










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 no.spe11 Texcoco Mai./Jun. 2015
https://doi.org/10.29312/remexca.v0i11.780
Investigation notes
Influence of inbreeding on the theoretical answer to long term mass selection in maize
1Universidad Autónoma Chapingo-Centro Regional Universitario Occidente, Guadalajara, Jalisco. México.
The theoretical influence of inbreeding occurs in mass selection in maize. With prior knowledge of the value of inbreeding, the influence of inbreeding is calculated from a formula that allows to separate the additive effects of non-additive effects in this research dominance. The yield reduction by inbreeding after a hundred rounds of selection was 11.76%.
Keywords: Zea mays L.; inbreeding influence; mass selection
Se presenta la influencia teórica de la endogamia en la selección masal en el maíz. Con el conocimiento previo del valor de la endogamia, la influencia de la endogamia se calculó a partir de una fórmula que permite separar los efectos aditivos de los efectos no aditivos, en esta investigación la dominancia. La reducción del rendimiento por la endogamia al cabo de cien ciclos de selección fue de 11.76%.
Palabras clave: Zea mays L.; influencia de la endogamia; selección masal
The mass selection in maize is used for two main purposes: to purify landraces and increase yield for a few rounds of selection, to be subsequently improved by a more efficient method and the modified ear-to-row selection. The practical reason for the change of mass selection by another method is its low response 2%, found in Chapter 7 of Hallauer and Miranda (1981) in which only the data were considered more than five cycles of selection.
As in any method of selective breeding, in each cycle as the best plants are selected eventually as inbreeding is generated after each cycle the selected plants are increasingly related.
Therefore, the purpose of this paper was to show how much reduction is caused by inbreeding through cycles, to be able to suspend the selection when the expected response does not justify additional cycles.
Background
Busbice (1970) showed that the genotypic value is:
Where: F= coefficient of inbreeding; A= additive component; and B= heterozygous component. When B= 1 there is additivity only in the genotype. Thus the overall additivity is manifested, for example, in the lines obtained over many self-fecundations. In practice the selection of the additive increases the response (R) to the selection, since as stated, after each cycle plants are increasingly related.
Additivity (A) in the selection cycle t is equal to the sum of the additive and the response (R) to the selection in the immediately preceding cycle:
On the right side of Equation [1] to the term (1 - F) B is called heterozygosity and varies from zero to B. Heterozygosity was defined by Falconer (1964) as "the frequency of heterozygous at any time in relation frequency in the population base”. Therefore, as the selection progresses inbreeding increases while heterozygosity is reduced.
In Hallauer et al. (2010), the reduction in yield in the BSSS population varied from 0-100% of homozygosity, which caused the reduction in the yield of 70 quintals per hectare equal to 20/70 = 28%, which meant a heterozygosity of 72%. In this manuscript were taken as a basis the respective approaches 30 and 70%.
The coefficient of inbreeding (Márquez- Sánchez, 2008) in cycle t selection for families n and m plants per family was calculated as:
Equation [2] is based on the constitution of a maize variety, as a population of sib families.
For the calculation of the response to the selection, the numbers n and m of Equation [2] must be adjusted for the actual number of variance according to Crossa and Venkovsky (1997).
Without the influence of genotypic mean inbreeding in the selection t cycle is
Response to selection
Márquez- Sánchez (2008) adaptation of the response equation for mass selection is a gene locus (Empig et al., 1971):
Where= i is the selection intensity; u= the semidifference between the genotypic values of homozygous dominant and the recessive homozygote; p= is the gene frequency and σ is the standard deviation P phenotype. Considering each a k loci, everyone with a frequency of 0.5 (Goodrich et al., 1975) determining the character, then ku= U. Therefore, in cycle t of selection the answer is:
The mean and phenotypic variance at the plant level were calculated with data from the doctoral thesis of Márquez-Sánchez (1969), with mean Y= 150 g, and σ 2 P= 228675.24 g (σ P= 478.2 g). The response to selection depends on the selection pressure in this study is: s= 5%, which corresponds to a selection intensity i= 2.08; also depends on the percentage value of the response in the cycle 0, approximately equal to 2, as already mentioned. With this information the value of U2, which also assumed complete dominance obtained as Gardner and Lonnquist (1959) found that there were approximately complete dominance, in a population derived from several generations of genetic recombination of the original cross. To calculate U2, the percentage response selection cycle 0 was used, that is:
Where: U2= 2759.0012; knowing i, U2, σP and Y= the absolute response in the cycle depends only on pt, and it is:
Inbreeding influence
Inbreeding is calculated using Equation [2], and as mentioned, the values of n and m must be set by the effective number of variance by:
As mass selection generally used n= 200 and m= 20, nm= 4 000, is obtained:
Adjusted numbers are n*= 85 280 and m*= 8528 according to Márquez-Sánchez (2010), so the working equation of Equation [2] is:
Finally, affected by genetic inbreeding half cycle t is calculated from the equation G= A + (1 - F) B, as:
The Table 1 shows yields at 100 cycles of selection in each tenth cycle, are also shown: the gene frequency (pt), the response to selection (Rt) calculated with Equation [5], additivity (At) calculated with Equation [1], and St calculated with Equation [3], F t calculated with Equation [6] and
Table 1 Parameters used for calculating the genotypic means without the effect of inbreeding (

It is noted that, the value of Rt is reduced to zero in the cycle 100,
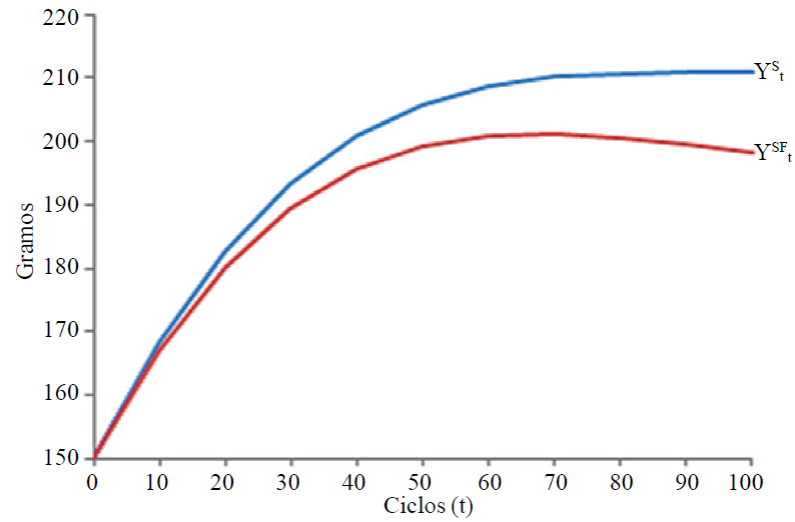
It is expected that, the effect of inbreeding also manifest in other characters that were not included in this research.
Literatura citada
Busbice, T. 1970. Predicting yield of synthetic varieties. Crop Sci. 10:265-269. [ Links ]
Crossa, J. R. and Venkovsky, R. 1997. Variance effective population size for two-stage sampling of monoeciuos species. Crop Sci. 37:14-26. [ Links ]
Empig, L. T.; Gardner, C. O. and Compton, W. A. 1971. Theoretical gains for different improvement procedures. University of Nebraska, College of Agriculture. 22 p. [ Links ]
Falconer, D. S. 1961. Introduction to quantitative genetics. Edinburgh and London. 365 p. [ Links ]
Gardner, C. O. and Lonnquist, J. H. 1959. Linkage and degree of dominance of genes controlling quantitative characters in maize. Agron. J. 51.524-528. [ Links ]
Goodrich, C. L.; Stucker, R. E. and Compton, W. A. 1975. Average gene frequency estimates in two open-pollinated cultivars of corn. Crop Sci. 6:746-749. [ Links ]
Hallauer, A. R. and Miranda, Fo. J. B. 1981. Quantitative genetics in maize breeding. 468 p. [ Links ]
Hallauer, A. R.; Carena, M. J. and Miranda, Fo. J. B. 2010. Quantitative genetics in maize breeding. Springer, New York. 663 p. [ Links ]
Márquez-Sánchez, F. 1969. Influence of half-sib family size on the estimation of genetic variances in maize. Ph. D. thesis, Iowa State University, Ames, Iowa. 205 p. [ Links ]
Márquez-Sánchez, F. 2008. A corrected calculation of inbreeding in mass selection. Maydica. 53:227-229. [ Links ]
Márquez-Sánchez, F. 2010. A proposal to long term response to maize selection. Maydica . 55:135-137. [ Links ]
Received: January 01, 2015; Accepted: April 01, 2015
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
