











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La agricultura es uno de los sectores más vulnerables a los impactos del cambio climático (FAO, 2007). La agricultura de temporal y aún la agricultura de riego, son sensibles a los cambios en los patrones climáticos regionales, incluso a escala interanual. Las predicciones climáticas del Siglo XXI (IPCC, 2007) no son promisorias y más bien señalan que el cambio climático se acentuará durante el transcurso del presente siglo y establecerá condiciones más extremosas para los cultivos, tanto en condiciones de temperatura como de humedad, por lo que será necesario generar medidas de adaptación a las nuevas condiciones climáticas. Una de las medidas más importantes que se visualizan, es la generación de nuevas variedades, las cuales se prevé deberán adaptarse a ambientes más cálidos y probablemente menos húmedos (IPCC, 2007; Zarazúa, 2011).
Ante este panorama, es muy probable que se requiera la formación de nuevas poblaciones genéticas para alimentar la generación de genotipos con tales características. En este proceso, la utilización de parientes silvestres de los cultivos será fundamental, ya que se sabe que en estos recursos fitogenéticos se encuentran genes asociados a una mayor rusticidad con relación a varios factores ligados a la producción de cultivos, tales como resistencia a sequía (Ruiz et al, 2008), resistencia a altas y bajas temperaturas (López et al, 2005; Ruiz et al, 2013), y resistencia al ataque de plagas y enfermedades (De la Paz et al, 2010), entre otros.
La presencia y distribución de las especies silvestres está estrechamente relacionada con sus requerimientos agroclimáticos, y como estos requerimientos son satisfechos en los diversos ambientes (FAO, 2007). La presencia del cambio climático puede constituir una amenaza para la satisfacción de estos requerimientos y por lo tanto para la presencia y distribución de las especies silvestres (Jarvis et al, 2008).
México, es considerado como uno de los países con mayor diversidad biológica (Mittermeier y Goettsch, 1992), debido entre otros factores a su gran diversidad climática (García, 2004). Actualmente se estima que 12% de la biodiversidad total mundial se localiza en México; de esta biodiversidad, 12% de los géneros y 50-60% de las especies de plantas son endémicas (CONABIO, 2006).
Con la presencia del cambio climático, la diversidad de climas del país se puede ver alterada, de tal forma que puede modificarse la composición de los ecosistemas (Lasco et al., 2008; Schneider et al, 2009), así como la distribución geográfica de los recursos fitogenéticos del país. Gossypium hirsutum es considerado uno de estos recursos, ya que México junto con Centroamérica, es considerado centro de origen de esta especie (Jenkins, 2003). La importancia de este recurso consiste en que actualmente 90% de la producción mundial de algodón se obtiene a partir de G. hirsutum. Probablemente los pobladores nativos de la Península de Yucatán fueron los primeros en domesticar G. hirsutum (Brubaker y Wendel, 1994). Estas formas iniciales de algodón semi-domesticado se dispersaron al resto de Mesoamérica y hacia el interior del Caribe (Iqbal et al, 2001). A partir de aquí, ocurrió una selección hacia una latencia de semilla reducida, un hábito de crecimiento anual, y una floración independiente del fotoperiodo, creando genotipos muy similares a los cultivares actuales (OGTR, 2008).
El objetivo de la presente investigación fue determinar el impacto potencial del cambio climático sobre los nichos de distribución geográfica de G. hirsutum L. bajo un escenario de referencia y de cambio climático con tres modelos de circulación general (MCG): ECHAM5, MIROC (Medres) y UKMO_HADCM3, bajo el escenario de emisiones de gases de efecto invernadero A2. Para esto, se utilizan algoritmos o modelos de nichos de distribución.
Uno de estos modelos es Maxent, el cual es un método de inteligencia artificial que aplica el principio de máxima entropía para calcular la distribución geográfica más probable para una especie. El resultado del modelo expresa el valor más adecuado del hábitat potencial para la especie en función de las variables ambientales utilizadas. Un valor alto de la función de distribución en cada pixel indica que éste presenta condiciones favorables para encontrar dicha especie (Phillips et al., 2008; Morales, 2012). Dicho modelo ha sido utilizado recientemente para determinar tanto la distribución de especies de plantas, animales e insectos (Contreras et al., 2010; González et al., 2010; Ibarra et al, 2010; Sork et al., 2010; Torres y Jayat, 2010; Paredes et al, 2011; Sobek et al, 2012).
La comparación entre los resultados obtenidos de los tres diferentes MCG, dará una idea de la tendencia de la distribución de la especie en estudio.
Materiales y métodos
Bases de datos
Se utilizó una base de datos de 387 accesiones geo-referenciadas de G. hirsutum L., las cuales se realizaron entre 1980 y 2011 por parte de nueve instituciones: Instituto Nacional de Investigaciones ForestalesAgrícolas y Pecuarias (INIFAP), Instituto Nacional de Ecología (INECOL), Universidad Autónoma de México (UNAM), Centro de Investigaciones Científicas de Yucatán (CICY), Departamento deAgricultura de Estados Unidos (USDA), Universidad Autónoma de Yucatán (UAY), Universidad Autónoma de Chapingo (UACH), Colegio de Postgraduados en Ciencias Agrícolas (CP) y Jardín Botánico de Missouri (JBM).
Para la caracterización de las condiciones ambientales de los sitios de colecta, se utilizó la climatología de WorldClim correspondiente al periodo climático 1960-1990. Para la modelación de los nichos de distribución de G. hirsutum se utilizó la climatología anteriormente mencionada y el escenario de cambio climático 2040-2069 donde se consideraron tres MCG (ECHAM5, MIROC (Medres) y UKMO_HADCM3) considerando un escenario de emisiones de gases de efecto invernadero tipo A2. Todos estos con una resolución espacial de 2.5 minutos. Ambos sistemas de información geográfica se compilaron en el software IDRISI Selva (Eastman, 2013). Además de estas variables, se incluyeron otras variables de diagnostico de tipo edáficas y topográficas provenientes del sistema de información ambiental (SIAN) del Instituto Nacional de Investigaciones ForestalesAgrícolas y Pecuarias (Díaz et al, 2012).
Variables de estudio
A partir de las coordenadas geográficas de los sitios de colecta, se generó un archivo vectorial y, con los archivo raster correspondientes a la climatología de referencia, se hizo uso del módulo GIS Analysis del sistema IDRISI Selva (Eastman, 2013), para extraer información puntual por accesión, relacionada con las siguientes variables: precipitación acumulada media anual (PAA), temperatura media anual (TMA), temperatura mínima media anual (TIA), temperatura máxima media anual (TXA), temperatura máxima media del mes más cálido (TXMMC), temperatura mínima media del mes más frío (TIMMF) y altitud (A).
Para la modelación de los nichos de distribución se utilizaron las variables anteriormente mencionadas, más las siguientes: pendiente, oscilación térmica anual, oscilación térmica, precipitación, temperatura mínima media, temperatura media y temperatura máxima media de los períodos mayo-octubre y noviembre-abril.
Modelación de la distribución geográfica
Para determinar la distribución geográfica potencial de G. hirsutum se utilizó el modelo Maxent (Phillips et al., 2006) versión 3.3.3 k, el cual ha sido evaluado y probado como uno de los mejores modeladores de nichos de distribución (Elith et al, 2006). Dicho software se basa en el principio estadístico de máxima entropía, el cual calcula la probabilidad de distribución de máxima entropía (la distribución más próxima a la uniforme), a partir de datos de presencia (colecta) y ausencia (lugares sin datos), ajustando la distribución de especies al conjunto de pixeles del área de estudio y tomando como principio la idea de que la mejor explicación a un fenómeno desconocido maximizará la incertidumbre de la distribución de la probabilidad dependiendo de aquellos pixeles en los cuales la especie ha sido detectada (Phillips et al, 2004, 2006). El resultado final es una expresión del tipo:
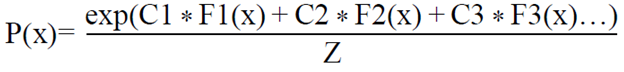
Donde= C1, C2.. .Cn, son constantes; F1, F2.. .Fn, son las variables climáticas y Z es una constante que asegura que la suma de todos los valores de P para todas las celdas sea igual a 1.
El resultado se puede dar por tres tipos de salidas: crudo, cumulativo y logístico, para el presente caso se utilizó la salida de tipo logística, la cual proporciona como resultado un mapa de distribución potencial donde el rango máximo de aptitud ambiental se ubica entre 0 y 1.
Descriptores ecológicos
Se determinaron los descriptores ecológicos de G. hirsutum a partir de todos los puntos de colecta utilizados para el análisis y los rangos climáticos en los que se establecen. Esto se realizó utilizando el sistema IDRISI Selva, las imágenes raster de cada variable ambiental y las coordenadas geográficas de cada punto de colecta. Una vez obtenidos estos datos, se buscaron los valores extremos para establecer los rangos ambientales en los que se establece la especie.
Resultados y discusión
En el Cuadro 1 se describen algunas estadísticas básicas de variables ambientales de los sitios de accesión de G. hirsutum. Como puede verse, los rangos ambientales de distribución de esta especie son amplios. Esto señala la rusticidad de esta especie, característica que podría ser favorable en ambientes donde el cambio climático se manifieste de manera más agresiva (FAO, 2007).
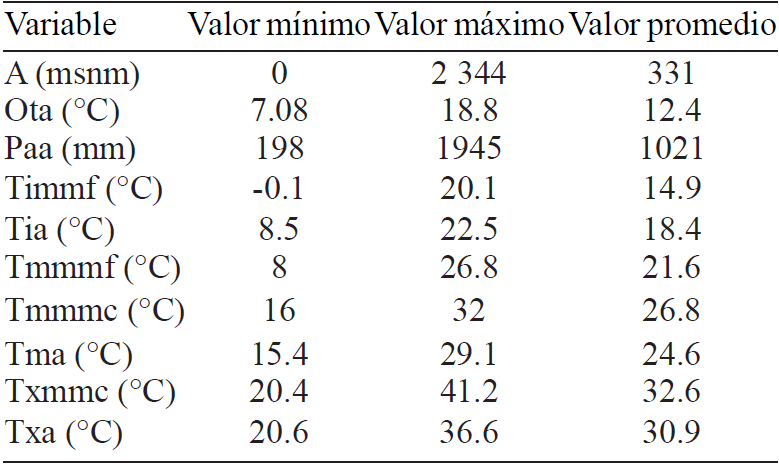
El valor máximo, el valor mínimo y el valor promedio de cada variable expresada en el Cuadro 1 constituyen también los descriptores ecológicos de G. hirsutum. De acuerdo con las accesiones analizadas, el rango de precipitación anual varía entre 198 y 1945 mm, valores que contrastan (sobre todo el valor mínimo) con lo señalado por Aragón (1995) quien establece un rango de 750 a1 500 mm anuales.Asimismo, los límites altitudinales resultado del análisis de las accesiones estudiadas, reportan que el rango de distribución de G. hirsutum va de los 0 a 2 344 m, valores que difieren con lo reportado por Baradas (1994), quien señala que el rango de distribución altitudinal es de 0 a 600 m.
Así mismo puede observarse que la especie se adapta a un gran rango climático, ya que puede encontrársele en sitios donde la temperatura varía desde los -0.1 hasta los 41.2 °C.
En la Figura 1, se muestran las accesiones utilizadas para la modelación de G. hirsutum con el software Maxent, donde claramente puede observarse que la especie se distribuye en la región subhúmeda cálida del país, presentándose algunas accesiones en la región árida.

Este algoritmo realiza algunas pruebas estadísticas a partir de 25% de datos asignados en el random test. Una de las estadísticas realizadas es referente a la sensibilidad especificidad de los datos para la generación del modelo, Receiver Operating Characteristic (ROC); es decir, la razón de verdaderos positivos frente a la razón de falsos positivos, correspondiente a la probabilidad de que tomando al azar un par de valores, uno positivo (pixel con presencia de la especie) y otro negativo (pixel con ausencia de la especie), el modelo adjudique al valor positivo un valor mayor (Hanley, 1982). Dicha prueba muestra que el AUC (área bajo la curva ROC) para los datos de entrenamiento es del 0.937 (datos empleados en la generación del modelo), esto significa que el modelo se ajusta casi perfectamente.
Cabe mencionar que la especie se adapta a regiones áridas y semiáridas con climas cálidos y semicálidos (Aragón 1995; Ruiz et al., 1999), aunque en el mapa puede observarse que las accesiones se encuentran en su mayoría en la región cálida húmeda del país.
En la Figura 2 (mapa a) se muestra el mapa resultante del análisis de aptitud ambiental clasificada por rangos, para la climatología de referencia. La clasificación se hizo en 5 intervalos de aptitud: muy alta, alta, media, baja y muy baja, correspondientes a rangos de aptitud de 0.20 a partir del resultado logístico arrojado por Maxent (0-1), siendo las zonas de muy baja aptitud ambiental todo aquello que se encuentra entre 0-0.20 y las de muy alta aptitud aquellas que se encuentran en rangos de 0.80-1.
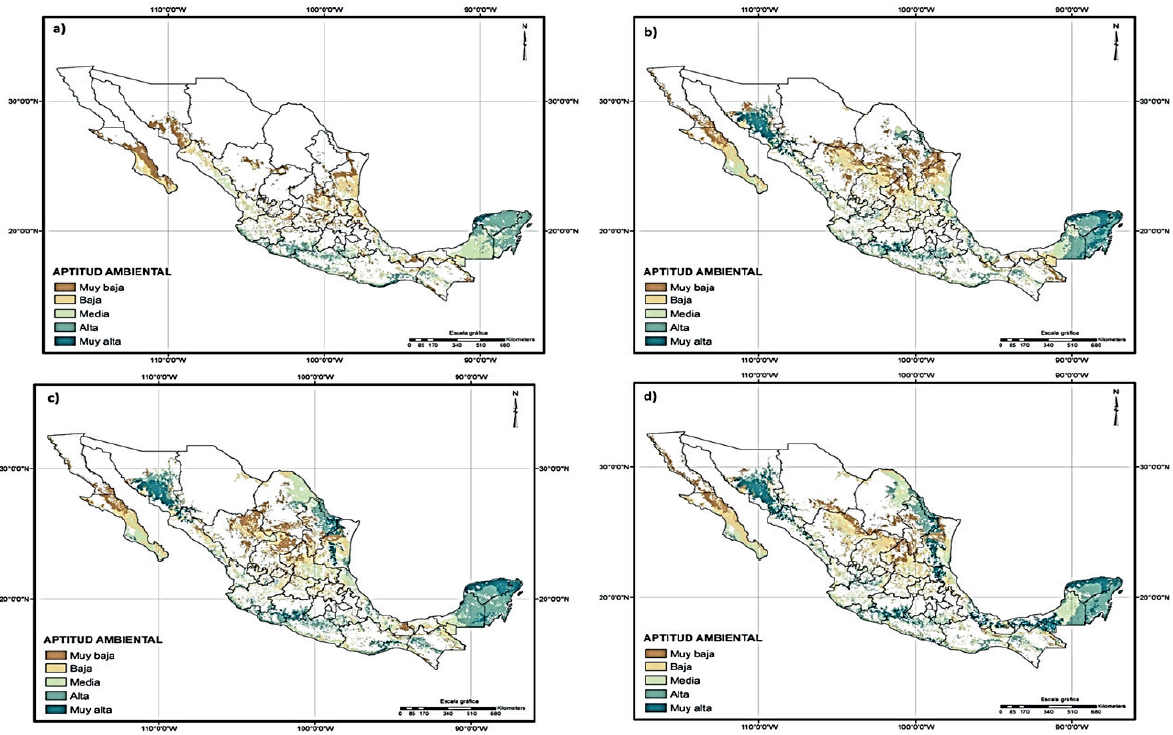
Figura 2 Aptitud ambiental de G. hirsutum en: a) climatología de referencia periodo 1960-1990; b) climatología 2040-2069 MCG ECHAM5; c) climatología 2040-2069 MCG UKMO_HADCM3; y d) climatología 2040-2069 MCG MIROC (Medres).
Por otra parte, los mapas b, c y d de la Figura 2, muestran la distribución geográfica potencial de G. hirsutum bajo el escenario de cambio climático 2040-2069 para los diferentes MCG empleados en el presente estudio, los cuales presentan resultados muy contrastantes con la situación actual y muy similares entre sí (Cuadro 2).
Cuadro 2 Porcentaje de ocupación territorial de G. hirsutum L. para el escenario de referencia y ocupación territorial potencial para el escenario de cambio climático 2040-2069 con tres MCG.
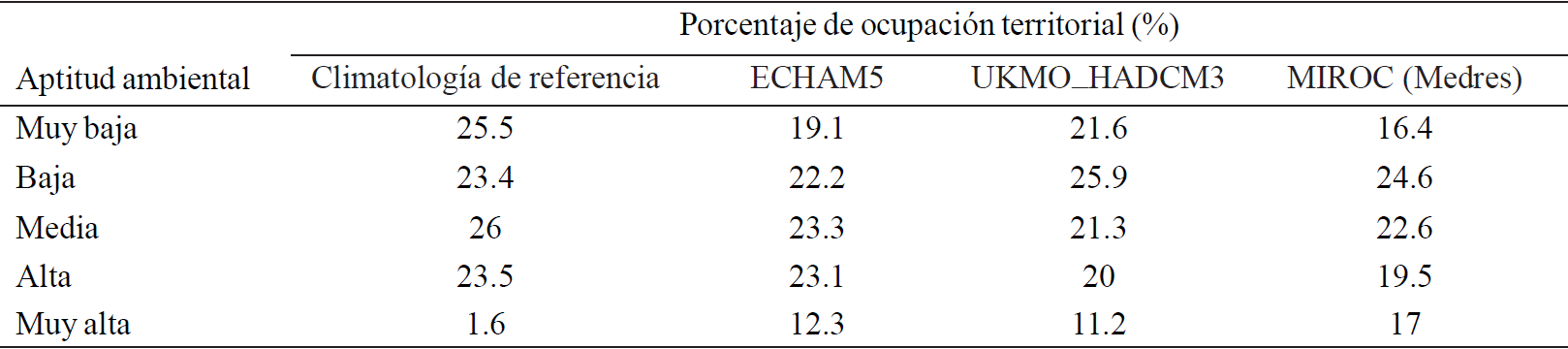
Cabe mencionar que a los porcentajes mencionados en el Cuadro 2 se les restó las superficies de los cuerpos de agua, manchas urbanas y todos aquellos tipos de vegetación donde G. hirsutum no se puede establecer, como bosques de coníferas y vegetación riparia, por mencionar algunos. Esta capa de información proviene de la carta digital de Uso de suelo y vegetación (INEGI, 2005).
De manera general se puede observar que respecto al escenario de referencia, todos los modelos presentan la misma tendencia, esto es dismunucion de la categoría muy baja y aumento en las categorías de aptitud ambiental baja, media, alta y muy alta.
El modelo que presentó mayor disminución en la superficie de la categoría muy baja fue el UKMO_HADCM3 con una disminución de 21.6%, siendo el modelo MIROC el que presentó menor disminución respecto a la climatología de referencia en esta categoría (16.4%). Respecto a las demás categorías MIROC presenta mayor ganancia en superficie en la categoría muy alta y ECHAM5 presenta mayor ganancia en las categorías media y alta.
Los modelos que presentan menor ganancia en porcentaje de superficie ocupada por categoría son ECHAM5 en la categoría baja, UKMO_HADCM3 en la categoría media y muy alta y MIROC (Medres) en la categoría alta.
De manera general se puede observar que todos los modelos muestran una tendencia en la cual la especie ampliará su distribución con las condiciones de cambio climático, extendiéndose a los estados de Sonora, Sinaloa, Tamaulipas y Nuevo León.
De lo mencionado anteriormente es evidente que el impacto ocasionado por el cambio climático beneficiará la distribución de G. hirsutum L., a diferencia de muchas otras especies cuya distribución geográfica se verá afectada por los cambios en los patrones climáticos (Jarvis, 2008; Villers, 2009; Contreras et al., 2010; Sork et al., 2010). Dentro del beneficio que el cambio climático traerá a G. hirsutum se encuentra la expansión de las áreas con alta y muy alta aptitud ambiental, lo cual provocará la aparición de nuevas áreas con este nivel de aptitud ambiental, sobre todo en la región formada por el sur del estado de Sonora y norte del estado de Sinaloa, la zona formada por la zona de colindancia de Tamaulipas y Nuevo León y la región del Golfo Sur a nivel del estado de Tabasco (Figura 2).
Estudios realizados en el suroeste de Estados Unidos de América, demuestran que los campos para cultivo de algodón (Gossipyum sp.) incrementarán hasta en 26-36% de su condición actual, en caso de que los niveles de dióxido de carbono continúen incrementando (Doherty, 2003).
Los procesos de las plantas que son directamente afectados por cambios en la atmósfera (concentración de CO2) son principalmente la fotosíntesis, foto-respiración, respiración nocturna y transpiración (Fitter, 1987). El aumento de la temperatura bajo el cambio climático futuro, así como los niveles elevados de CO2 serán benéficos para la productividad del algodón hasta cierto umbral, en donde la tasa de fotosíntesis sea menor que la óptima o el estrés ambiental sea alto (Reddy et al., 2000).
Conclusiones
Para los escenarios climáticos futuros, se espera que la distribución de la especie se vea favorecida, debido a que el cambio climático incrementará las zonas con mayor aptitud ambiental para G. hirsutum. Sobre esta conclusión existe coincidencia en los tres MCG utilizados, siendo el modelo MIROC (Medres) el que reporta mayor superficie de muy alta aptitud ambiental.
Por lo anterior, es importante identificar aquellas zonas que se verán favorecidas o afectadas y comenzar a implementar las estrategias necesarias para asegurar la conservación de la especie como recurso fitogenético.
En cuanto a la producción de algodón a partir de G. hirsutum, es muy probable que por el momento no se requieran variedades adicionales a las ya disponibles, pero es un mediano plazo habrá que ir desarrollando estrategias enfocadas en el desarrollo de variedades que fotosinteticen eficientemente a altas temperaturas y en condiciones de baja disponibilidad de humedad en el suelo, con la finalidad de adaptarse ante cualquier escenario climático.

