










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 spe 12 Texcoco nov./dic. 2015
Articles
Histological and morphological study of autotetraploid and diploid plants of tomatillo
1Universidad Autónoma Agraria Antonio-Departamento de Fitomejoramiento. (godramf@gmail.com; MathGenome@gmail.com; bocardo_lety@hotmail.com; atorres_tapia@hotmail.com; hgosuna@hotmail.com).
2Universidad Autónoma Agraria Antonio-Departamento de Horticultura. Calzada Antonio Narro 1923, C.P. 25315. Saltillo, Coahuila, México.
Husk tomato (Physalis ixocarpa) is a species from Mexico, its importance lies in its high consumption in Mexico and export to the United States and Canada. Autopolyploids formation is an alternative to introduce new genetic potential in this species; however, the impact of artificial polyploidization on histology and morphology of plant is unknown, so it was proposed as objective to study histology and morphology of diploid plants (Rendidora) and autotetraploid. The plant material used were three autotetraploid (11, 16 and 20) selected based on yield and quality of fruit and rendidora diploid (19). The experiment was established in the field in General Cepeda, Coahuila in 2011, under a complete randomized block design with four replications, the useful plot were 10 plants. The morphological variables were: widths (AH), leaf length (LH), stem diameter (DT), flower diameter (DF) and plant height (AP). Histological were: leaf xylem vessels area (AXH), petiole (AXP), root (AXR) and stem (AXT), palisade parenchyma (LPEH) and lacunar parenchyma in leaf (APLH). Tetraploid showed leaf width and flower diameter significantly higher than diploid (p≤ 0.05); however in plant height diploid was significantly higher than autotetraploid. The area of xylem vessels of stem and root of autotetraploids showed significantly higher values (p< 0.01) than diploid according to Tukey test, also autotetraploid plants were more vigorous, an important characteristic in the improvement of this species, so this kind of studies provide data to understand the anatomical behavior of autotetraploids in tomatillo.
Keywords: Physalis ixocapa Brot; colchicine; histology; polyploidy; xylem vessels
El tomate de cáscara (Physalis ixocarpa) es una especie originaria de México, su importancia radica en su alto consumo en México y exportación a los Estados Unidos de América y Canadá. La formación de autopoliploides es una alternativa de introducción de nuevo potencial genético en esta especie; sin embargo, se desconoce el impacto de la poliploidización artificial sobre la histología y morfología de la planta, por lo que se planteó como objetivo; estudiar la histología y morfología de plantas diploides (Rendidora) y autotetraploides. El material vegetal utilizado fueron tres autotetraploides (11,16 y 20) seleccionados en base a rendimiento y calidad de fruto y el diploide rendidora (19). El experimento se estableció a cielo abierto en General Cepeda, Coahuila, en 2011, bajo un diseño experimental de bloques completos al azar con cuatro repeticiones, la parcela útil fueron 10 plantas. Las variables morfológicas fueron: ancho (AH) y largo de hoja (LH), diámetro de tallo (DT), diámetro de flores (DF), altura de planta (AP). Las histológicas fueron: área de vasos de xilema de hoja (AXH), pecíolo (AXP), raíz (AXR) y tallo(AXT), parénquima en empalizada (LPEH) y parénquima lagunar en hojas(APLH). Los tetraploides mostraron ancho de hoja y diámetro de flor significativamente mayor que el diploide (p≤ 0.05); sin embargo en altura de planta el diploide fue significativamente mayor que los autotetraploides. El área de vasos de xilema de tallo y raíz de los autotetraploides mostraron valores significativamente (p< 0.01) superiores a los diploides de acuerdo a la prueba de Tukey, además las plantas autotetraploides fueron más vigorosas, característica importante en el mejoramiento de esta especie, por lo que este tipo de estudios aportará datos para la comprensión del comportamiento anatómico de los autotetraploides en tomate de cáscara.
Palabras clave: Physalis ixocapa Brot.; histología; poliploidia; colchicina; vasos de xilema
Introduction
Physalis ixocarpa is a species from Mexico, known as tomatillo, green tomato, husk tomato, fresadilla tomato. 2014 was the fifth vegetable in acreage, with 46 524.96 ha and a national average yield of 14.94 t ha-1 (SIAP-SAGARPA, 2014). The importance of this vegetable is due to its high consumption in Mexico and export to the United States and Canada. Despite there is a wide genetic variability in both wild and domesticated tomato in Mexico (Santiaguillo et al., 2004), the national average yield is considered low.
Autopoliploidy is an inducible condition characterized by the duplication in the number of genomes from the same individual, which manages to achieve an increase in genetic variability, which can be exploited by plant breeders. Autopoliploidy increases the effective size of the population and increases genomic flexibility, facilitating the management of artificial selection. The genetic redundancy may allow adaptive divergence of duplicated genes (Parisod et al., 2010). In this regard Robledo et al. (2011) developed an autotetraploid population 2n= 4x= 48, using colchicine in Rendidora variety.
The new material is promising for new varieties or hybrids of high growth and quality (Robledo et al., 2011) and according to that mentioned by Tang et al. (2010) polyploidization is a major trend in plant evolution, which has many advantages over diploid and because the impact of artificial self polyploidisation at histological and morphological level in husk tomato is unknown; the following objective was proposed; to study histology and morphology if autotetraploid and diploid plants (Rendidora) of husk tomato previously formed by the action of colchicine, as the anatomical knowledge of artificial autotetraploid of husk tomato will provide data for basic understanding of the induction of autopoliploidy in this species.
Materials and methods
Field establishment
The experiment was carried out in 2011 in General Cepeda, Coahuila and in the Cytogenetics Laboratory from the Universidad Autonoma Agraria Antonio Narro in Saltillo Coahuila. The plant materials were three autotetraploid (11, 16 and 20) selected based on yield and quality of fruit and the diploid (19) rendidora. The populations were sown depositing two or three seeds per well in polystyrene trays of 200 cavities, using as substrate peat and perlite at a ratio of 1:1. The trays were carried to a greenhouse where seedlings developed and when these reached a height of 10 to 12 cm and two pairs of true leaves, the seedlings were transplanted in the field on grooves with padded black plastic, 5 m long and 1.8 m wide with a spacing of 60 cm between plants and two rows per groove with spacing of 30 cm between rows. Ten days after transplantation a thinned was performed leaving only one seedling per point. The treatments were established under a randomized block design with four replications, each treatment consisted of 10 plants. Nutrition is applied through irrigation, three times a week; irrigation was daily applying two liters per day per plant until 45 days after transplantation, then increased to four liters per day per plant.
Morphological characterization
At 60 days after transplantation, the following morphological variables were evaluated in diploid and tetraploids: leaf width (AH) and length (LH), stem diameter in cm (DT), flower diameter in cm (DF), plant height in cm (AP), these variables were measured in three plants taken randomly from each treatment in each of the four repetitions, making a total of 12 plants per each population. The measurements were made using a digital precision vernier AutoTEC®, plant height was measured with a tape measure in cm. These variables were evaluated in order to characterize tetraploid populations and to identify morphological changes induced by polyploidization in tomatillo plants.
Histological study
Samples were taken at 60 days after planting three plants per treatment of each replication and taken to the laboratory, cut a piece of tissue (2 cm) from the main root at 3 cm below soil surface, piece of tissue (2 cm) from stem at 10 cm above soil surface, in addition to the base of the first leaf a piece of tissue (approximately 1 cm2) taking as reference the midrib and petiole fragment (2 cm) from the first branch of each plant; the tissues were processed by the histological paraffin technique as follows: to preserve the tissues with minimal alterations were placed in glass jar 15 ml with fixative FAA (Formaldehyde (36-40%) 5cc, ethyl alcohol (70%) 90 cc and 5 cc glacial acetic acid). Then the samples were dehydrated in ethyl alcohol concentrations 50, 60, 70, 85 and 96%, plus eosin, continuing with absolute ethyl alcohol I, absolute ethyl alcohol II, absolute ethyl alcohol plus xylene in volume ratios of 3:1, absolute ethyl alcohol plus xylene in proportion 1:1, absolute ethyl alcohol plus xylene in proportions of 1:3 and finally the tissues passed to pure xylene and remained in each solution for 2 h and embedded in paraffin with melting point at 58 °C then the alcohol was replaced with xylene (Hernandez, 1990; Jauregui, 2003; Cañizares et al., 2005).
Transversal cuts of tissue at 20 microns with a rotary microtome were made, which were mounted on a slide. For staining a dual color staining of safranin-fast green were used, passing through different alcohol and xylene solutions, the preparations were pulled out from the last xylol and drained. Sealed with a drop of canada balsam and a slide of the tissue size.
Thus the preparations were put to dry in an oven at 30 °C for a week. The tissues were analyzed with the aid of a microscope with an integrated camera Pixera Wiender Pro; the best samples were selected and photographed with 10X and 40X objectives (Wilkinson, 1979). The measurements were made from digital images, with measurement software Axion Vision Rel. 4.8. The variables were xylem vessels area from the midrib of the leaf (AXH), petiole xylem vessels (AXP), xylem vessels from the main root (AXR), and stem xylem vessels (AXT) palisade parenchyma (LPEH) and lacunar parenchyma in leafs (APLH).
Statistical analysis
An analysis of variance was performed on a randomized block design with four replications, the significance for morphological variables: leaf width (AH) and length (LH), stem diameter (DT) flowers diameter (DF), plant height (AP), and histologic: xylem vessel area from the midrib of the leaf (AXH) petiole xylem vessels (AXP), xylem vessels from the main root (AXR), and stem xylem vessels (AXT) palisade parenchyma (LPEH) and lacunar parenchyma in leaf (APLH), was considered on the ploidy level, where the variation factor were three autotetraploid populations and one diploid population of tomatillo, for mean comparison Tukey test (p≤ 0.05). SAS statistical software version 9.0 was used.
Results and discussion
Morphological characterization
Diploid population had on average a width (AH) of 6.81 cm while the three autotetraploid populations had on average 8.51 cm, autotetraploid population 11 showed the highest value with 9.21 cm and was statistically similar (p> 0.05) to the other two tetraploid populations, but different (p< 0.05) from rendidora diploid (Table 1) observing that the leaves of autotetraploid plants showed a darker green color than diploid plants, also the life cycle of autotetraploid plants was longer (about 15 days) than diploid, these results agree with those reported by Chen et al. (2011) in tetraploid plants of Anthurium andraeanum "Arizona" induced with colchicine since tetraploid plants showed firm petioles, thick leaves and a dark green color, also had a longer cycle compared to diploid plants. Regarding to leaf length (LH) in this study, no significant differences (p> 0.05) were found between diploid and autotetraploid populations.
Table 1 Mean values of morphological characteristics of diploid and autotetraploid tomatillo.
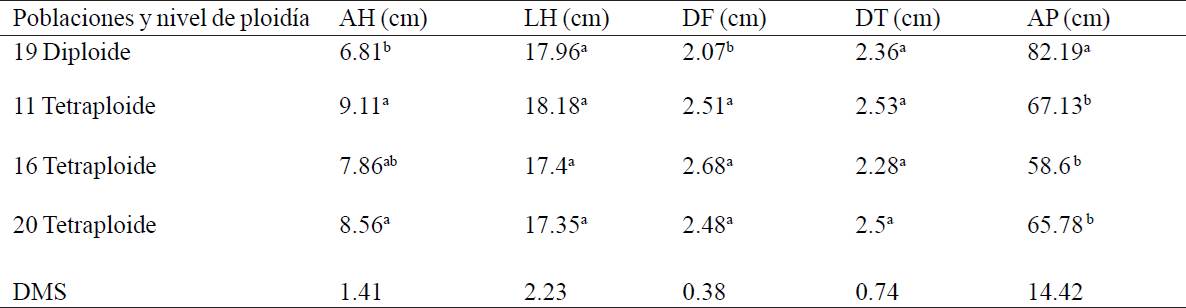
AH= ancho de hoja, LH=largo de hoja; DF= diámetro de flor; DT= diámetro de tallo; AP=altura de planta; *Letras diferentes dentro de las columnas indican diferencias significativas a nivel (P≤ 0.05); DMS= Diferencia mínima significativa.
In flower diameter (DF) significant differences between autotetraploid and diploid populations were found, the tetraploid populations showed between 2.47 and 2.68 cm, being tetraploid 16 which had the highest value and were statistically similar (p> 0.05) between them. The population that showed the smaller flower diameter was diploid 19 (2.07 cm), agreeing with Liu et al. (2007) who found morphological differences between diploid and tetraploid on Platanus acerifolia developed with the use of colchicine in Gerbera jamesonii Bolus cv. Sciella, the tetraploids showed slow growth, but had greater vigor, broad and thick leaves, larger flowers, longer stems and improved vase life (Gantait et al., 2011).
In stem diameter (DT), there were no significant differences between autotetraploid and diploid population, the tetraploid population 11 (2.53 cm), showed the highest value and was statistically similar (p> 0.05) to all others, including diploid population 19.
As for plant height (AP) diploid 19 (82.2 cm) had the highest value and was statistically different (p <0.05) to autotetraploid populations showing the lowest values (58.60 cm to 67.13cm) and were statistically equal (p> 0.05). Tomatillo autotetraploids had higher values for leaf width and flower diameter compared to diploid, coinciding with Cequea (2000) who mentions that autopolyploids have a wider ecological tolerance and larger cell sizes, meanwhile Imery and Cequea (2001) found that new tetraploid genotypes of A. vera produced greater leaf biomass accumulation, especially due to the increase in width and thickness of the leaves, also, Rego et al. (2011) found that with the formation of autotetraploid of Passiflora edulis Sims. (2n= 2x= 18) using colchicine and oryzalin obtained more vigorous plants (2n= 2x= 36).
In this research was noted that tetraploids growth was slower because at 60 days when diploid variables were taken outperformed tetraploids in plant height; however, at the end of the cycle tetraploid exceeded diploid, so it is recommend to take these variables at different stages of development and thus be able to match the observed, Molero and Matos (2008) who found that the gene duplication in Aloe vera increased plant height, length, width , thickness and leaf volume regarding to diploid plants, so it is estimated that polyploidization could be an alternative to increase production of cell biomass in this species. Leaf area is an important variable because it is directly related to the photosynthetic activity and this with the accumulation of reserves (Poehlman and Allen, 2005). A variable that normally is related to fruit size is flower size and in this case according to observed values it can be said that an increase was induced in autotetraploids regarding this characteristic.
Histological study
Histological variable cell area of lacunar parenchyma in leaf (APLH) the autotetraploid populations had on average 6 912.67µm2 being surpassed by diploid population showing a value of 7 398.97µm2, in xylem vessels area from the midrib leaf (AXH) the diploid showed 655.08 µm2 while autotetraploid presented 602.81µm2, length of palisade parenchyma cells (LPEH) autotetraploids exhibited 100.61µm with a value that exceeded the diploid, since this had 97 361µm (Figure 1), xylem vessels area in petioles (AXP) autotetraploid populations showed on average 604.54 µm2 while diploid plants showed a value of 714.56 µm2; however, no significant differences were found between diploid and autotetraploid plants since they were statistically equal (p≥ 0.05). The fact that in the present study no significant differences were found in these variables, may indicate that despite chromosomes and genes duplication, leaf and petioles characteristics were little affected by polyploidization in tomatillo, this may also be due to the fact that autotetraploid have a slow growth and the cycle is longer than diploids and for the present study diploid and autotetraploid samples were taken on the same date.
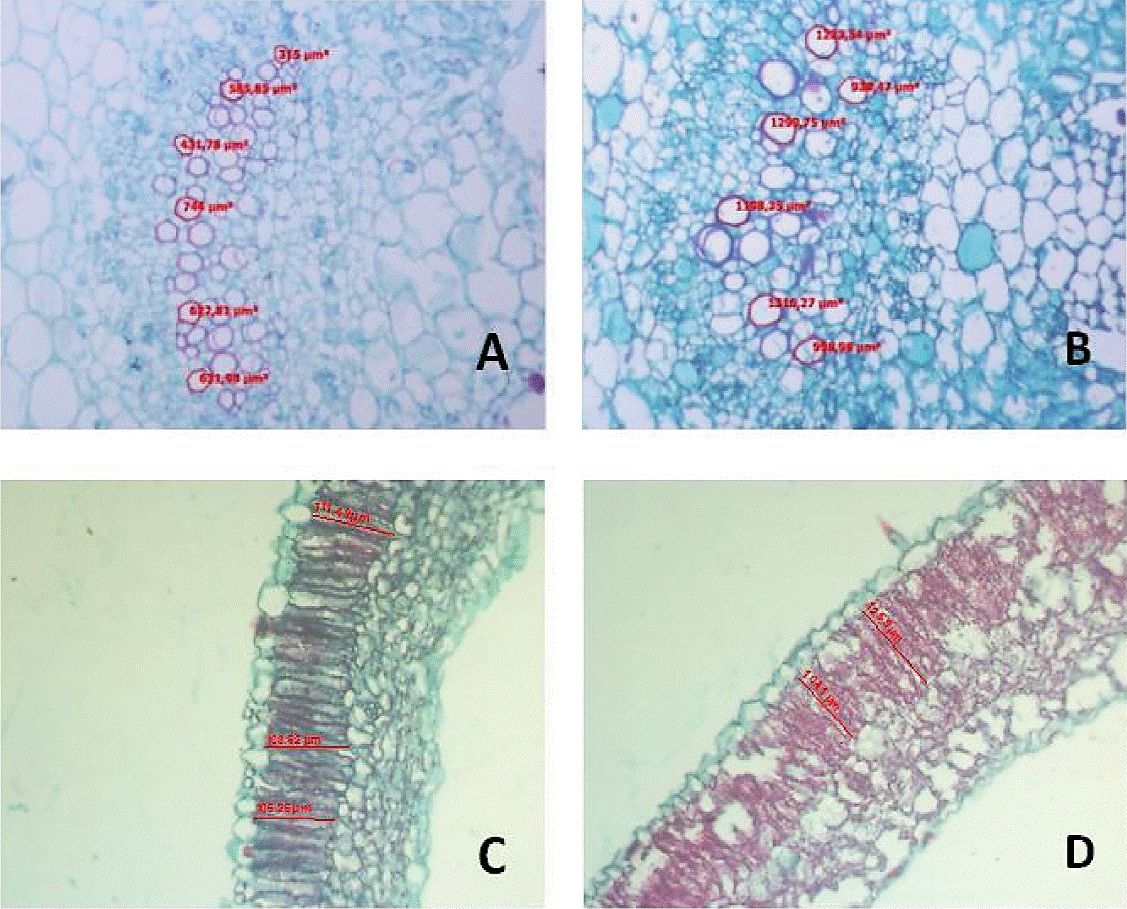
Figure 1 Xylem vessels of the midrib of the leaf; A) Diploid; autotetraploid. Palisade parenchyma of the leaf; Diploide; and D) autotetraploid in tomatillo 10 X.
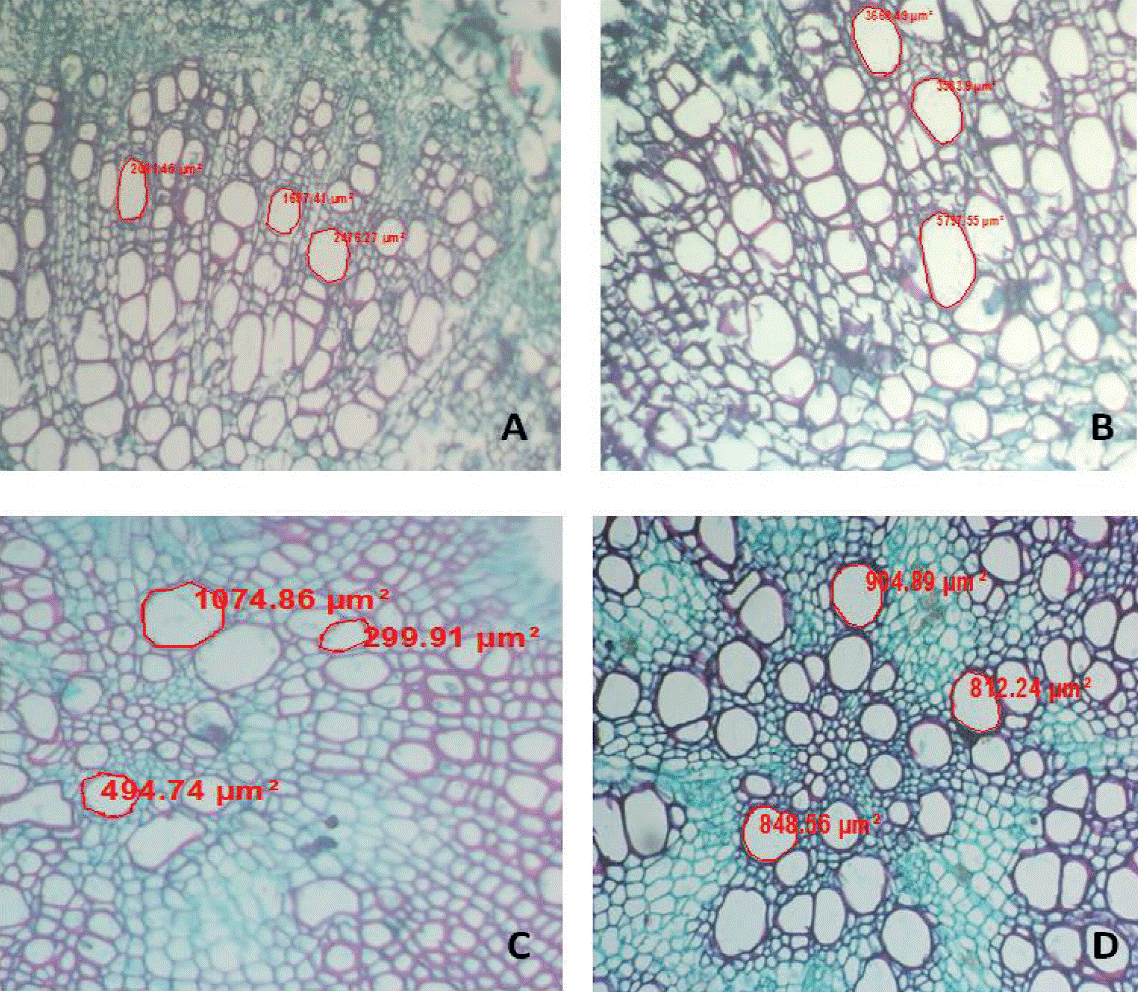
Variance analysis applied to diploid and autotetraploid plants presented significant differences (p< 0.01) between diploid and autotetraploid regarding to xylem vessels area from stem (AXT) where the three autotetraploid populations (1 842.66 µm2) exceeded the diploid (748.4 µm2) in 146.17%, for xylem vessels area in root (AXR) autotetraploid plants showed on average 1 309.68 µm2 while diploid had 738.4 µm2 and were excelled by tetraploids in 77.36%, therefore it was found that doubling the genome of tomatillo, increased xylem vessels area in stem and root (Figure 2 and 3), these results could be positive for this species according to Zahng and Cao (2009) who assume that it has been proven that the structure of the vascular system of the stem has a strong influence on leaf structure and functioning, assimilation of carbon dioxide, efficiency in the use of nutrients and growth rate.
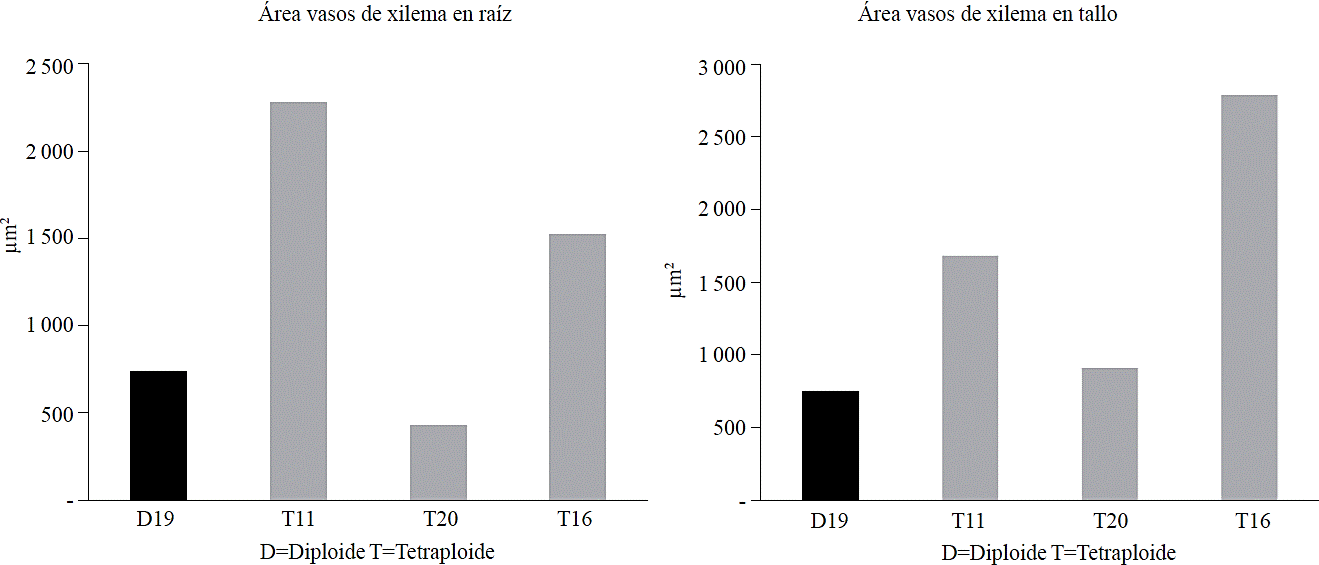
Figure 2 Area of xylem vessels in roots and stems of the diploid population and population autotetraploid tomatillo.
Conclusions
In autotetraploid tomatillo morphological variables such as leaf width and flower diameter were modified being higher than in diploid, giving the appearance of greater vigor in autotetraploid plants, also in autotetraploids th e xylem vessels area of stem and root was larger than diploid rendidora an important characteristic in water conduction and mineral salts.
Literatura citada
Cañizares, A.; Sanabria, M. y Rojas, E. 2005. Anatomía de la hoja de Lima Tahití (Citrus latifolia Tanaka). Revista UDO Agrícola. 5(1):68-73. [ Links ]
Cequea, A. C. 2000. Cytogenetic analysis of the artificial tetraploid Lycopersicon esculentum var cerasiforme. Ciencia. 8(2):119-126. [ Links ]
Chen, Ch.; Hou, X.; Zhang, H.; Wang, G. and Tian, L. 2011. Induction of Anthurium andraeanum “Arizona” tetraploid by colchicine in vitro. Euphytica. 181(2):137-14. [ Links ]
Gantait, S.; Mandal, N.; Bhattacharyya, S. and Das, P. K. 2011. Induction and identification of tetraploids using in vitro colchicine treatment of Gerbera jamesonii Bolus cv. Sciella. Plant Cell, Tissue Organ Culture. 106(3):485-493. [ Links ]
Hernández, S. M. 1990 Manual de laboratorio citología y citogenética. Editorial Trillas. Universidad Autónoma Agraria Antonio Narro. 105 p. [ Links ]
Imery, J. and Cequea, H. 2001. Colchicine-induce autotetraploid in Aloe vera L. Cytologia. 66:409-413. [ Links ]
Liu, G.; Li, Z. and Bao, M. 2007. Colchicine-induced chromosome doubling in Platanus acerifolia and its effect on plant morphology. Euphytica. 157(1-2):145-154. [ Links ]
Jáuregui, D. 2003. Manual práctico de microtecnia vegetal. Departamento de Botánica Agrícola. Laboratorio de Botánica. Facultad de Agronomía. Postgrado de Botánica Agrícola. Universidad Central de Venezuela. Maracay, Venezuela. 71 p. [ Links ]
Molero, P. y Matos, A. 2008. Efectos de la inducción artificial de la poliploidía en plantas de Aloe vera (L.). ReviCy HLUZ. Boletín Centro de Investigaciones Biológicas Venezuela. 42(1):111-133. [ Links ]
Parisod, C.; Holderegger, R. and Brochmann, C. 2010. Evolutionary consequences of autopolyploidy. New Phytologist. 186:5-17. [ Links ]
Poehlman, J. M. y Allen, S. D. 2005. Mejoramiento genético de las cosechas. Segunda edición. Limusa. México. 511 p. [ Links ]
Rêgo, M. M.; Rêgo, E. R.; Bruckner, C. H.; Finger, F. L. and Otoni, W. C. 2011. In vitro induction of autotetraploids from diploid yellow passion fruit mediated by colchicine and oryzalin. Plant Cell, Tissue Organ Culture. 107(3):451-459. [ Links ]
Robledo, T. V.; Ramírez, G. F.; Foroughbakhch, P. R.; Benavides, M. A.; Hernández, G. G. and Reyes, M. H. V. 2011. Development of tomatillo (Physalis ixocarpa Brot.) autotetraploids and their chromosome and phenotypic characterization. Breeding Sci. 61:288-293. [ Links ]
Santiaguillo, H. J. F.; Cervantes, S. T. y Peña, L. A. 2004. Selección para rendimiento y calidad de fruto de cruzas planta x planta entre variedades de tomate de cáscara. Rev. Fitotec. Mex. 27:85-91. [ Links ]
Statistical Analysis System (SAS) Institute. 2008. SAS user’s guide. Statistics. Version 8. SAS Inst., Cary, NC. USA. Quality, and elemental removal. J. Environ. Qual. 19:749-756. [ Links ]
SIAP-SAGARPA (Sistema de Información Agrícola y Pecuaria). 2014. Resumen nacional de avances agrícolas por estado. Producción anual. Cierre de la producción agrícola por cultivo. www.siap.gob.mx. [ Links ]
Tang, Z. Q.; Chen, D. L.; Song, Z. J.; He, Y. C. and Cai, D. T. 2010. In vitro induction and identification of tetraploid plants of Paulownia tomentosa. Plant Cell, Tissue Organ Culture. 102(2):213-220. [ Links ]
Wilkinson, H. 1979. The plant surface (maily leaf). In: C.R. Metcalfe y Chalk (edo). Anatomy of dicotiledons. Oxford Cloredons Press. 1:97-165. [ Links ]
Zhang, J. L. and Cao, K. F. 2009. Stem hydraulics mediates leaf water status, carbon gain, nutrient use efficiencies and plant growth rates across dipterocarp species. Func. Ecol. 23:658-667. [ Links ]
Received: May 2015; Accepted: August 2015
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
