










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 no.5 Texcoco Jun./Ago. 2015
Artículos
Análisis de crecimiento del cultivo de tomate en invernadero*
Growth analysis of tomato in greenhouse
Antonio Juárez-Maldonado1, Karim de Alba Romenus1, Alejandro Zermeño González2, Homero Ramírez1 y Adalberto Benavides Mendoza1§
1 Departamento de Horticultura - Universidad Autónoma Agraria Antonio Narro. Buenavista Saltillo, Coahuila México, C. P. 25315. (juma841025@hotmail.com; kdealba@uaaan.mx; hrr_homero@hotmail.com).
2 Departamento de Riego y Drenaje-Universidad Autónoma Agraria Antonio Narro, Buenavista, Saltillo, Coahuila, México. C. P. 25315. (azermenog@hotmail.com). §Autor para correspondencia: abenmen@gmail.com.
* Recibido: octubre de 2014
Aceptado: marzo de 2015
Resumen
En México el cultivo del tomate es de gran importancia, 70% de los cultivos que se producen bajo condiciones protegidas corresponde al tomate. Por esto es importante realizar un manejo eficiente en la agricultura intensiva para lo que se requieren conocer los factores que condicionan el potencial de producción de los cultivos. El objetivo de éste trabajo fue realizar un análisis de crecimiento de tomate en invernadero. Se desarrollaron dos ciclos de cultivo de tomate durante los años 2011 y 2012. Se eliminaron 4 plantas por semana en las que se determinó el peso fresco y seco de los diferentes órganos. Se realizaron análisis de correlación y de regresión entre los diferentes órganos de la planta de tomate, y una prueba de t-Student entre las variables climáticas de ambos ciclos. Con los datos obtenidos se generaron curvas de crecimiento de los diferentes óiganos de la planta. Se encontró que el cultivo de tomate presentó una etapa de crecimiento exponencial y otra lineal. Se demostró que existe una fuerte correlación entre los pesos frescos y secos de los diferentes órganos de la planta (p≤ 0.01), y en las diferentes etapas de crecimiento (p≤ 0.01). También se observó que la generación y acumulación de biomasa por las plantas de tomate se afectó por las condiciones climáticas internas del invernadero, ya que fueron diferentes en ambos ciclos (p≤ 0.05). La información presentada es útil para diferentes factores del cultivo de tomate en invernadero.
Palabras clave: Lycopersicon esculentum, correlación, curvas de crecimiento, regresión lineal.
Abstract
In Mexico tomato is of great importance, 70% of the crops grown under protected conditions correspond to tomato; therefore it is important to make an efficient management in intensive agriculture, for this is necessary to understand the factors that determine yield potential of the crop. The aim of this work was to perform a growth analysis of tomato under greenhouse. Two crop cycles of tomato were evaluated during 2011 and 2012. 4 plants per week where removed, to which were determined fresh and dry weight of the different organs. Correlation and regression analysis between the different organs of tomato plant and T Student test between climatic variables of both cycles were performed. With the data obtained, growth curves of different plant organs were generated. It was found that tomato showed an exponential and a linear growth stage. It was shown that there is a strong correlation between fresh and dry weight of different plant organs (p≤ 0.01), and at different growth stages (p≤ 0.01). It was also observed that the generation and accumulation of biomass by tomato plants were affected by the climatic conditions in the greenhouse, since they were different in both cycles (p≤ 0.05). The information in this paper is useful for different factors in tomato under greenhouse conditions.
Keywords: Lycopersicon esculentum, correlation, growth curves, linear regression.
Introducción
La producción de cultivos en invernaderos es de suma importancia ya que nos da una ventaja sobre la producción a cielo abierto porque se establece una barrera entre el ambiente externo y el cultivo, creando un microclima interno que permite proteger el cultivo de condiciones adversas (viento, granizo, plagas, etc.) y controlar factores como la temperatura, radiación, concentración de CO2, humedad relativa, etc. En México, el uso de invernaderos para la producción de hortalizas ha aumentado rápidamente, de 721 ha en 1999 a 3 200 ha en 2005 (Ocaña-Romo, 2008), la cual en 2009 se extendió a una superficie de 10 000 ha (Perea, 2009).
Los datos más recientes muestran que en 2012 se llegó a 12 000 ha de invernaderos, esto sin incluir otras 8 000 has que corresponden a malla sombra y macrotúnel (SAGARPA, 2012). En México el cultivo del tomate es sumamente importante, ya que de los principales cultivos que se producen en condiciones protegidas este ocupa 70%, seguido por pimiento (16%) y pepino (10%) (SAGARPA, 2012).Aunado a esto, México es el principal exportador a nivel internacional, enviando el producto a Estados Unidos de América, Canadá y El Salvador, tan sólo en 2011 se produjeron 1 872 000 toneladas (MÉXICOPRODUCE, 2012).
Considerando el factor de importancia que tiene el cultivo del tomate es importante realizar un manejo eficiente en la agricultura intensiva por lo que se requieren conocer los factores que condicionan el potencial de producción de los cultivos.
En este sentido, la correcta aplicación de riego es uno de los principales factores que afecta el rendimiento del cultivo (Flores et al, 2007), entendiendo que el rendimiento está determinado por la capacidad de acumular materia seca en los órganos destinados a la cosecha (Casierra-Posada et al., 2007). Además, aunque existen técnicas exitosas como el fertirriego, aún hay problemas con la dosificación de fertilizantes que deben aplicarse, por lo que se ha propuesto usar la acumulación de materia seca para cuantificar la demanda nutrimental (Bugarín-Montoya et al, 2002). Por esta razón, prácticas como la fertilización y el riego deben ser definidas en función de las características propias de crecimiento del cultivo de interés (Fynn et al., 1989; Enriquez-Reyes et al., 2003).
Aunado a lo anterior, los avances tecnológicos brindan técnicas novedosas como la simulación de cultivos bajo invernadero, principalmente en el tomate (Heuvelink y Marcelis, 1989; Marcelis et al, 1989; Heuvelink, 1995; Heuvelink y Buiskool, 1995; Heuvelink, 1996; Heuvelink, 1999; Marcelis y Heuvelink, 1999; de Gelder et al, 2005; Marcelis et al., 2009). La simulación de cultivos trata de imitar el crecimiento de los cultivos mediante ecuaciones matemáticas; para estos modelos de simulación es de vital importancia conocer a fondo el crecimiento y distribución de materia seca entre los diferentes órganos de la planta.
En este sentido y debido a la importancia económica del cultivo de tomate en México, el objetivo del presente estudio fue realizar un análisis de crecimiento bajo condiciones de invernadero, el cual nos permita entender de mejor manera el comportamiento de este cultivo para determinar un manejo eficiente de los recursos tanto de agua como fertilizantes, así como proveer de información para la generación de modelos de crecimiento.
Materiales y métodos
Desarrollo del cultivo de tomate
Se estableció un experimento con plantas de tomate (Lycopersicon esculentum Mill.) con el fin de conocer la dinámica de crecimiento del cultivo bajo condiciones de invernadero en la región norte de México. Para ello se realizaron dos repeticiones del cultivo durante los años 2011 y 2012, del 3 julio al 30 de octubre y del 6 de mayo al 23 de septiembre respectivamente. La semilla utilizada fue el híbrido "Caimán" de hábito de crecimiento indeterminado y de tipo bola. El invernadero en el que se estableció el experimento es de tipo capilla con cubierta de policarbonato, además de contar con un sistema automático para el control de las temperaturas en su interior.
Las plantas fueron establecidas en macetas plásticas de 19 L y con una densidad de 3 plantas m-2; se usó un sistema de cultivo sin suelo utilizando como sustrato una mezcla de perlita y peat moss en proporción 1:1. Se utilizó un sistema de riego dirigido con microtubín y goteros tipo estaca de alto flujo para cada maceta. Además se instalaron temporizadores automáticos para realizar cuatro riegos por día a diferentes horarios (8:00, 12:00, 16:00 y 20:00 h). La cantidad de riego aplicada fue diferente para cada etapa fenológica aplicando alrededor de 2.4 L por planta por día en las etapas de mayor consumo. La nutrición del cultivo se dio con la aplicación de solución Steiner (Steiner, 1961) y manejando diferentes concentraciones de acuerdo a la etapa fenológica. El cultivo se trabajó a un tallo, y buscando que los racimos en la medida de lo posible fueran de cinco frutos, por lo que en la mayor parte del tiempo se realizaron aclareos de fruto. Las plantas se limitaron en su crecimiento eliminando la parte apical a las 13 semanas después del trasplante (SDT) alcanzando un promedio de 10 racimos por planta y una altura aproximada de 3.5 m.
Para determinar el crecimiento del cultivo se realizaron muestreos destructivos semanales de cuatro plantas de tomate; éstas se separaron en hojas, tallo y frutos y se obtuvieron sus pesos frescos. Después de secar en horno de secado a 80 °C durante 4 días se obtuvo el peso seco de las diferentes partes de la planta. Asimismo, se cuantificó el total de poda realizada a cada planta y el total de frutos cosechados, de los que se obtuvo también peso fresco y seco. Con estos datos se determinó a su vez el peso de la parte aérea que considera la suma de hojas, tallo y frutos tanto del peso fresco como del peso seco. También se obtuvo la biomasa acumulada total que considera la suma de la parte aérea más la hoja podada y el fruto cosechado, tanto de peso fresco como de peso seco. La suma de fruto en la planta más el fruto cosechado fue otra variable que se obtuvo para peso el fresco y seco.
Análisis estadísticos
El diseño del experimento fue completamente al azar y la obtención de las muestras fue de manera aleatoria. Para determinar el grado de correlación existente entre los diferentes órganos de la planta evaluados, se obtuvo el coeficiente de correlación de Pearson. Para dicho proceso se utilizó el complemento EZAnalyze© para Excel. Con esto se obtuvieron matrices de correlación entre las diferentes variables evaluadas, así como la correlación existente entre el peso fresco y seco de los diferentes órganos de la planta. Finalmente, para determinar las tendencias de crecimiento a lo largo de ambos ciclos de cultivo, se realizaron análisis de regresión simple y para verificar el ajuste de los mismos se usó el coeficiente de determinación (R2), este proceso se realizó en el complemento Regress© para Excel.
Resultados y discusión
Después de concluir los procesos experimentales se generaron curvas de crecimiento de los diferentes órganos de las plantas de tomate. En las Figuras 1 y 2 se presentan las curvas correspondientes al ciclo 2011 para los pesos frescos y secos respectivamente, mientras que las curvas correspondientes al ciclo 2012 se muestran en las Figuras 3 y 4. Dichas curvas muestran el comportamiento del cultivo de tomate con respecto al tiempo por lo que se pueden utilizar en la planeación de las labores culturales (Casierra-Posada, 2007). En las Figuras 1 y 2 puede observarse que las tendencias de crecimiento tanto en peso fresco como seco son muy similares, corroborándose lo anterior en el ciclo 2012 (Figuras 3 y 4).


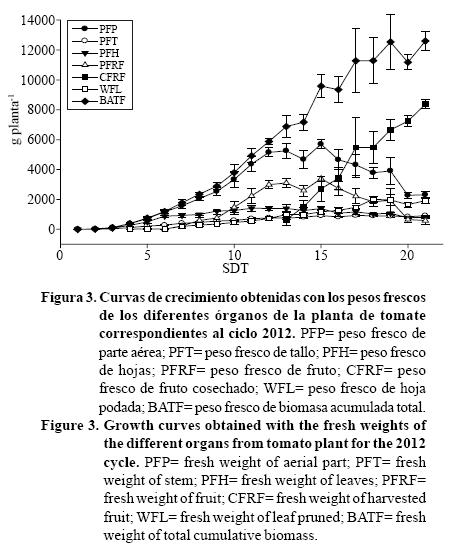

Por otra parte, en el Cuadro 1 se presentan las correlaciones obtenidas entre los pesos frescos y secos de los diferentes órganos de la planta, además se muestran las ecuaciones de regresión generadas y su correspondiente R2. La similitud mostrada entre las curvas de crecimiento en peso fresco y seco, se debe a que el peso fresco está directamente correlacionado con el peso seco (Heuvelink y Marcelis, 1989), demostrándose esto con los coeficientes de correlación obtenidos entre los diferentes órganos de la planta, ya que todos fueron mayores a 0.97 (Pearson, p≤ 0.01) (Cuadro 1). Considerando estos resultados, es posible entonces estimar el peso seco a partir del peso fresco de cualquier parte de la planta de tomate con gran exactitud utilizando las ecuaciones de regresión generadas, ya que el R2 para casi todos los casos fue mayor que 0.94, con excepción del correspondiente a hoja podada en 2011 (R2= 0.8056) pero en este caso puede emplearse la ecuación correspondiente al ciclo 2012 que fue significativamente mayor (R2= 0.9813) (Cuadro 1).
Tanto en el ciclo 2011 como en el 2012 se aprecian algunas tendencias de crecimiento presentadas en etapas específicas. En el Cuadro 2, se muestran las ecuaciones de regresión obtenidas mediante análisis de regresión junto con su correspondiente R2, además, se incluye el periodo de tiempo del que se tomaron los datos para realizar dicho análisis. En ambos ciclos se presenta un crecimiento exponencial en el tallo, hojas, parte aérea y BAT correspondiente a las primeras 4 SDT, esto sin importar si es peso fresco o seco. Esto se constata con las ecuaciones exponenciales obtenidas y sus correspondientes R2 que para todos los casos fue mayor que 0.97 (Cuadro 2).
Asimismo, se observó que entre las semanas 5-13 el crecimiento cambia su comportamiento y se vuelve lineal para todos los órganos de la planta incluso en el fruto, parte aérea y BAT (Cuadro 2). Este comportamiento concuerda con el reportado por Enriquez-Reyes et al. (2003), al mencionar que el tomate de crecimiento indeterminado presenta dos etapas bien definidas, a las que denominan una de crecimiento juvenil y la otra de reproducción. También considerando sólo la BAT, la tendencia es similar a los resultados presentados en otros estudios (Heuvelink, 1995; Heuvelink y Buiskool, 1995; Heuvelink, 1999; Xiao et al, 2004; de Gelder et al., 2005; Marcelis et al., 2009). Aunque las ecuaciones de regresión obtenidas mostraron valores del R2 más bajos que en el crecimiento exponencial, sólo el de las hojas del ciclo 2012 mostró un R2= 0.7248 mientras que el resto de los órganos presentó valores superiores a 0.80 (Cuadro 2). Este hecho puede explicarse debido a que constantemente se realizó poda de hojas lo que generó variabilidad en los datos de esta variable en particular, tal como lo indica Heuvelink (1999).
Debido a que en el ciclo 2011 el cultivo del tomate se trabajó sólo por 18 SDT, solamente en el ciclo 2012 fue posible determinar una tendencia clara posterior a la semana 14 después del trasplante, ya que en este ciclo se trabajó 21 SDT. En dicho ciclo, la parte aérea, hojas y frutos mostraron una tendencia lineal decreciente entre las semanas 15-21, aunque los R2 estuvieron entre 0.58 y 0.78 (Cuadro 2) esto fue debido posiblemente a la gran variabilidad de los datos. En las Figuras 3 y 4 se puede apreciar el comportamiento decreciente antes mencionado, éste se debió a que la planta se limitó en su crecimiento en la semana 13 después del trasplante, y a que la poda de hojas y cosecha de frutos se mantuvieron hasta el final del cultivo.
A diferencia de lo anterior, la BAT se mantuvo con una tendencia de crecimiento lineal desde la 5ta SDT hasta el final del cultivo para ambos ciclos, esto se confirma ya que el R2 para ambos casos fue mayor que 0.95 (Cuadro 2). Esta tendencia es importante ya que a diferencia de otros estudios en los que se presenta solo el crecimiento lineal de tomate (Heuvelink, 1995; Heuvelink y Buiskool, 1995; Heuvelink, 1999; Xiao et al, 2004; de Gelder et al, 2005; Marcelis et al, 2009), el conocer el comportamiento de este cultivo después de que se limita su crecimiento es útil para determinar de mejor manera el manejo tanto cultural como la aplicación de riego y nutrientes en esa etapa específica. Además, es posible estimar de manera más adecuada el tiempo de cosecha esperado cuando se trabaja el cultivo de tomate en ciclos cortos con el objetivo de concentrar la producción en periodos de demanda específicos tal y como lo resaltan Flores et al. (2007).
En los Cuadros 3 y 4, se presentan las matrices de correlación obtenidas para las diferentes variables evaluadas. En éstas, se detalla el grado de correlación, el ciclo de cultivo al que corresponde (2011 ó 2012) y adicionalmente se muestra el número de SDT (13 ó 18) empleado para realizar dicho análisis. En el Cuadro 5 se muestra la correlación obtenida 21 SDT y correspondiente solamente a los datos del ciclo 2012 debido a las fechas en que se trabajó dicho cultivo como ya se explicó anteriormente. En el Cuadro 3, se puede observar que sólo la variable fruto fresco cosechado no presenta alguna correlación clara con el resto, todas las demás presentan correlación altamente significativa (Pearson, p≤ 0.01). Esto se puede atribuir a las fechas en que se inicia la cosecha ya que en 2011 comenzó en la 14va SDT mientras que en 2012 inició en la 13va SDT y las correlaciones mostradas abarcan sólo hasta la semana 18.
Cuando se consideró mayor tiempo de evaluación lo anterior cambió, ya que a las 21 SDT el fruto fresco cosechado mostró correlaciones altamente significativas (Pearson, p≤ 0.01), siendo positivas con respecto al peso fresco de tallo y negativas con respecto a la peso fresco de la parte aérea, peso fresco de hojas y al peso fresco del fruto en la planta (Cuadro 5). En el Cuadro 4 correspondiente a los pesos secos se observa prácticamente lo mismo que en el Cuadro 3, lo cual se esperaba debido a la alta correlación existente entre el peso fresco y seco (Cuadro 1) (Heuvelink y Marcelis, 1989).
Considerando lo mostrado en los Cuadros 3, 4 y 5, se puede afirmar que existe una distribución estable de la biomasa total generada entre los diferentes órganos de la planta en la etapa de crecimiento exponencial y en la de crecimiento lineal la cual puede variar principalmente por efecto de la temperatura (Heuvelink y Marcelis, 1989; Heuvelink, 1999; Enriquez-Reyes et al, 2003; Xiao et al, 2004). Aunque es importante recalcar que la proporción de biomasa distribuida cambia de acuerdo con la etapa fenológica (Marcelis et al, 1989), lo cual se entiende ya que la aparición de frutos se da hasta después de la 4ta SDT (Figuras 1-4)(2,3). Dicha distribución se puede mantener siempre y cuando se mantenga en crecimiento la planta, ya que al momento en que se detiene el crecimiento cambia el comportamiento (Cuadro 2, Figuras 1-4)(2,3) y por consecuencia la distribución de biomasa.
Conclusiones
En éste trabajo se presentó un análisis de crecimiento de tomate en invernadero en el que se generaron las curvas de crecimiento de los diferentes órganos de la planta. El cultivo de tomate presentó dos etapas de crecimiento bien definidas, una exponencial en las primeras cuatro semanas después del trasplante y la otra lineal a partir de la cuarta semana. Adicionalmente, cuando se detiene el crecimiento del cultivo se presenta una etapa más con tendencia decreciente en frutos y hojas.
Se demostró que existe una fuerte correlación entre los pesos frescos y secos de los diferentes órganos de la planta. Además se encontró también correlación entre los diferentes órganos de la planta en las etapas de crecimiento exponencial y lineal.
La información presentada aquí puede ser útil en la planeación de los requerimientos de agua y nutrientes a lo largo de un ciclo de cultivo. También se puede utilizar para definir épocas de producción para aprovechar los mejores precios del mercado. Además, se puede aplicar en áreas poco exploradas como la modelación y simulación de tomate cultivado en invernadero.
Agradecimientos
Los autores agradecen al Departamento Forestal de la UAAAN, por facilitar el espacio necesario para la realización de este trabajo.
Literatura citada
Bugarín-Montoya, R.; Galvis-Spinola, A.; Sánchez-García, P. and García-Paredes, D. 2002. Daily accumulation of aboveground dry matter and potassium in tomato. Terra Latinoamericana. 20(4):401-409. [ Links ]
Casierra-Posada, F.; Cardozo, M. C. and Cárdenas-Hernámdez, J. F. 2007. Growth analysis of tomato fruits (Lycopersicon esculentum Mill.) cultivated in greenhouse. Agron. Colombiana. 25(2):299-305. [ Links ]
De Gelder, A.; Heuvelink, E. and Opdam, J. J. G. 2005. Tomato yield in a closed greenhouse and comparison with simulated yields in closed and conventional greenhouses. Acta Hortic. 691:549-552. [ Links ]
Enriquez-Reyes, S. A.; Alcántar-González, G.; Castellanos- Ramos, J. Z.; Suárez, E.A.; González-Eguiarte D. and Lazcano-Ferrat, I. 2003. NUMAC-N tomato: mineral nutrition fit at growth. The nitrogen nutrition in tomato greenhouse production. 1. Model description and parameters adjust. Terra Latinoamericana. 21(2):167-175. [ Links ]
Flores, J.; Ojeda-Bustamante, W.; López. I.; Rojano, A. and Salazar, I. 2007. Water requirements for greenhouse tomato. Terra latinoamericana. 25(2):127-134. [ Links ]
Fynn, R. P.; Roller, W. L. and Keener, H. M. 1989. A decision model for nutrition management in controlled environment agriculture. Agric. Systems. 31:35-53. [ Links ]
Heuvelink, E. 1995. Dry matter production in a tomato crop: measurments and simulation. Ann. Bot. 75:369-379. [ Links ]
Heuvelink, E. 1996. Dry matter partitioning in tomato: validation of a dynamic simulation model. Ann. Bot. 77:71-80. [ Links ]
Heuvelink, E. 1999. Evaluation ofa dynamic simulation model for tomato crop growth and development. Ann. Bot. 83:413-422. [ Links ]
Heuvelink, E. and Marcelis, L. F. M. 1989. Dry matter distribution in tomato and cucumber. Acta Hortic. 260:149-157. [ Links ]
Heuvelink, E. and Buiskool, R. P. M. 1995. Influence of sink-source interaction on dry matter production in tomato. Ann. Bot. 75:381-389. [ Links ]
Marcelis, L. F. M. and E. Heuvelink, E. 1999. Modelling fruit growth and dry matter partitioning. Acta Hortic. 499:39-49. [ Links ]
Marcelis, L. F. M.; Heuvelink, E. and de Koning, A. N. M. 1989. Dynamic simulation ofdry matter distribution in greenhouse crops. Acta Hortic. 248:269-276. [ Links ]
Marcelis, L. F. M.; Elings, A.; de Visser, P. H. B. and Heuvelink, E. 2009. Simulating growth and development of tomato crop. Acta Hortic. 821:101-110. [ Links ]
MÉXICOPRODUCE. 2012. Productos: jitomate. http://www.mexicoproduce.mx/productos.html#jitomate. [ Links ]
Ocaña-Romo, C. R. 2008. En crecimiento. Desarrollo de invernaderos en México. http://www.hortalizas.com. [ Links ]
Perea, 2009. Invernaderos y riego. Aun con crisis seguirá creciendo superficie de invernadero. La imagen agropecuaria. http://www.imagenagropecuaria.com. [ Links ]
SAGARPA (Secretaría de Agricultura, Ganadería, Pesca y Alimentación). 2012. Agricultura protegida 2012. http://www.sagarpa.gob.mx/agricultura/paginas/agricultura-protegida2012.aspx. [ Links ]
Steiner, A. A. 1961. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil. 15:134-154. [ Links ]
Xiao, S.; van der Ploeg, A.; Bakker, M. and Heuvelink, E. 2004. Two instead of three leaves between tomato trusses: Measured and simulated effects on partitioning and yield. Acta Hortic. 654:303-308. [ Links ]

