almera, Marabel y Vivaldi), encontrando diferencias en el oscurecimiento entre variedades, mostrando que Marabel se caracterizó por presentar un bajo contenido de fenoles totales, una baja actividad de PPO y menor incidencia de oscurecimiento, mientras que Almera presentó el mayor contenido de fenoles totales, de actividad de PPO y de incidencia de oscurecimiento. De igual manera, Cabezas et al. (2009b) encontraron diferencias en la susceptibilidad al oscurecimiento de cinco variedades de alcachofa (C3, Catanese, Tema, Violetto Foggiano y Violetto Sardo), indicando que C3 mostró el mayor contenido de fenoles totales y de actividad de PPO, así como la mayor incidencia de oscurecimiento; mientras que Catanese mostró el mayor contenido de ácido ascórbico total y la más baja incidencia de oscurecimiento.
Los resultados del presente estudio muestran que tal y como se indicó en papa y manzana (Artés et al., 1998), papa (Cabezas et al., 2009a) y alcachofa (Cabezas et al., 2009b) la variedad es el factor que más influye en la susceptibilidad al oscurecimiento de los cladodios de nopal verdura, ya que la susceptibilidad a oscurecerse fue similar entre los cladodios de los distintos estados de desarrollo. Sin embargo no se observaron diferencias la actividad específica de PPO en las cuatro variedades evaluadas. Los cladodios de COPENA F-1 mostraron mayor tendencia a oscurecerse y aunque presentaron el mayor contenido de ácido ascórbico, compuesto que reduce a las quinonas, también mostraron el mayor contenido de fenoles totales, los cuales son los sustratos de la enzima PPO. Los cladodios de Atlixco mostraron menor tendencia a oscurecerse y un menor contenido de ácido ascórbico y de compuestos fenólicos totales.
Conclusiones
La actividad específica de PPO de los cladodios de las cuatro variedades de Opuntia ficus indica aumentó significativamente al avanzar el desarrollo, alcanzando los máximos valores en el estado VI; y un comportamiento inverso ocurrió en el contenido de ácido ascórbico, que disminuyó al avanzar el estado de desarrollo. Se encontraron diferencias en el contenido ácido ascórbico total, de fenoles totales y oscurecimiento de las variedades COPENA F-1, COPENA V-1, Milpa Alta y Atlixco, presentando los cladodios de COPENA F-1 el mayor contenido de ácido ascórbico total, de fenoles totales y de oscurecimiento, mientras que los de Atlixco presentaron los menores valores. Estos resultados sugieren que desde el punto de vista de oscurecimiento los cladodios de Atlixco pudieran ser más adecuados para destinarse a la elaboración de nopal fresco cortado.

