










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 no.8 Texcoco nov./dic. 2015
Articles
Boron fertilization and its relation to the production of seedless fruit in mango ʻAtaulfoʼ
1Campo Experimental San Luis-INIFAP. Carretera San Luis-Matehuala, km 14.5, Soledad de Graciano Sánchez, San Luis Potosí, México. C. P. 78430. Tel. 01 (444) 8524316. (hernandez.alfredo@inifap.gob.mx).
2Departamento de Fitotecnia, Universidad Autónoma Chapingo. Carretera México-Texcoco km 38.5, Chapingo, Estado de México, México. C. P. 56220. Tel: 01 (595) 9521500.
3Campo Experimental Santiago Ixcuintla-INIFAP. Entronque Santiago Ixcuintla km 5, carretera Internacional. Santiago Ixcuintla, Nayarit, México. C. P. 63300. Tel: 01 (323) 2352031. (hipeba60@hotmail.com; osuna.jorgealberto@inifap.gob.mx).
4Programa de Edafología, Colegio de Postgraduados Campus Montecillo. Carretera México-Texcoco, km 36.5. Montecillo, Estado de México, México. C. P. 56230. Tel: 01 (595) 9520200. (tlibia@colpos.mx).
5Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5 Chapingo, Estado de México, México. C. P. 56220. Tel: 01 (595) 9542200. (rosgar08@ hotmail.com).
Mexico is the fourth largest producer of mango (Mangifera indica L.) in the world. The Ataulfo cultivar has commercial importance because of its larger shelf life compared to other cultivars; but it has the disadvantage of producing seedless fruits that have no value for export. In order to determine whether the application of boron to soil can reduce the formation of seedless fruit and, improve the nutritional status of the tree, soil applications of boron were made (Solubor) 25, 50 and 100 g of B per tree, after three months after harvest in an orchard eight years old in Tepic, Nayarit. The percentage of seedless fruit, the nutrient concentration in leaves, inflorescences and fruits were evaluated. The nutritional analysis foliar before application of said treatments trees presented nitrogen deficiency. Doses of 50 and 100 g of B decreased by 45 and 35%, respectively, the number of seedless fruit and improved concentration of Ca and B on the plant, and that of B in shoots with inflorescences and leaves at the end of fruit development. The inflorescences concentrated N, P, K and Mg. Ca and B were concentrated in the leaves. The seeds accumulated more N, P, K, Ca and Mg than the pulp with epidermis. The fruit with seeds outscored the seedless in the concentration of N, P and Mg, while the latter concentrated more Ca.
Keywords: Mangifera indica L.; nutrient concentration; pulp with epidermis; seed
México es el cuarto productor de mango (Mangifera indica L.) en el mundo. El cultivar Ataulfo tiene importancia comercial por presentar mayor vida en anaquel que la de otros cultivares; pero presenta la desventaja de producir frutos sin semilla que carecen de valor para la exportación. Para determinar si aplicaciones de boro al suelo pueden disminuir la formación de frutos sin semilla y mejorar el estado nutrimental del árbol, se hicieron aplicaciones al suelo de boro (Solubor) de 25, 50 y 100 g de B por árbol, a los tres meses después de la cosecha, en una huerta de ocho años de edad en Tepic, Nayarit. Se evaluó el porcentaje de frutos sin semilla, la concentración nutrimental en hojas, inflorescencias y frutos. El análisis nutrimental foliar antes de la aplicación de los tratamientos indicó que los árboles presentaron deficiencia de nitrógeno. Las dosis de 50 y 100 g de B, disminuyeron en 45 y 35%, respectivamente, el número de frutos sin semilla y mejoraron la concentración de Ca y B en la planta, así como la de B en brotes con inflorescencia y de hojas al final del desarrollo del fruto. Las inflorescencias concentraron N, P, K y Mg. El Ca y B se concentraron en las hojas. La semilla acumuló más N, P, K, Ca y Mg que la pulpa con epidermis. Los frutos con semilla superaron a los sin semilla en la concentración de N, P y Mg, en tanto que éstos últimos concentraron más Ca.
Palabras clave: Mangifera indica L.; concentración nutrimental; pulpa con epidermis; semilla
Introduction
Mango (Mangifera indica L.) is one of the most important tropical fruit in the country, is grown on about 185 hectares with a production of approximately 1.5 million tons (SIAP, 2011). Mexico is the fourth largest producer, second only to India, Thailand and China. The export of this fruit is mainly based on Florida cultivars Tommy Atkins, Haden, Kent and Keitt; in recent years cultivar Ataulfo joint this group, an originating material of Mexico, which has a wide acceptance in international markets for having some advantages such as longer shelf life than the cultivars Florida. The fruit of the cv. Ataulfo has export quality, which represents 13% of annual export volume (Vázquez-Valdivia et al., 2006).
One of the most important problems of the mango ʻAtaulfoʼ is the presence of a large number of fruits that are small and have no export value; these mangos are known technically as seedless fruit and its size producers call them "child"; said fruits lack seeds and many of them have a longitudinal crack, some fruits fall and stick to the tree until harvest.
The presence of this type of fruit is variable within the gardens, some of them is minimal, but sometimes can reach up to 80% of the fruit (Pérez et al., 2007). The causes of the formation of these fruits and how to avoid, so far unknown. Singh (2005) observed reduction in size and weight of the fruit as a result of the abortion of the embryo in the Glenn, Irwin, Haden, Kent and Kensington cultivars. Problems in pollination and fertilization lead to the abortion of embryos and the formation of small seedless fruits (Beasley et al., 1999), which can fall in early stages of development or reach harvest.
Regarding the nutritional aspects, it is well known that boron (B) is necessary for the production and germination of pollen grains and pollen tube growth (Lovatt and Dugger, 1984; Delgado et al., 1994; Marschner, 1995), plays an important role in seed production, as under moderate or severe deficiency of this nutrient, plants stop producing flowers and functional can stop producing seed (Gupta, 2007). On flowers generally of low concentrations of B reduced male fertility upon deterioration of micro-sporogenesis involvement and pollen tube growth. Post-fertilization effects, including deterioration of embryogenesis, resulting in seed abortion or incomplete formation of malformed or damaged embryos and fruits (Dell and Huang, 1997).
B is also important for the growth of the fruit, for their effect on cell division and expansion process (Díaz, 2002); some mango cultivars, such as Tommy Atkins, Haden 2H and Van Dyke are sensitive to low concentration of boron, presenting sharp drop in young fruit, seed abortion and hence low production; When these cultivars were applied with Borax, at doses of 300 g per plant in the first year and 100 g of the following year, all significantly increased production (Rossetto et al., 2000). Xue-Qun et al. (2011) found that the spray borate (K2B4O7) at doses of 15 mM, to mango trees in bloom, significantly decreased the percentage off lowers with anthracnose (Colletotrichum gloeosporioides) and the number of fruits per branch increased.
Therefore, this study was proposed with the objectives for: a) determining whether the application B to the soil can reduce the formation of seedless fruit; b) whether the applied B improves and B macronutrient content in leaves, flowers, fruits with seeds and seedless; c) determining the distribution of macronutrients and B in the fruit; and d) comparing the nutrient concentration between fruit with seed and without seed.
Materials and methods
This study was conducted in a mango orchard of ʻAtaulfoʼ, with seedless fruits, located in the town of Atonalisco, in Tepic, Nayarit, Mexico; which is among the geographical coordinates 21° 37' north latitude and 104° 48' west longitude at an elevation of 601 m. The climate of the region is Aw2(w) (i') corresponding to a warm humid with rains in summer (García, 1973). The characteristics of the soil were (0-40 cm depth): pH 6.0, electrical conductivity of 0.4 dS m-1, cation exchange capacity 12 cmol c kg-1, organic matter content of 3.5%, texture franco-sandy-clay and poor (Castellanos et al., 2000) on nutrient content (N 4.9, P 6.7, 184 K, 423 Ca, Mg 183 and B 0.27 mg kg-1). The orchard had a density of 156 trees ha-1 of 8 years old (real-frame 8 m), grafted on landrace mango. Two applications (the beginning and end of the rainy season) were performed with 3 kg per tree of 17-17-17 fertilizer formula.
The experiment consisted of the administration of three doses of boron down 25, 50 and 100 g per tree. The commercial source used was Solubor (20.5% B). To which randomly selected 5 trees for each treatment, including a control treatment (without fertilization). The application was made three months after harvest, equally in the four corners of the cup and drip area of the tree. The experimental design was completely randomized, with five repetitions; the experimental unit was a tree.
Before the application of boron, we made a foliar sampling on the vegetative buds, in order to meet the initial nutritional status of the trees; to evaluate the effect of treatments during flowering were sampled leaf shoots with inflorescences and buds without inflorescences and inflorescence; Finally, two weeks before harvest samples of leaves, fruits and seedless fruit with seed were performed. For complete foliar samplings 40 healthy leaves (leaf with petiole), mature tree harvested by the middle of outbreaks around the tip, similar to Oosthuyse (2000).
Four inflorescences per tree were sampled (one per cardinal point of the tip) and from each inflorescence 6 minor shafts were collected (2 baseline, 2 intermediate and 2 apical). For the sampling of fruits, those at physiological maturity (epidermis light yellowish green) from the four corners of the tip were selected; 8 seedless fruit and 4 fruit seed (also called standard) were collected per tree. In the laboratory the normal fruits were cut into pulp with epidermis and seed, to make the nutritional analysis separately.
The concentration of N, P, K, Ca, Mg and B was evaluated on samples of each body; to which 0.5 g of dry sample underwent digestion with a diacid wet mixture (H2SO4:HClO4, 2:1 v/v) and hydrogen peroxide. The determination of N was made by the microkjeldahl method, the other elements were determined in an atomic emission spectrophotometer by inductively coupled plasma (ICP) AES of Varian Australia Pty Ltd. (Victoria, Australia) in all the cases was followed the methodology described by Alcántar and Sandoval (1999). Finally, the percentage of fruits and pitted were calculated by the total tree fruits per treatment.
In order to analyse the results, an analysis of variance and the comparison test with Tukey package SAS version 9.0 (SAS Institute, 1999) were performed.
Results and discussion
Nutrient concentration. In the Table 1 is shown the concentrations of the elements evaluated in the leaves before application of boron. According to Agustí (2004), the concentration of N sufficiency on the leaf of mango is between 1 and 1.5%, implying that trees ʻAtaulfoʼ used in this research N concentration was low, which is related to the very low content of this element in the soil of the orchard as demonstrated by the analysis (4.9 mg kg-1); while the foliar concentrations of the other elements, even though in low concentrations, according to the interpretation of soil test by Castellanos et al. (2000), were found within the ranges of sufficiency.
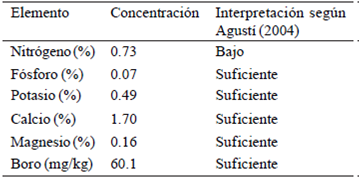
Nitrogen is one of the main structural elements of plants and plays an important role in the vegetative and reproductive growth. Zong-min et al. (2012), stated that this element represents 2% of the dry matter of the plants and is a component of proteins, nucleic acids, coenzymes and numerous secondary metabolites (Miller and Cramer, 2004), its failure reduces plant growth and leaf area with the consequent decrease in the rate of photosynthesis. Proper management of N in horticultural plants leads to obtain high-quality flowers, fruits and seeds.
In fruit species, nitrogen deficiency may induce abortion of pollen; if properly supply the N improves the longevity of the egg (Díaz, 2002) which leads to improve the fruit's grip.
In mango it has been observed that the mismanagement of N supply to the tree affects the production of flowers, fruits number and quality; the excess of this element in the soil promotes a vigorous vegetative growth at the expense of the reproductive (Medeiros et al., 2004).
When analysing the effect of treatments on the nutrient concentration in leaves it was observed that the application of B, only influenced the concentration of Ca and B; the highest concentration of Ca was obtained with the dose of 100 g of B; it was observed that all three doses of boron into the soil improved the concentration of B in the plant (Table 2). This means that the B applied was absorbed and transported via xylem to the tree, with improvement in the absorption of Ca, which was in very low in concentration (Castellanos et al., 2000) on the soil of the orchard (423 mg kg-1 of soil).
Table 2 Nutrient concentration in mango leaves ʻAtaulfoʼ with applications of boron into the soil in Nayarit, Mexico.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa. NS= no significativo.
Fageria (2001) notes that the B helps in the translocation of Ca absorbed into the aerial part of the plants. However, this does not match Marschner (1995) and Castellanos et al. (2000), who pointed out that at soil level and reactions on the surface of B reduces root using Ca. However, none of the treatments B improved foliar N concentration, which does not coincide with Marschner(1995) who indicates that B can have a positive indirect effect on the absorption and assimilation of nitrate, because it leads to an increase in activity nitrate reductase, as observed by Bellaloui et al. (2010) in soybean [Glycine max (L.) Merr.].
The treatments had only effect on the concentration of B in leaves and shoots with inflorescences leaves before harvest of the fruits harvested. In both types of leaves, the concentration of B at 100 g of the soil doubled the concentration of boron, compared with the concentration of the leaves of the control (Table 3). This means that B was transported to that kind of leaves because there were demand points of the element, such as flowers and growing fruits. In olive trees Delgado et al. (1994) found higher concentration of B in young leaves of trees in bloom in the trees that were not in bloom, indicating a higher need for B by trees during the reproductive process.
Table 3 Concentration of boron in mango leaves ʻAtaulfoʼ applications of boron to the soil in Nayarit, Mexico.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
The initial N concentration (before application of treatments), decreased 50% during flowering, as indicated concentrations in the leaves of the vegetative and reproductive buds; inflorescences concentration was equal to that of the leaves before applying the (initial) treatment and leaves before harvest; normal fruits were those with the highest concentration of N, seedless fruits had the same concentration as the leaves of vegetative and floral buds (Table 4). The highest concentration of Pand K was observed in normal fruits, followed by inflorescence; P concentration leaves the various stages and seedless fruit was similar; K the leaves of all samples was similar; while the seedless fruit was 50% lower than normal fruit.
Table 4 Nutrient concentrations (mg g1 dry matter) in leaves, inflorescences and fruits of mango ʻAtaulfoʼin Nayarit, Mexico.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
The initial foliar concentrations of Ca and leaves when the harvest of fruits was similar and higher than the rest of the organs; the lowest concentration of this element was recorded in the inflorescences and the two types of fruit. The highest concentration of Mg was observed in vegetative buds leaves and inflorescences followed by leaves and floral buds of normal fruit; with the lowest concentration the seedless fruits. The initial concentration of B was increased and decreased in the rest of the structures tested, including normal fruit; seedless fruits had the lowest concentration of this element (Table 4).
These results indicate that, the N, P, K and Mg were transported to the points of demand, as were the normal inflorescences and fruits; as their concentrations were equal to or exceeded the initial foliar concentrations (Table 4), because they are all moving elements in phloem (Marschner, 1995). In the case of Ca and B, the results indicate that both elements are concentrated in the leaves, mainly before flowering (baseline); while inflorescences and fruits (demand points) of the two types received very little input; This is because the transport of them is in the xylem and perspiration depends on the organ, and the leaves are the structures with more transpiration rate; also Ca and B are considered of very low mobility in the phloem (Epstein and Bloom, 2005; Fageria, 2009), so that most of the Ca and B demanded by points growth as shoot apices, young leaves or fruit must be covered by transport via xylem (Marschner, 1995). Medeiros et al. (2004) when evaluating the nutritional content of the leaves in four phenological stages of the cultivar Tommy Atkins, found that Ca had the highest nutrient concentration in leaves but not in the flowers and fruits.
Percentage of fruits with and without seeds. Doses of 50 g of B per tree cut in half the percentage of seedless fruit; whereas the dose of 100 g showed 35% less of these fruits (Table 5). This is because the B is essential for fruit production with seed micronutrient. In the flowers, the low concentration of B reduces male fertility due the impairment of microsporogenesis and pollen tube growth by promoting the formation of callose in the style. The post-fertilization effects include the deterioration of embryogenesis, resulting in seed abortion or incomplete formation of malformed or damaged embryos and fruits (Dell and Huang, 1997; Díaz, 2002).
Table 5 Percentage of fruit with and without seeds in trees of mango ʻAtaulfoʼ with applications of boron to the soil in Nayarit, Mexico.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
In flowers of grapevine (Vitis vinifera L.), whose stigmas containing from 8 to 20 mg of B per g of dry matter had poor fertilization compared to those where stigma containing 50 to 60 mg of B (Díaz, 2002). Rossetto et al. (2000) found that mango cultivars have different behaviour in relation to the level of boron in the leaf. The mango ʻWinterʼ had good production and retention of normal fruit under low boron concentration in leaves (10 mg kg-1 ms); while the mangos ʻHaden 2Hʼ and ʻVan Dykeʼ were sensitive to this concentration, and consequently fall had intensive young fruit, seed abortion and low production.
With the results obtained in this study on ʻAtaulfoʼ mango, concentrations of B kg-1 ms corresponding to 60 mg in the leaves before flowering (baseline); in bloom, 41 mg shoots without leaves inflorescences and 48 mg in shoots with inflorescences leaves (Table 4). Treatment with 100 mg of B, leaf buds productive concentration was increased to 63 mg kg-1 ms (Table 3); indicating that in ʻAtaulfoʼ flowers require more B than other cultivars and that doses of 50 and 100 g of B reached concentrations in production outbreaks that could prevent abortion of the embryo.
Nutrient concentration in fruits. The concentration of N, P and Mg was significantly higher in the fruits with seeds in seedless fruits. K concentration was similar in both types of fruits and Ca was higher in seedless fruit. In fruits with seeds, this was the structure that concentrated more N, P, K, Ca and Mg pulp with epidermis. B concentration was similar between the two types of fruits and between the seed and pulp with epidermis of normal fruit (Figure 1).

Figure 1 Nutrient concentration in fruit without seed (FSS), pulp with epidermis (PCE), seed (SEM) and fruit with complete seed (FCS). Means with the same letter are statistically equal according to the mean test Tukey (p≤ 0.05).
The concentration of N, P, K, Mg and B of seedless fruit was similar to that found in the pulp of normal fruit; only the concentration of Ca in the seedless fruit exceeded the seed and pulp with epidermis (1.2 mg) (Figure 1).
Nutrient distribution in the fruit varies by cultivar and especially between the pulp and skin, as in mango 'Manila' Guzmán et al. (1996) found that the skin of the fruit was the richest tissue in Mg, Fe and Mn, the pulp in K, the bone in CA and the seed in N, P, Cu and Zn; so the nutritional fruit extraction was in the following order: K> N> Ca> Mg> P. Moreover, Singh (2005) indicates that found no significant differences between the concentrations of N, K and Mg neither in the epidermis and the pulp, nor in those of P and Ca in seedless fruit pulp and fruit with seed in five cultivars evaluated. In the case of cv. Ataulfo, it was observed that the seed which was concentrated again all evaluated items in comparison with the pulp with epidermis.
As mentioned, there is a high positive correlation between the distribution of Ca and transpiration rate of organs, which explains the low Ca content (<0.3%) in fruits (structure with little perspiration), compared with the leaves (3 to 5%) in the same plant (Marschner, 1995); therefore, in the mango fruit ʻAtaulfoʼ low concentration of Ca, and much higher in the leaves. The distribution B is also related to the loss of body water, so the leaves accumulate more boron than the seeds and fruit tissues, as observed in this study with mango ʻAtaulfoʼ (Table 4). Although, studies on olive trees (Olea europea L.) have shown that flowers and fruits, in their early stages of development, can promote the mobilization of leaf B to meet its demand for this element (Delgado et al., 1994).
In the case of seedless fruits, the absences of the embryo in the seed makes the demand, by the elements decrease, and therefore are less concentrated. The presence of seeds represents an important fruit growth factor because it is the regulatory body of their development (Díaz, 2002) and affects the accumulation of nutrients, particularly Ca in pulp and epidermis (Singh, 2005). During seed development, the embryo produces plant hormones such as auxins and gibberellins, which stimulate the development of seed and fruit tissues, such as pulp (Díaz, 2002) exert strong nutrient demand. The priorities among demands of a tree in the distribution of photosynthate and nutrients, which determines the rate of growth (demand activity) and the size of the demands; in this context, the seed is stronger on the demand of the parts of the fruit, growing shoots and stem tissues storage (Wolstenholme, 1990). The highest concentration of Ca registered seedless fruits of ʻAtaulfoʼ because it is a fruit smaller than the fruit with seed and therefore the Ca concentrated even more.
Conclusions
Applying B into the soil at doses of 50 and 100 g decreased the formation of seedless fruit. Mango trees ʻAtaulfoʼ used in this work, showed N deficiency and it will be necessary to assess whether this influences the formation of seedless fruit or not. The application of 100 g of B only improved the concentration of Ca and B in the plant, and only improved the concentration of B in leaves and floral buds in the end of fruit development. The distribution of nutrients in the normal fruit was as follows: the seed accumulated more N, P, K, Ca and Mg than the pulp with epidermis and both structures had the same concentration of B. The fruits with seeds outperformed those seedless in the concentration of N, P and Mg; while the latter concentrated more Ca, and both types of fruits had the same concentration of K and B.
Literatura citada
Agustí, M. 2004. Fruticultura. Ediciones Mundi-Prensa. Madrid, España. 493 p. [ Links ]
Alcántar, G. G. y Sandoval, V. M. 1999. Manual de análisis químico de tejido vegetal. Sociedad Mexicana de la Ciencia del Suelo. Instituto de Recursos Naturales. Colegio de Postgraduados en Ciencias Agrícolas, Montecillo, Estado de México, México. 156 p. [ Links ]
Beasley, D. R.; Joyce, D. C. and Hofman, P. J. 1999. Effect of preharvest bagging and of embryo abortion on calcium levels in 'Kensington Pride' mango fruit. Aust. J. Exp. Agric. 39: 345-349. [ Links ]
Bellaloui, N.; Reddy, K. N.; Gillen, A. M. and Abel, C. A. 2010. Nitrogen metabolism and seed composition as influenced by foliar boron application in soybean. Plant Soil. 336: 143-155. [ Links ]
Castellanos, J. Z.; Uvalle-Bueno, J. X. y Aguilar-Santelises, A. 2000. Manual de interpretación de análisis de suelos y aguas. Segunda Edición. Instituto de Capacitación para la Productividad Agrícola. Guanajuato, México. 226 p. [ Links ]
Dell, B. and Huang, L. 1997. Physiological response of plants to low boron. Plant Soil. 193: 103-120. [ Links ]
Delgado, A.; Benlloch, M. and Fernández-Escobar, R. 1994. Mobilization of boron in olive trees during flowering and fruit development. HortScience. 29(6):616-618. [ Links ]
Díaz, M. D. H. 2002. Fisiología de árboles frutales. AGT Editor, S. A. México. 390 p. [ Links ]
Epstein, E. and Bloom, A. J. 2005. Mineral nutrition ofplants: principles and perspectives. Second Edition. Sinauer Associates, Inc. Publishers. Sunderland, Massachusetts, USA. 400 p. [ Links ]
Fageria, V. D. 2001. Nutrient interactions in crop plants. J. Plant Nutrit. 24(8): 1269-1290. [ Links ]
Fageria, N. K. 2009. The use of nutrients in crop plants. CRC Press, Boca Raton, Florida. USA. 430 p. [ Links ]
García, E. 1973. Modificación al sistema de clasificación climática de Köppen. Instituto de Geografía. Universidad Nacional Autónoma de México (UNAM). D. F., México. 156 p. [ Links ]
Gupta, U. C. 2007. Boron. Barker, A. V. and Pilbeam, D. J. (Eds). Handbook of plan nutrition. Taylor & Francis Group, Boca Raton, Florida, USA. 241-277 p. [ Links ]
Guzmán, E. C.; Alcalde, B. S.; Mosqueda, V. R. y Martínez, J. A. 1996. Contenido y extracción de algunos nutrimentos por el fruto de mango cv. Manila. Agronomía Tropical. 46(4):431-446. [ Links ]
Lovatt, C. J. and Dugger, W. M. 1984. Boron. Frieden, E. (Ed). Biochemistry of the essential ultratrace elements. Plenum Publisher Corporation. New York, USA. 389-421 p. [ Links ]
Marschner, H. 1995. Mineral nutrition of higher plants. Second Edition. Academic Press. San Diego, California, USA. 889 p. [ Links ]
Medeiros, A. A.; Amorim, J. R. A.; Silva, D. J.; Dantas, J. A. and Guerra, A. G. 2004. Mineral composition of leaves and fruits of irrigated mango trees in Rio Grande do Norte State, Brazil. Acta Horticulturae. 645: 403-408. [ Links ]
Miller, A. J. and Cramer, M. D. 2004. Root nitrogen acquisition and assimilation. Plant Soil 274: 1-36. [ Links ]
Oosthuyse, S.A. 2000. Variation of leaf nutrition status in relation to fruit growth in mango. Acta Horticulturae. 509: 375-378. [ Links ]
Pérez, B. M. H.; Vázquez, V. V. y Osuna, G. J. A. 2007. Caracterización e incidencia del mango niño en huertos comerciales de mango ʻAtaulfoʼ en Nayarit. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Centro. Campo Experimental Santiago Ixcuintla. Santiago Ixcuintla, Nayarit, México. Folleto técnico Num. 355 p. [ Links ]
Rossetto, C. J.; Furlani, P. R.; Bortoletto, N.; Quaggio, J. A. and Igue, T. 2000. Differential response of mango varieties to boron. Acta Horticulturae. 509: 259-264. [ Links ]
Statistical Analysis System (SAS) Institute. 1999. SAS user's guide. Statistics. Version 9. SAS Inst., Cary, NC. USA. Quality, and elemental removal. J. Environ. Qual. 19: 749-756. [ Links ]
SIAP (Servicio de Información Agroalimentaria y Pesquera). 2011. Sistema de Información Estadística Agroalimentaria y Pesquera. Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. http://www.siap.gob.mx/index.php?option=com_content&view=article&id=10&Itemid=15. [ Links ]
Singh, Z. 2005. Embryo abortion in relation to fruit size, quality, and concentrations of nutrients in skin and pulp of mango. J. Plant Nutrit. 28: 1723-1737. [ Links ]
Vázquez-Valdivia, V.; Pérez-Barraza, M. H. y Osuna-García, J. A. 2006. Importancia del cultivo y generalidades. Vázquez-Valdivia, V. y Pérez-Barraza, M. H. (Eds.). El cultivo del mango: principios y tecnología de producción. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Centro, Campo Experimental Santiago Ixcuintla. Santiago Ixcuintla, Nayarit, México. Libro técnico Num. 1. 1-30 pp. [ Links ]
Wolstenholme, B. N. 1990. Resource allocation and vegetative-reproductive competition: opportunities for manipulation in evergreen fruit trees. Acta Horticulturae . 275: 451-459. [ Links ]
Xue-Qun, S.; Bo-Qiang, L.; Guo-Zheng, Q. and Shi-Ping, T. 2011. Antifungal activity and posible mode of action of borate against Colletotrichum gloeosporioides on mango. Plant Dis. 95(1): 63-69. [ Links ]
Zong-min, M.; Ning, Y.; Shu-yun, L. and Hong, H. 2012. Nitrogen requirements for vegetative growth, flowering, seed production, and ramet growth of Paphiopedilum armeniacum (Orchid). HortScience . 47(5): 585-588. [ Links ]
Received: May 2015; Accepted: September 2015
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
