










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 no.1 Texcoco Jan./Fev. 2015
Artículos
Viabilidad de polen, receptividad del estigma y tipo de polinización en cinco especies Echeveria en condiciones de invernadero*
Pollen viability, stigma receptivity and pollination type in five Echeveria species under greenhouse conditions
Teresa J. Rodríguez-Rojas1, María Andrade-Rodríguez1§, Jaime Canul-Ku2, Antonio Castillo-Gutiérrez1, Edgar Martínez-Fernández1 y Dagoberto Guillén-Sánchez1
¹ Facultad de Ciencias Agropecuarias-Universidad Autónoma del Estado de Morelos. Avenida Universidad 1001. 62209. Colonia Chamilpa, Cuernavaca, Morelos, México. Tel: 01 777 3 70 29 46. (tererodrojas@hotmail.com; mariamaria.andrade65@gmail.com; acastillo-g@hotmail.com; edgar@uaem.mx; dagoguillen@yahoo.com).
² Instituto Nacional de Invetigaciones Forestales Agrícolas y Pecuarias. Carretera Zacatepec-Galeana, km 0.5. 62780. Colonia Centro, Zacatepec, Morelos, México. (canul.jaime@inifap.gob.mx). §Autora para correspondencia: andradem65@hotmail.com.
* Recibido: septiembre de 2014
Aceptado: enero de 2015
Resumen
Echeveria, género representativo de Crassulaceae por representar 97% de especies endémicas; sus plantas tienen características morfológicas atractivas para la horticultura ornamental. El conocimiento de las características reproductivas es útil para el mejoramiento genético, necesario para ser utilizado en las polinizaciones y aumentar la posibilidad de éxito de la fecundación. El objetivo fue conocer la viabilidad de polen, receptividad del estigma y tipo de polinización de cinco especies de Echeveria, con fines de mejoramiento genético. El trabajo se realizó de 2011 a 2013, en Cuernavaca, Morelos. Se usaron cinco especies E. agavoides, E. elegans, E. runyonii, E. pumila, E. perle. La viabilidad de polen se determinó mediante el método de tinción con ácido ácetico-carmín. La receptividad del estigma se evaluó con el método de Osborn; la evaluación fue a las 8:00, 10:00, 12:00, 14:00, 16:00, y 18:00 h. Para tipo de polinización se realizaron cuatro modalidades: 1) autopolinización; 2) emascular y cubrir flores; 3) polinización cruzada intra específica; y 4) polinización cruzada inter específica; se evaluo amarre de fruto y viabilidad de semillas. E. agavoides tuvo mayor porcentaje de polen viable (72.7%). La mayor receptividad se tuvo de 12:00 a 14:00 h (94.6 a 98%). En las cuatro modalidades de polinización se obtuvó 100% de amarre de fruto; solo hubo semilla viable en los frutos de polinización cruzada intra-específica (12.3%) y polinización cruzada inter-específica (11.3 a 12.3%). La polinización de las especies estudiadas fue cruzada. E. perle y E. runyonii no pueden fungir como hembras por no formar semillas viables.
Palabras clave: Echeveria spp., polinización cruzada, reproducción generativa, viabilidad de semillas.
Abstract
Echeveria, is a representative genus from Crassulaceae to represent 97% of endemic genus; its plants have attractive morphological characteristics for ornamental horticulture. Knowledge of reproductive characteristics are useful for genetic improvement, necessary to be used in pollination and increase the chance of successful fertilization. The objective was to determine pollen viability, stigma receptivity and pollination type of five species of Echeveria, for breeding purposes. The work was done from 2011 to 2013, in Cuernavaca, Morelos. Five species E. agavoides, E. elegans, E. runyonii, E. pumila, E. perle were used. Pollen viability was determined by staining with carmine acetic acid. Stigma receptivity was assessed with the Osborn method; assessment was at 8:00, 10:00, 12:00, 14:00, 16:00, and 18:00 h. For pollination type four modes were performed: 1) self-pollination; 2) emasculating and cover flowers; 3) intra-specific cross-pollination; and 4) inter-specific cross-pollination; fruit set and seed viability was evaluated. E. agavoides had higher percentage of viable pollen (72.7%). Greater receptivity was from 12:00 to 14:00 hrs (94.6 to 98%). In the four modes of pollination 100% of fruit set was obtained; there was only viable seeds in fruits of intra-specific cross pollination (12.3%) and inter-specific cross pollination (11.3 to 12.3%). The pollination of the species studied were crossed E. perle and E. runyonii can not serve as females by not forming viable seeds.
Keywords: Echeveria spp., cross-pollination, generative reproduction, seed viability.
Introducción
La familia Crassulaceae es la más representativa de México, debido al alto endemismo de la mayoría de sus especies, su significado histórico-cultural, y por el papel que desempeñan en la estructura y función de los ecosistemas áridos del país (Toledo, 1988). Los géneros de mayor importancia son Sedum, Echeveria, Villadia y Dudleya, porque tienen mayor número de especies, y varias especies son endémicas de México. Echeveria cuenta con 127 especies, de las cuales 97.1% son endémicas (Thiede, 1995).
La Echeveria son plantas perenes, suculentas, glabras a hirsutas, fibrosa o tuberosas, compactas o difusas, con hojas en forma de roseta, submembranosas a muy suculentas, usualmente lanceoladas o oblanceoladas, inflorescencias con numerosas flores, las cuales varían de cilíndricas a pentagonales, con 5 pétalos de varios colores que van desde el blanco, amarillo, anaranjado, y rojo (Eggli, 2003).
Conocer aspectos de la biología reproductiva es el primer paso para el mejoramiento genético clásico, basado en la selección de individuos superiores y cruzamientos dirigidos. Poco se conoce de la biología reproductiva de estas especies; estos conocimientos son indispensables para ser utilizados en las polinizaciones, y poder aumentar la posibilidad de éxito de la fecundación. Algunos de estos aspectos son la viabilidad del polen (Nyffeler, 1992), la receptividad del estigma (Shivanna y Sawhney, 1997), y el tipo de polinización, ya que de estos depende una reproducción exitosa (Márquez et al., 2013).
La viabilidad del polen es una medida de la fertilidad masculina (Kearns e Inouye, 1993), el polen viable es fundamental para el proceso de reproducción, por tanto, la evaluación de la fertilidad del polen es esencial (Nyffeler, 1992); en general la longevidad del polen puede ser afectada por la temperatura y la humedad relativa (Shivanna et al., 1997); en muchos híbridos el polen puede ser encogido o contraído y no viable (Kearns e Inouye, 1993).
La receptividad del estigma es la capacidad para recibir el polen, permitir que se adhiera, se hidrate y germine (Shivanna et al., 1997). Esto puede determinarse mediante la evaluación de la presencia de enzimas peroxidasas (Kearns e Inouye, 1993).
La polinización consiste en la transferencia de polen desde los órganos sexuales masculinos a los órganos sexuales femeninos; a pesar de su aparente simplicidad, la incapacidad de las plantas para mover sus gametos por sí mismas ha propiciado su evolución mediante diferentes adaptaciones; es decir, diferentes tipos de polinización (Márquez et al., 2013), como polinización autógama, y polinización alógama.
Para que el proceso de la polinización ocurra, la transferencia del polen al estigma debe suceder durante el periodo en que el estigma se encuentre receptivo, en caso contrario, el polen no puede adherirse y no puede germinar (Kearns e Inouye, 1993).
Para que la polinización autógama ocurra debe haber compatibilidad del polen, esto garantiza que se fertilice cada óvulo; la autoincompatibilidad evita la producción de semillas por autopolinización (autógama), esto se debe a que el polen que se adhiere al estigma no germina, y es incapaz de penetrar o crecer por el estilo (Kearns e Inouye, 1993).
La emasculación de todas las flores en plantas multiflorales puede demostrar si la polinización alógama ocurre, ya sea intra o inter específica, si se produce semilla debe ser de polinización cruzada a menos que ocurra apomixis (Kearns e Inouye, 1993); es decir, que se produzcan semillas independientemente de la fecundación (Márquez et al., 2013).
Charlesworth y Charlesworth (1987); Cruden (1976) sugieren que la autopolinización es muy común en las especies de la familia Crassulaceae, y que presentan autogamia facultativa. En contraste, Carque et al. (1996) observaron que las Crassulaceae tienen polinización anemófila, hidrófila (produciendo autopolinización), y entomófila (produciendo polinización cruzada). Izco et al. (1998) observó que en Kalanchoe la polinización es entomófila u ornitófila.
En muchos estudios la producción de semillas es suficiente como indicador de una polinización exitosa, pero debido a la gran importancia de la semilla es necesario confirmar su viabilidad, para esto la prueba definitiva es la germinación (Kearns e Inouye, 1993), ya que las semillas contienen los genes recombinados y de ellas depende la supervivencia del germoplasma (Espinosa-Osornio y Engleman, 1998).
Por lo anterior, el objetivo de la presente investigación fue conocer algunos aspectos de la biología reproductiva de cinco especies del género Echeveria, determinando la viabilidad de polen, el momento de receptividad del estigma y tipo de polinización, con fines de mejoramiento genético.
Materiales y métodos
La investigación se desarrolló de agosto de 2011 a septiembre de 2013, en un laboratorio de propagación vegetativa y en un invernadero cubierto con plástico lechoso uv 2, con ubicación geográfica de 18° 58’ 54.71’’ latitud norte y 99° 13’ 59.14’’ longitud oeste, y 1 876 msnm, en Cuernavaca, Morelos. El clima es semicalido y subhúmedo, precipitación anual de 1 061 mm, temperatura media anual de 20 °C, (A)Ca(w1)(w)(i´)gw´´ (García, 1981).
El material genético estudiado fueron cinco especies del género Echeveria cultivadas en maceta: E. agavoides Lem., E. elegans Rose, E. runyonii topsy-turvy, E. pumila var. glauca x E. walther, E. perle von Nuremberg; obtenidas en viveros comerciales, éstas fueron seleccionadas por su porte, el color y forma de las hojas, la capacidad de producir hijuelos, ser algunas de las más comercializadas, así como por la coincidencia en época de floración (Figura 1).

Viabilidad de polen
Para estudiar la viabilidad de polen se usaron tres plantas de cada especie, se colectaron tres flores por planta, y tres anteras por flor, antes de que ocurriera la antesis, éstas se colocaron en un tubo eppendorf de 1.5 mL, y a cada tubo se le agregó una solución Farmer (alcohol etílico absoluto, ácido acético glacial, 3:1), y se conservaron en refrigeración a 4 ºC hasta su evaluación.
La viabilidad se evaluó mediante el método de tinción con ácido ácetico-carmín; cada antera se colocó sobre un portaobjetos, se cortó transversalmente con un bisturí, se agregó una gota de ácido acético-carmín y se aplastó con cuidado para que se liberará el polen en la gota de colorante; el portaobjetos se pasó ligeramente por la flama de un mechero, después se observaron los granos de polen al microscopio (40 x). Se realizó una preparación por cada antera, en cada preparación se observaron cuatro campos, los granos de polen fértiles se tiñeron de color rojo y los no viables sin coloración, y la variable fue el porcentaje de polen viable, además se obsevó la ornamentación de los granos de polen y se midieron con una regla para microscopio cuyas unidades de medición fueron las micras.
Se utilizó un diseño experimental completamente al azar, se tuvieron cinco tratamientos (especies) con tres repeticiones (plantas) y se evaluaron nueve anteras por repetición.
Receptividad del estigma
Se estudio la receptividad del estigma durante el transcurso del día (8:00, 10:00,12:00, 14:00, 16:00, y 18:00 h), cuando la flor estaba en apertura floral; se empleó el método de Osborn et al. (1988), que se basa en la reacción de la enzima peroxidasa al colocar una gota de peróxido de hidrógeno al 40% sobre los estigmas de las flores, la producción de burbujas indica que el estigma esta receptivo.
Este experimento se realizó en un diseño completamente al azar con arreglo factorial de tratamientos 5 * 6 (5 especies - 6 horarios); se usaron cinco repeticiones (plantas), en cada repetición se evaluaron tres flores por planta, así se estudiaron 15 flores por especie en cada horario. La variable de respuesta fue la receptividad o no receptividad del estigma.
Tipo de polinización
Se realizaron cuatro modalidades de polinización para saber si las plantas de las cinco especies son autógamas, apomícticas, alógamas intra-específicas o alógamas inter-específicas. Las flores seleccionadas se cubrieron con agribon® después de aplicar el tratamiento de polinización correspondiente, se etiquetaron y se cuidaron hasta cosechar los frutos. Los tratamientos fueron: 1) polinización autógama, las flores en botón se cubrieron para evitar la llegada de polen externo y así se mantuvieron hasta el desarrollo de fruto; 2) apomixis, se emascularon las flores antes de la antesis y se cubrieron para comprobar si desarrollan fruto; 3) polinización cruzada intra-específica, las flores se emascularon antes de la antesis, se cubrieron y posteriormente se llevó polen de otra planta de la misma especie colocándolo en el estigma de las flores, éstas se cubrieron y se mantuvieron así hasta observar fruto; y 4) polinización cruzada inter-específica, las flores se emascularon antes de la antesis y se cubrieron, se aplicó polen de una planta de otra especie colocándolo en el estigma de las flores, éstas se cubrieron nuevamente hasta que ocurrió desarrollo de fruto.
El experimento fue desarrollado en un diseño completamente al azar con arreglo factorial de tratamientos 5 * 4 (5 especies - 4 modalidades de polinización), en cada tratamiento se tuvieron cinco repeticiones (plantas) por especie, con tres flores por planta. Se evaluó porcentaje de amarre de fruto y viabilidad de semillas.
Porcentaje de amarre de fruto. Se cosecharon los frutos desarrollados en cada una de las cuatro modalidades de polinización y se observó la presencia de semillas. Dado que se obtuvó 100% de amarre de fruto y no hubo varianza en cuanto a tratamientos, no se realizó el análisis estadístico de los datos.
Viabilidad de semillas. La presencia de semillas no es garantía de viabilidad, razón por la cual las semillas desarrolladas en cada uno de los cuatro tratamientos de polinización se pusieron a germinar in vitro, dado que son muy pequeñas (0.3 a 0.6 mm) para evaluar su viabilidad. Se usó el medio de cultivo WPM modificado por Parada y Villegas (2009), adicionando 5 ppm de AG3. Se contó el número de plántulas por frasco (repetición) y se obtuvó el porcentaje de germinación.
Los datos de los tres experimentos fueron estudiados mediante análisis de varianza y prueba de comparación de medias (Tukey, p≤ 0.05).
Resultados y discusión
Viabilidad de polen
Los granos de polen de las cinco especies midieron de 30 a 40 µm de diámetro, el polen maduro es esferoidal y tricolporado (cuenta con tres polos), se observó la misma forma de polen en las cinco especies estudiadas (Figura 2). Lo anterior fue similar a lo encontado por Arreguín-Sánchez et al. (1990) quienes estudiaron la morfología del polen de los géneros Echeveria, Sedum y Villadia, y observaron que en los tres géneros los granos de polen son prolato-esferoidal y tricolporados; indican que en Echeveria el tamaño varió de 27 a 42 µm y concluyen que los granos de polen de la familia Crassulaceae son homogéneos en su forma, tamaño y ornamentación. De igual forma, otros autores observaron la ornamentación de los granos de polen de las Crassulaceae y los describen como tricolporados (Huang, 1972). Sin embargo, Karaer et al. (2010) reportan que las características de polen de tres especies Crassulaceae, Sempervivum sosnowskyi, S. armenum y S. glabrifolium fueron similares entre ellas, la forma fue sub oblonga a ovalada esferoidal y su tamaño varió de 3 a 23.2 µm, mencionan que la forma del polen puede ser significativo en la distinción de especies. También Parnell (1991) evaluó el polen de 32 taxones del género Sempervivum y Jovibarba (Crassulaceae) y mencionan que ambos géneros tienen pequeños granos de polen (diámetro <30 µm), pero con diferente ornamentación.
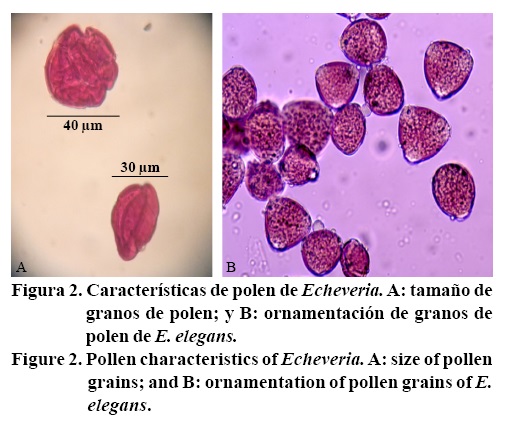
Los resultados en viabilidad de polen mostraron que existen diferencias estadísticas (p≤ 0.1) entre especies, E. agavoides presentó mayor porcentaje de viabilidad de polen, en comparación con E. runyonii y E. perle que presentaron la menor cantidad de gametos masculinos fértiles (Figura 3). El bajo porcentaje de viabilidad en E. runyonii (59%) y E. perle (59.27%) puede deberse a su origen genético, E. runyonii proviene de una mutación con rayos gama de Echeveria Chalk Rose 'Lucita', realizada en California, E.U; E. perle es un híbrido inter-específico de Echeveria gibbif lora var. metalica por E. potosina, creada en Alemania (Sajeva y Constanzo, 1997; Anónimo, 2013), y de acuerdo con Uhl (1982), los granos de polen de los mutantes e híbridos interespecíficos son altamente estériles. La baja viabilidad de polen puede afectar la cantidad de semillas viables en los cruzamientos donde se usen como progenitores masculinos éstas dos especies de Echeveria.

Receptividad del estigma
El análisis de varianza indicó que no hubo diferencias en receptividad del estigma por efecto de la especie, ya que ésta fue similar en las cinco especies estudiadas. Sin embargo, ésta caracteristica si varió en función del horario en que se hizo la evaluación, el cual tuvo efecto altamente significativo (p≤ 0.01); la interacción entre las especies y los horarios de evaluación no fue significativa (Cuadro 1).
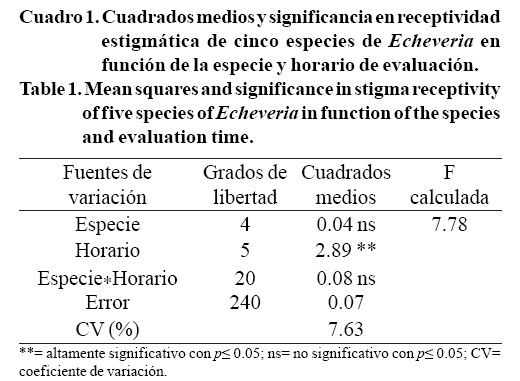
El horario en que hubo mayor porcentaje de estigmas receptivos fue de 12:00 a 14:00 h; además, en estos horarios se observó que el burbujeo fue de mayor duración y las burbujas fueron más grandes, en comparación con las 8:00, 10:00 y 16:00 h, sin diferencias estadísticas entre ellos. A las 18:00 h hubo el menor porcentaje de estigmas receptivos, menor cantidad de burbujas y de menor duración (Figura 4 y 5). Esto concuerda con Consiglio y Bourne (2001), quienes mencionan que el calor puede servir para atraer a los insectos hacia una flor abierta a través de la volatilización del aroma floral durante la antesis, y también ayuda a mantener un período de máxima receptividad del estigma; lo cual explica porque hubo mayor receptividad estigmatica entre las 12:00 y 14:00 h, debido a que fue el periodo en que se registró mayor temperatura ambiental (30 °C).

En otras especies se ha observado que las flores pueden ser receptivas desde antes de abrir hasta el cierre de las mismas. Así, Parés et al. (2002) evaluaron la receptividad del estigma con la metodología de peróxido de hidrógeno en flores de Carica papaya L. y observaron que la receptividad estigmática comenzó antes de la apertura floral. Sin embargo, Ángel-Coca et al. (2011) investigaron la receptividad estigmática en Passif lora edulis con el método de la enzima peroxidasa y obtuvieron resultados positivos desde la apertura floral hasta el cierre de la misma, en tanto que en la presente investigación la receptividad estigmática ocurrió en la apertura floral, y no se observó burbujeo antes del cierre de la flor.
Los resultados de esta investigación indican que las polinizaciones que se realicen entre las 12:00 y las 14:00 h tendrán mayor probabilidad de generar frutos y semillas, pues es cuando la mayoría de las flores abiertas están receptivas.
Tipo de polinización
Amarre de fruto. Se observó que todas las flores evaluadas desarrollaron folículos después de realizar los tratamientos de polinización, por lo cual hubo 100% de amarre de fruto. El porcentaje obtenido en esta investigación fue similar al reportado por Jones et al. (2010) quienes mencionan que en Dudleya multicaulis (Crassulaceae) el amarre de frutos varió desde 86.9 hasta 94.4%, ellos reportan que el alto porcentaje de cuajado de frutos no está limitado por polinizadores; indican que D. multicaulis es capaz de auto-polinizarse en ausencia de vectores de polen; también Jimeno-Sevilla et al. (2014) reportan que el amarre de fruto es más alto en condiciones naturales que en condiciones controladas. Contrario a ésto, Parra et al. (1993) reportan que en E. gibbif lora el amarre del fruto es de 35.5 a 56.6% en promedio, y mencionan que ésto ocurrió aun cuando expusieron el polen al mismo tiempo en que los estigmas estuvieron receptivos.
El alto porcentaje de amarre de fruto no necesariamente indicó que se hallan desarrollado semillas viables; cabe destacar, que en los estudios citados no se evalúo la germinación de las semillas obtenidas para determinar si eran viables; de igual manera no reportan número de semillas por fruto ni el color de las mismas.
Viabilidad de semillas. En todos los frutos de las cinco especies de Echeveria se desarrollaron semillas y en promedio fueron 900; se observó que un mismo fruto hubo semillas cafés y semillas blancas. En los frutos generados por polinización cruzada intra-específica y polinización cruzada inter-específica hubo mayor cantidad de semillas cafés (600 y 900 respectivamente); por el contrario, en la polinización autógama y apomixis hubo mayor cantidad de semillas blancas (700 y 900 respectivamente). Con respecto a las especies, en E. perle y E. runyonii se observó que las semillas eran casi en su totalidad blancas independientemente del tipo de polinización. Con respecto a las semillas, la familia de plantas suculentas produce miles de semillas de color parduzco que son fértiles y semillas blancas que son infértiles o vanas, en cada inflorescencia estas semillas se encuentran dentro de los folículos y se pueden colectar antes de su maduración total.
El análisis de varianza indicó que la viabilidad de las semillas varió en función de la especie, tipo de polinización, así como por la interacción entre ambos, ya que los tres tuvieron efecto altamente significativo (Cuadro 2).
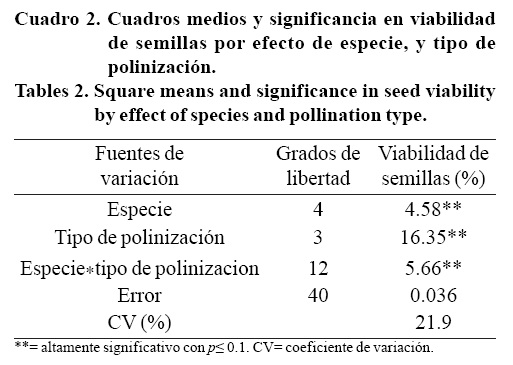
Solo tres de las cinco especies estudiadas produjeron semillas viables (Cuadro 3), E. runyonii y E. perle tuvieron 0% de germinación de semillas, lo que indica que sus gametos no generaron descendencia fértil, por ser un mutante y un híbrido inter-específico respectivamente. Al respecto, Uhl (1982), indica que los gametos de los mutantes e híbridos inter-específicos son en su mayoría estériles.
Por otra parte, se observó que las semillas fueron viables en las polinizaciones alógama intra-específica y alógama inter-específica; en tanto que, en el tratamiento de autopolinización y posible apomixis no germinó ninguna semilla, indicando no viabilidad (Figura 6). Lo anterior indica que en condiciones de invernadero no fue posible la autofecundación y que no existió agamospermia. Sin embargo, Charlesworth y Charlesworth (1987) y Cruden (1976), dicen que es muy común la autofecundación en las Crassulaceae, cabe destacar que estas investigaciónes se realizaron a cielo abierto.

La interaccion especies y tipo de polinización resultó altamente significativa, lo que demuestra que el tipo de polinización tiene un efecto directo en las especies; con base en lo anterior, es claro que hubo sinergia entre E. agavoides, E. elegans y E. pumila con la polinización alógama intra-específica. Algo similar ocurrió con E. agavoides y E. elegans con la polinización alógama inter-específica (Cuadro 3). Lo anterior indicó que los gametos de diferentes plantas de la misma especie generaron descendencia fértil en tres de las cinco especies evaluadas. Los gametos de plantas de diferente especie formaron semilla viable sólo en el caso de E. agavoides y E. elegans; en tanto que, E. runyonii y E. perle no produjeron semilla viable, independientemente del tipo de polinización, lo anterior pudo deberse a que los óvulos son estériles o con baja fertilidad como ocurrió con su polen que tuvo 59 y 59.27% de viabilidad, respectivamente. La escasa fertilidad de los gametos, como ya se mencionó, se debe a que E. runyonii es una mutación generada por rayos gama y E. perle es un híbrido inter-específico; al respecto, Uhl (1982) señala que los gametos de los mutantes e híbridos inter-específicos son estériles en su mayoría. De igual forma, Wyatt (1981) menciona que los híbridos de Sedum no generan semillas viables cuando se realiza un cruzamiento artificial, como se realizó en la presente investigación.
La cantidad de semillas viables fue baja posiblemente porque la cantidad de polen depositado en los estigmas fue insuficiente. Es probable que estas especies generen mayor descendencia si se cultivan en un área abierta para los polinizadores, pues como señala Charlesworth y Charlesworth (1987), Cruden (1976), Carque et al. (1996), Izco et al. (1998) y Jimeno-Sevilla et al. (2013), la polinización de las Crassulaceae ocurre por insectos, aves, viento, y agua; la cantidad de polen disponible sería mayor. Al respecto, Parra-Tabla et al. (1998) indican que el amarre de semillas puede disminuir por efecto de la disponibilidad de polen, más aun cuando la polinización es manual.
Aunque se reporta que puede haber autopolinización (Charlesworth y Charlesworth, 1987 y Cruden, 1976), los resultados obtenidos indican que la polinización en Echeveria fue cruzada, lo cual concuerda con Izco et al. (1998); Jimeno-Sevilla et al. (2013); Jimeno-Sevilla et al. (2014).
También Rojas-Aréchiga y Batis (2001) mencionan que la baja viabilidad de semillas se puede deber a que el embrión no se encuentra totalmente desarrollado al momento de su dispersión, por lo que se necesita de un periodo de tiempo para completar su maduración; cabe señalar que en la presente investigación, la semilla fue cosechada cuando los folículos estaban abriendo y las semillas habían madurado. De igual manera, Rojas-Aréchiga et al. (1998) señalan que la baja germinación se puede deber a que las especies suculentas desarrollan diversos mecanismos de germinación para poder persistir en ambientes desérticos. Sin embargo, en esta investigación se aplicó AG3 (5 ppm) porque en un primer ensayo de siembra no hubo germinación de semillas, lo cual evidencio presencia de algún tipo de inhibidor de la germinación.
Con los resultados anteriores se pudo conocer el tipo de polinización de las cinco especies de Echeveria estudiadas, para poder proponer un método de mejoramiento genético y poder generar nuevas variedades mediante hibridación, y poder introducir al comercio ornamental especies generadas en México.
Conclusiones
Con base en los resultados obtenidos y las condiciones en que se llevó a cabo el estudio, se concluye que la viabilidad de polen de cinco especies de Echeveria varió de 59.2 a 72.7%. El momento de receptividad del estigma de las cinco especies de Echeveria ocurrió de las 12:00 a las 14:00 h. Solo los cruzamientos intra e inter específicos generaron semillas con capacidad germinativa; lo que indica que las Echeverias son de polinización cruzada. Las especies de E. perle y E. runyonii no fomaron semillas, por lo cual no pueden ser parte de un programa de mejoramiento genético, fungiendo como hembras.
Agradecimientos
El primer autor agradece a la Facultad de Ciencias Agropecuarias de la Universidad Autónoma del Estado de Morelos, y al Consejo Nacional de Ciencia y Tecnología (CONACYT), por su apoyo a través de la beca, y su valioso apoyo para la realización de los estudios de doctorado.
Literatura citada
Ángel-Coca, C.; Nates-Parra, G.; Ospina-Torres, R. y Melo, C. D. O. 2011. Biología floral y reproductiva de la gulupa Passif lora edulis Sims. F. edulis. Caldasia 33:433-451. [ Links ]
Anónimo. 2013. Especies de Crassulaceae. http://www.worlfieldguide.com. [ Links ]
Arreguín-Sánchez, M. L.; Palacios-Chávez, R. y Quiroz-García, D. L. 1990. Morfologia de los granos de polen de los géneros Echeveria, Sedum y Villadia (Crassulaceae) del Valle de México. Cactáceas y Suculentas Mexicanas 3: 51-61. [ Links ]
Carque, A. E.; Arco, A. M.; León, A. M. C. y Wilpret, T. W. 1996. Estudio seminal germinativo y de plántulas de la flora vascular endémica canaria. I. Crassulaceae (1a parte). Vieraea 25:7-30. [ Links ]
Charlesworth, D. and Charlesworth, B. 1987. Inbreeding depression and its evolutionary consequences. Ann. Rev. Ecol. Systematics 18:237-268. [ Links ]
Consiglio, T. K. and Bourne, G. R. 2001. Pollination and breeding system of a neotropical palm Astrocaryum vulgare in Guyana: a test of the predictability of syndromes. J. Trop. Ecol 17:577-592. [ Links ]
Cruden, R. W. 1976. Pollen-ovule ration: a conservative indicator of breeding systems in flowering plants. Evolution 31:32-46. [ Links ]
Eggli, U. 2003. Illustrated Handbook of succulent Plants: Crassulaceae. Springer. Zürich Switzerland. 458 p. [ Links ]
Espinosa-Osornio, G. y Engleman, E. M. 1998. Breve recopilación de anatomía de semillas. Colegio de Posgraduados en Ciencias Agrícolas. Texcoco, Estado de México. 43 p. [ Links ]
García, E. 1981. Modificaciones al sistema de clasificación climática de Koppen (para adaptarla a las condiciones de la República Mexicana): Universidad Nacional Autónoma de México (UNAM), Instituto de Geografia. México, D. F. 246 p. [ Links ]
Huang, T. C. 1972. Pollen flora of Taiwan National Taiwan University Botany Department Press. 276 p. [ Links ]
Izco, J.; Barreno, E.; Brugués, M.; Costa, M. y Devesa, J. 1998. Botánica. McGraw-Hill Interamericana. Madrid, España. 781 p. [ Links ]
Jimeno-Sevilla, H. D.; Hernández-Ramírez, A. M and Krömer, T. 2013. Echeveria rosea Lindley (Crassulaceae): a hummingbird-dependent succulent epiphyte. Cactus and Succulent Journal. 85:128-131. [ Links ]
Jimeno-Sevilla, H.; Hérnandez-Ramírez, A. M.; Ornelas, J. F. and Marten-Rodríguez, S. 2014. Morphological and néctar traits in Echeveria rosea Lindley (Crassulaceae) linked to humminbird pollination in Central Veracruz, Mexico. Haseltonia 19:17-25. [ Links ]
Jones, C. E.; Shropshire, F. M.; Allen, R. L.; and Atallah, Y. C. 2010. Pollination and reproduction in natural and mitigation populations of the many-stemmed Dudleya, Dudleya multicaulis (Crassulaceae). California Botanical Society 57:42-53. [ Links ]
Karaer, F.; Celep, F. and Kutbay, H. G. 2010. Morphological, ecological and palynological studies on Sempervivum sosnowskyi Ter-Chatsch. (Crassulaceae) with a new distribution record from Turkey. Aust. J. Crop Sci. 4:247-251. [ Links ]
Kearns, C. A.; and Inouye, D. W. 1993. Techniques for pollination biologist. University press of Colorado. Niwot, Colorado. 583 p. [ Links ]
Márquez, G. J.; Collazo, O. M.; Martínez, G. M.; Orozco, S. A. y Vázquez, S. S. 2013. Biología de angiospermas. Universidad Nacional Autónoma de México (UNAM). México, D. F. 755 p. [ Links ]
Nyffeler, R. 1992. A taxonomic revisión of the genus Monanthes (Crassulaceae). Bradleya. 10:49-82. [ Links ]
Osborn, M.; Kevan, P. and Meredith, A. 1988. Pollination biology of Opuntia polyacantha and Opuntia phaceacantha (Cactaceae) in souther Colorado. Plant. Syst. Evol. 159:139-144. [ Links ]
Parada, P. D. M. y Villegas, A. M. 2009. Propagación in vitro del híbrido almendro x durazno h1. Rev. Fitotec. Mex. 32:103-109. [ Links ]
Parés, J.; Basso, C. y Jáureguii, D. 2002. Momento de antesis, dehiscencia de anteras y receptividad estigmática en flores de lechosa (Carica papaya L.) Cv. Cartagena Amarillas. Bioagro 14:17-24. [ Links ]
Parnell, J. 1991. Pullen morphology of Jovibarba Opiz and Sempervivum L. (Crassulceae). Kew Bulletin. 46:733-738. [ Links ]
Parra, V.; Vargas, C. F. and Eguiarte, L. E. 1993. Reproductive biology, pollen and seed dispersal, and neighborhood size in the hummingbird-pollinated Echeveria gibbif lora (Crassulaceae). Am. J. Bot. 80:153-159. [ Links ]
Parra-Tabla, V.; Vargas, C. F. and Eguiarte, L. E. 1998. Is Echeveria gibbiflora (Crassulaceae) fecundity limited by pollen availability? An experimental study. Funcional Ecol. 12:591-595. [ Links ]
Rojas-Aréchiga, M.; Vázquez-Yanes, C. and Orozco-Segovia, A. 1998. Seed responses to temperatura of Mexican cacti species from two life forms: an ecophysiological interpretation. Plant Ecol. 135:207-241. [ Links ]
Rojas-Aréchiga, M. y Batis, A. I. 2001. Las semillas de cactáceas…forman bancos en el suelo? Cactáceas y Suculentas Mexicanas 46:76-82. [ Links ]
Sajeva, M.; and Constanzo, M. 1997. Succulents the illustrated dictionary. Ed. Timber Press. Portland, Oregon. 239 p. [ Links ]
Shivanna, K. R. and Sawhney, V. K. 1997. Pollen biotechnology for crop production and improvement. 28-29 pp. [ Links ]
Thiede, J. 1995. Quantitative phytogeography, species richness, and evolution of American Crassulaceae. In: evolution and systematics of the Crassulaceae, Henk’t Hart y Urs Eggli (Eds.). Backhuys, Leiden. 89-123 pp. [ Links ]
Toledo, V. M. 1988. La diversidad biológica de México. Ciencia y Desarrollo 81:17-30. [ Links ]
Uhl, C. H. 1982. The problem of ploidy in Echeveria (Crassulaceae): I diplody in E. ciliata. Am. J. Bot. 62:843-854. [ Links ]
Wyatt, R. 1981. Ant.- pollination of the granite outcrop Endemic Diamorpha smallii (Crassulaceae). Am. J. Bot. 68:1212-1217. [ Links ]

