











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkLa androesterilidad para realizar selección recurrente en trigo Male Sterile Facilited Recurrent Selection (MSFRS es una técnica que requiere una fuente de esterilidad que no esté ligada a efectos deletéreos, que de preferencia esté gobernada por un gen simple dominante y que sea efectiva en su incorporación a germoplasma elite base (Villaseñor et al., 2014). En especies autógamas poco se ha utilizado, porque de manera natural no hay germoplasma androestéril que permita la recombinación y el mejoramiento con la misma eficiencia que se logra en las alógamas (Sorrels y Fritz, 1982; Bockelman y Sharp, 1986). Para disponer de fuentes androestériles en trigo se han aislado mutantes inducidos mediante radiación recurrente o debido a mutaciones espontáneas, como por ejemplo, los reconocidos como Pugsley (Suneson, 1962); Probus (Fossati e Ingold, 1970); Cornerstone (Driscoll, 1977); Taigu-1 (Liu et al., 1986) y LZ (Zhou et al., 2008), los que en su mayoría están ligados a efectos deletéreos, por lo que han sido poco efectivos en el mejoramiento poblacional.
En cereales de grano pequeño han sido reconocidos los avances genéticos en el mejoramiento poblacional cuando se utiliza la MSFRS (Ramage, 1977). En México la técnica se ha utilizado por Villaseñor et al. (2002a), quienes reportaron avances genéticos para rendimiento de grano de 4.7% ciclo de selección, y por Solís et al. (2002) en donde indican que después de siete ciclos de selección recurrente, aumentó la frecuencia de genotipos de alto rendimiento y con resistencia a roya amarilla. Es importante indicar que el éxito del mejoramiento poblacional cuando se utiliza la MSFRS depende de la fuente de esterilidad, sin embargo, también es importante el germoplasma base a utilizar para formar las poblaciones (Ramage, 1977; Geraldi y de Souza, 1997; Villaseñor et al., 2002b), ya que éste deberá de contener los genes recombinantes de interés para lograr los avances genéticos del programa.
Una vez elegido el germoplasma base, se procede a incorporarles la fuente de androesterilidad para formar las poblaciones recombinantes (Ramage, 1977; Geraldi y de Souza, 1997). La retrocruza es una técnica de mejoramiento genético que ha sido efectiva para incorporar caracteres cualitativos como la androesterilidad (Márquez, 1988) y lo es aún más cuando el carácter a incorporar es dominante (Geraldi y de Souza, 1997; Villaseñor et al., 2014); entre mayor número de retrocruzas se realicen sobre el progenitor recurrente, mayor será la similitud. Villaseñor et al. (2014) realizaron cuatro retrocruzas en ocho variedades de trigo y evaluaron la similitud en seis caracteres cualitativos, encontrando entre 80% y 90% de efectividad. De acuerdo con lo anterior, el objetivo de la presente investigación fue valorar las ventajas que tiene el mutante androestéril dominante “Oly” en la reconversión e incorporación del carácter a material elite de trigo.
La fuente de androesterilidad “Oly”, debida a un gene simple dominante de trigo harinero (Triticum aestivum L.), se utilizó para incorporarla a 20 progenitores del programa de trigo de temporal del INIFAP. El proceso de reconversión se inició en ciclo primavera- verano de 2005 en el Campo Experimental Valle de México (CEVAMEX) ubicado en Chapingo, Texcoco, México y terminó en el ciclo Otoño-Invierno del 2009-10 en el Campo Experimental Bajío (CEBAJ) ubicado en Celaya, Gto. De la población recombinante identificada como PANDOLY, portadora del gen androestéril dominante que segrega en la proporción 1:1 de estériles y fértiles (Villaseñor et al., 2014), fueron sembradas 2000 semillas a una separación entre ellas de 15 cm (siembra espaciada); también fueron sembradas las semillas de los 20 progenitores a una separación de 15 cm en dos surcos separados a 30 cm y de 3 m de largo; antes de la floración, se cubrieron espigas de plantas androestériles para evitar que se polinizaran; posteriormente, grupos de cinco espigas cubiertas fueron elegidas para ser polinizadas con cada una de los 20 progenitores; la semilla recombinante se cosechó para obtener la F1 de la cruza (F1C). Para obtener la primera retrocruza, la semilla F1C se sembró espaciada a 15 cm en dos curcos separados a 30 cm y de 5 m de largo, junto con su respectivo progenitor (parcelas apareadas); antes de floración se cubrieron espigas de plantas androestériles de cada F1C; posteriormente se polinizaron con su progenitor y fue así como se obtuvo la semilla F1 de la retrocruza uno (F1RC1); este procedimiento se repitió hasta obtener la semilla F1 de la retrocruza seis (F1RC6); se tuvo especial cuidado en realizar la retrocruza hacia plantas androestériles cada vez más semejantes al progenitor.
La semilla de los 40 genotipos F1 de la retrocruza seis “F1RC6” de cada progenitor y los 20 progenitores) se sembró durante el ciclo primavera-verano de 2012 en el CEVAMEX espaciada a 15 cm en parcelas apareadas, cada una de dos surcos de 5 m y espaciados a 30 cm para dar un total aproximado de 60 plantas por parcela. El manejo agronómico del lote experimental se realizó bajo condiciones de temporal y de acuerdo a las sugerencias del INIFAP. Durante el espigamiento se cubrió una espiga en todas las plantas de las isolíneas androestériles para determinar la segregación fértil: estéril. En llenado de grano se evaluó la incidencia de roya amarilla (Puccinia striformis f.sp. tritici) en cada una de las parcelas y en madurez, en cada progenitor y su respectiva reconversión, se eligieron al azar 20 plantas en las que se midieron las variables altura de planta y longitud de espiga, que son las variables más visibles en el proceso de retrocruzamiento, y que en cierta forma son indicativas de la eficiencia de técnica MEFSR. Con los datos obtenidos se procedió a realizar un análisis de varianza general mediante el diseño completamente al azar con el paquete estadístico SAS (SAS-Institute 2002) obteniéndose así los promedios de cada genotipo por cada variable.
En total se tuvieron 556 plantas en las 20 isolíneas androestériles; se determinó la relación fértil:estéril, resultando que 268 fueron fértiles y 288 estériles, lo que de acuerdo con la Prueba de Ji Cuadrada (Ji2 tablas= 3.6 > Ji2 Calculada= 0.93) se ajusta a una relación 1:1, semejante a la segregación esperada para un gen simple dominante (Sun et al., 1994; Villaseñor et al., 2014). Todas las plantas fértiles lo fueron 100% y todas las plantas estériles lo fueron 100%, lo que confirma que la androesterilidad de mutante Oly se debe a un gen simple dominante (Villaseñor et al., 2014).
En el Cuadro 1 se presentan los promedios por variable para cada progenitor y su isolínea androestéril, en donde se puede apreciar que para Altura de Planta solamente en las variedades Norteña F2007, Eneida F94 y Gálvez M87 hubo diferencia significativas, mientras que en los restantes 17 grupos de genotipos no se detectó diferencia, información que indica que para este carácter la incorporación del gene androestéril fue efectivo; para las tres variedades en donde hubo diferencia significativa, las isolíneas androestériles fueron más altas hasta en 12 cm, lo que indica que en su momento las plantas androestériles retrocruzadas no fueron bien elegidas, ya que en poblaciones recombinantes androestériles el porte de planta alto es uno de los caracteres más modificados cuando no se hace la selección adecuada de las plantas recombinantes o cuando se deja al azar (Sun et al., 1994; Villaseñor et al., 2002b). En la variable longitud de espiga (Cuadro 1), sólo en las variedades Altiplano F2007 y Gálvez M87 hubo diferencia significativas, mientras que en los otros 18 grupos de genotipos no se detectó diferencia, información que también confirma que la incorporación del gene androestéril fue efectiva. También en el Cuadro 1 se presenta la reacción en el follaje a roya amarilla (Puccinia striformis f.sp. tritici) en los genotipos evaluados, en donde se observa que la respuesta en 15 progenitores y sus isolíneas androestériles respectivas fue prácticamente la misma en cuanto a porcentaje de infección y tipo de reacción, mientras que en los cinco grupos restantes varió ligeramente el tipo de reacción o el porcentaje de infección, información que permite determinar que para este carácter cualitativo también la incorporación del gene androestéril fue efectiva.
Cuadro 1 Variables agronómicas y fitopatológicas de progenitores y su reconversión androestéril después de seis retrocruzas. INIFAP-CEVAMEX, Chapingo, Estado de México, primavera-verano-2012.
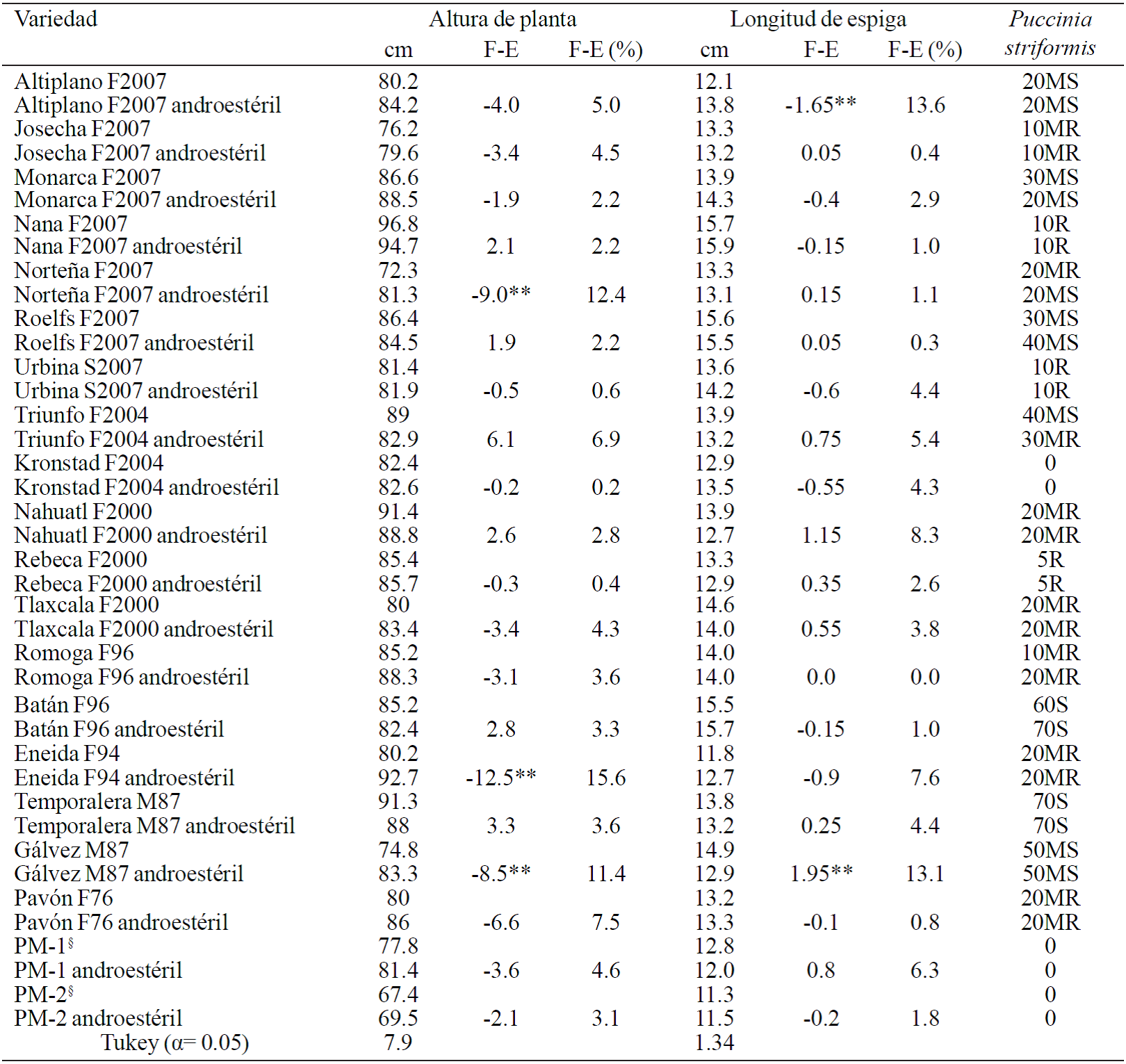
§líneas avanzadas; **= diferencia altamente significativas; F= fértiles; E= estériles; R= resistente; MR= moderadamente resistente; MS= moderadamente susceptible; S= susceptible.
Los resultados presentados indican que el esquema de retrocruzas aplicado al germoplasma elite del programa de trigo del INIFAP-CEVAMEX fue efectivo para incorporar el gen estéril dominante “Oly”, ya que se logró gran similitud entre los progenitores y sus isolíneas androestériles. Esta reconversión hacia androesterilidad es muy importante, ya que de acuerdo con Geraldi y de Souza (1997), el germoplasma elite de un programa de mejoramiento genético es el portador de los genes favorables que van a permitir los avances a mediano y largo plazo. Por otra parte, la referida reconversión permitirá la formación de poblaciones recombinantes para realizar selección recurrente, tal y como lo reportan Sun et al. (1994) cuando evaluaron el efecto del mejoramiento utilizando el gen mutante dominante Taigu-1, quienes concluyeron que la reconversión a androestériles del su germoplasma más sobresaliente permitió lograr mayores avances.
Conclusiones
El mutante androestéril “Oly”, después de seis retrocruzas y tomando como referencia caracteres agronómicos y fitopatológicos altamente influenciados por el ambiente, fue adecuado para incorporarle la fuente de androesterilidad a germoplasma elite de trigo, no se asoció con efectos deletéreos y segregó en la proporción 1:1 para plantas fértiles y estériles. La reconversión a androestériles del germoplasma elite considerado es una labor muy importante, ya que será la base para la formación de poblaciones recombinantes que permitirán realizar selección recurrente con la misma eficiencia que se logra con la MSFRS

