










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.5 no.1 Texcoco ene./feb. 2014
Notas de investigación
Impacto de la salinidad y la temperatura diurna sobre la fluorescencia de la clorofila en fresa*
Impact of diurnal temperature and salinity on chlorophyll fluorescence in Strawberry
Nazario Francisco-Francisco1 y Adalberto Benavides-Mendoza1§
1 Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1923. Buenavista, Saltillo, Coahuila, 25315 México. (fafnaz@hotmail.com). §Autor para correspondencia: abenmen@uaaan.mx.
* Recibido: agosto de 2013
Aceptado: diciembre de 2013
Resumen
Este estudio fue realizado para investigar el comportamiento de la fluorescencia de la clorofila de plantas de fresa (Fragaria x ananassa) ante la salinidad y la temperatura diurna bajo invernadero. Para ello, se establecieron plantas de fresa en macetas de polietileno negro con una capacidad de 5 L conteniendo turba de musgo como sustrato. Los tratamientos de salinidad fueron inducidos por la aplicación de CaCl2 a 0 y 12 mM el cual fue adicionado a una solución nutritiva Steiner. Las plantas testigo y las tratadas crecieron en un invernadero ventilado con extractores de aire y fueron irrigadas cada dos días. La investigación del impacto de la temperatura diurna sobre la fluorescencia de la clorofila fue realizado registrando la temperatura ambiente del invernadero cada hora entre las 8:00 am a 5:00 pm. Las mediciones de los parámetros de fluorescencia fueron realizadas en los días 0, 14, 28, y 42 después del inicio de los tratamientos de salinidad. El tratamiento de 12 mM de CaCl2 redujo el rendimiento cuántico máximo (Fv/Fm) y efectivo (ePS2), pero no aumentó el decaimiento no fotoquímico (NPQ) interpretándose esta respuesta como una mínima afectación al PSII. En cuanto a la temperatura las plantas mostraron disminución en la Fv/Fm y ePS2 con temperaturas del aire superiores a los 28 °C. La máxima eficiencia fotoquímica se observó con temperaturas menores a 19 °C.
Palabras clave: eficiencia fotoquímica, fotoinhibición, fotoprotección.
Abstract
This study was conducted to investigate the behavior of the chlorophyll fluorescence of plants of strawberry (Fragaria x ananassa) with the daytime temperature salinity and low emissions. For this, strawberry plants were established in pots of black polyethylene with a capacity of 5 L containing peat moss as substrate. Salinity treatments were induced by the application of CaCl2 at 0 and 12 mM, which was added to a nutrient solution Steiner. The control and the treated plants grown in a greenhouse ventilated with exhaust fans and were irrigated every two days. The investigation of the impact of diurnal temperature on chlorophyll fluorescence recording was made greenhouse temperature every hour between 8:00 am to 5:00 pm. Measurements of the fluorescence parameters were performed on days 0, 14, 28, and 42 after initiation of salinity treatments. Treatment of 12 mM ofCaCl2 reduced the maximum quantum efficiency (Fv / Fm) and effective (EPS2) but did not increase the decay non-photochemical (NPQ) is interpreted as a response to the PSII minimal involvement. As to the temperature decrease in the plants showed the Fv/Fm and EPS2 air temperatures exceeding 28 °C. The maximum photochemical efficiency was observed with temperatures below 19 °C.
Key words: photochemical efficiency, photoinhibition, photoprotection.
Al adaptarse a factores ambientales como la salinidad, el déficit hídrico y extremos de temperatura, las plantas llevan a cabo ajustes en las actividades metabólicas (Hughes y Smith, 2007). Siendo la radiación electromagnética la fuente de energía primaria, es de esperarse que varios de los citados cambios ocurran en los sistemas de pigmentos fotosintéticos. Los ajustes en el sistema de captura de fotones son medibles mediante una técnica denominada fluorescencia modulada de la clorofila, que consiste en la aplicación de cuatro fuentes de radiación con balance espectral e irradiancia diferentes para lograr distintos estímulos y respuestas del aparato fotoquímico, ello en un pequeño volumen del tejido foliar. El registro de las respuestas es utilizado para calcular los parámetros con los que se define la eficiencia fotoquímica la cual depende de los cambios en las variables ambientales (Baker, 2008).
Cada factor ambiental modifica de manera particular el aparato fotoquímico (Baker, 2008). La salinidad provoca por una parte la modificación de la estructura de las membranas tilacoides y por otro lado inhibe parcialmente el ciclo de Calvin (Jaramillo-Giraldo et al, 2009), disminuyendo el rendimiento cuántico. Las temperaturas altas modifican la fluidez de la membrana, alterando el funcionamiento de las proteínas transportadoras de electrones (Xu et al, 2011). La fresa (Fragaria sp.) con poca capacidad de tolerancia frente a factores ambientales como la salinidad (Casierra-Posada y García-Riaño, 2006) y los extremos de temperatura (Kadir y Sidhu, 2006) es entonces un modelo biológico útil para describir desde el punto de vista ecofisiológico los ajustes fotoquímicos frente a éstos factores ambientales adversos. Por otro lado, diferentes sales con contenido de calcio como el CaCl2 han sido usadas como fertilizantes en plantaciones de fresa. También ha sido documentado el efecto positivo de esta sal en la firmeza de sus frutos (Lanauskas et al., 2006). No obstante, poca información existe acerca de los efectos negativos del CaCl2, a diferencia de otras sales más estudiadas como el NaCl (Bisko et al, 2010). El objetivo de este estudio fue investigar el efecto de la salinidad y las variaciones en la temperatura ambiente en la eficiencia fotoquímica de las plantas de fresa var. Festival considerando los parámetros Fo, Fs, Fm, Fm', Fv/Fm, ePS2, qP, y NPQ.
Material vegetal y tratamientos. Las plantas de fresa, cultivar "Festival" fueron establecidas en macetas de polietileno de 5 L con un sustrato de turba de musgo (Berger BM2) más perlita (Hortiperlita) a una proporción de 75:25 en volumen. Los estolones fueron establecidos el 15 de octubre de 2011 en un invernadero ventilado con extractores de aire y ventilación pasiva lateral con temperatura diurna máxima de 35°C y temperatura nocturna mínima de 15°C y humedad relativa promedio de 50%. La fertilización de las plantas fue en base a una solución nutritiva de Steiner a 15% más elementos menores a 100% (Steiner, 1961), dando una concentración de 0.372 g/L de fertilizantes. Para inducir la salinidad se aplicó un tratamiento de cloruro de calcio (CaCl2) a una concentración de 0 y 12 mM que se mezcló con la solución nutritiva resultando una concentración final de 0.372 y 1.704 g/L y una conductividad eléctrica de 555 y 2543 (μS•cm-1 respectivamente. Previo a ello, se llevó a cabo una determinación preliminar del comportamiento de los parámetros de fluorescencia frente a las concentraciones de 0, 3, 6, 9 y 12 mM de CaCl2, en la que no se observó diferencia significativa en las dosis de 3, 6 y 9 mM en los primeros 3 días de la aplicación, razón por la que se decidió continuar con las determinaciones únicamente del testigo y de la concentración que mostró diferencia significativa, que fue la más alta (12 mM).
Cabe señalar que la conductividad alcanzada con la dosis de 12 mM de CaCl2 es superior a la máxima recomendada en el cultivo de fresa que es de 2000 (μS•cm-1 (Barroso y Alvarez, 1997). Se aplicaron diariamente 150 mL de éstas mezclas a las macetas designadas. El diseño experimental utilizado en este ensayo fue un factorial a x b conducido bajo un arreglo completamente al azar con 3 plantas por tratamiento, dando un total de 6 plantas, a las que se les tomó 1 pulso de fluorescencia en los 0, 14, 28, y 42 días posteriores de comenzado el tratamiento. Cada medición fue tomada entre las 8:00 am y 8:30 h a una temperatura que oscilo entre 18-21 °C. El registro de los parámetros para verificar el efecto de los cambios en la temperatura ambiente se realizó en 3 plantas a intervalos de una hora, 4 veces en un día, entre las 8:00 y las 17:00 h, en tres días consecutivos con condiciones similares de temperatura que oscilaron entre los 18-34 °C. La temperatura se verificó con un termómetro ubicado a la altura del dosel de las plantas.
El diseño experimental utilizado en éste ensayo fue un arreglo completamente al azar. Sobre los datos se realizó un análisis de varianza y una prueba de medias con Tukey (p≤ 0.05). Previo a cada medición las hojas fueron adaptadas a un período de obscuridad por 30 min con el dispositivo incluído en el fluorómetro. Todas las mediciones se realizaron en los foliolos con madurez fisiológica. Para las mediciones de la fluorescencia de clorofila se utilizó un fluorómetro de pulso modulado (Fluorescence Monitoring System [FMS-2]; Hansatech Instruments Ltd., Norfolk, United Kingdom).
La fluorescencia mínima (Fo) se obtuvo al aplicar una luz modulada débil a una intensidad de 0.1 μmol de fotones m-2 s-1 y la fluorescencia máxima con la aplicación de un pulso de 18 000 μmol de fotones m-2 s-1 con una duración de 0.8 segundos. A la misma sección de tejido foliar se le aplicó durante cuatro minutos luz actínica con irradiancia de 1 020 μmol de fotones m-2 s-1 determinando el estado estable de la fluorescencia (Fs) así como la fluorescencia máxima (Fm'). Los parámetros adicionales obtenidos del equipo fueron: rendimiento cuántico máximo (Fv/Fm), rendimiento cuántico efectivo (ePS2), decaimiento fotoquímico (qP), y decaimiento no fotoquímico (NPQ).
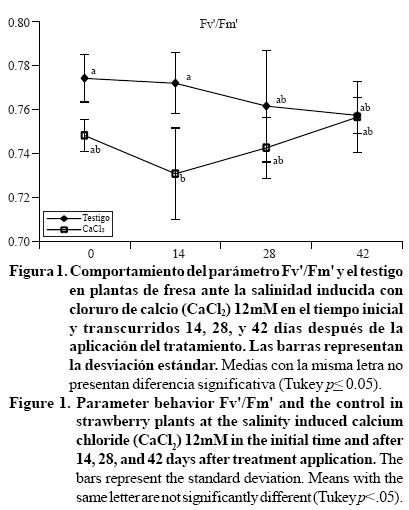
La aplicación de CaCl2 no afectó significativamente a la mayoría de los parámetros de fluorescencia. No obstante el parámetro Fv'/Fm' mostró afectación al inicio de la aplicación y a los 14 días, posterior a la cual se observó una estabilización (Figura 1). Este parámetro provee un estimado de la eficiencia máxima de la fotoquímica del fotosistema II a una densidad dada de flujo fotónico de la radiación PAR, el cual representa la eficiencia operativa del fotosistema II si todos los centros correspondiente fueron abiertos (quinona Q oxidada) (Baker, 2008). Ésta respuesta puede deberse a que la salinidad produjo la disminución en la eficiencia del fotosistema II, pero sin activar su mecanismo de disipación energética, observado en la no afectación del parámetro (NPQ) y la no disminución en la eficiencia operativa del fotosistema II (ePS2) (Tabla 1). Esto sugiere que la disminución de la eficiencia máxima del fotosistema II, observado a partir del día 28, es atribuible a la restauración en la habilidad de los centros de reacción para realizar el transporte de electrones. Sobre esto último, se tienen reportes de que el parámetro NPQ es el primer parámetro afectado en los tratamientos salinos (Han et al., 2010), situación no observada en este experimento, donde la dosis de 12 mM de CaCl2 sólo afectó al mínimo la eficiencia fotoquímica.
Las plantas de fresa mostraron una disminución paulatina en la eficiencia fotoquímica al aumentar la temperatura de 19 a 34 °C, siendo más acentuado el efecto a partir de los 28 °C. Esta reducción pudiera ser resultado de la caída en la eficiencia cuántica máxima (Fv/Fm), debida principalmente a la disminución en la fluorescencia máxima (Fm) (Figura 2). Al mismo tiempo, no hubo afectación en los parámetros referentes a la fluorescencia mínima (Fo), rendimiento cuántico efectivo (ePS2) ni en los decaimientos de la fluorescencia debidas a procesos fotoquímicos (qP) y no fotoquímicos (NPQ), lo que puede indicar que en las temperaturas de 19 a 34 °C no se presentó estrés oxidativo, que es la respuesta más común de las plantas frente a temperaturas extremas (Ogweno et al., 2009). A pesar de ello, se observó que el parámetro de estabilidad (Fs) disminuyó, lo que pudiera indicar una disminución en la capacidad de disipación del gradiente de protones (ΔpH) entre el lúmen del tilacoide y el estroma (Rohacek, 2002). Éstos resultados indican un declive en la eficiencia fotoquímica de las plantas por encima de los 28 °C, mientras que por debajo de los 19 °C mostró una mayor eficiencia.
La máxima eficiencia cuántica (Fv/Fm) se encontró por debajo de los 19 °C, disminuyendo al aumentar la temperatura y mostrando un fuerte declive con temperaturas mayores a 28 °C. Sin embargo, el parámetro de estabilidad (Fs) fue reducida a temperaturas superiores a los 19 °C lo cual indica efectos adversos en la síntesis de moléculas NADPH y ATP para la fotosíntesis. Los parámetros qP y NPQ indicaron que no se presentó estrés oxidativo en el espectro de temperatura estudiado.
Literatura citada
Barroso, M. C. y Alvarez, C. E. 1997. Toxicity symptoms and tolerance of strawberry to salinity in the irrigation water. Sci. Hort. 71(3):177-188. [ Links ]
Baker, N. R. 2008. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 59(1):89-103. [ Links ]
Bisko, A., Cosió, T., y Jelaska, S. 2010. Reaction of Three Strawberry Cultivars to the Salinity: Vegetative Parameters. Agriculturae Conspectus Scientificus (ACS). 75(2):83-90. [ Links ]
Casierra-Posada, F. y García-Riaño, N. 2006. Producción y calidad de fruta en cultivares de fresa (Fragaria sp.) afectados por estrés salino. Rev. Fac. de Agro. Medllin. 59(2):3527-3542. [ Links ]
Jaramillo-Giraldo, M. A.; Oliva-Cano y Ferreira-Ribas, R. 2009. Respuesta fotosintética de diferentes ecotipos de frijol a la radiación y la salinidad. Corpoica Colombiana de Investigación Agropecuaria. 10(2):129-140. [ Links ]
Kadir, S. and Sidhu, G. 2006. Strawberry (Fragaria xananassa Duch.) growth and productivity as affected by temperature. Hortscience. 41(6):1423-1430. [ Links ]
Han, W.; Xu, X. W.; Li. L.; Lei, J. Q. and Li, S. Y. 2010. Chlorophyll a fluorescence responses of Haloxylon ammodendron seedlings subjected to progressive saline stress in the Tarim desert highway ecological shelterbelt. Photosynthetica. 48(4):635-640. [ Links ]
Hughes, N. M. and Smith, W. K. 2007. Seasonal photosynthesis and anthocyanin production in 10 broadleaf evergreen species. Funct. Plant Biol. 34(12):1072-1079. [ Links ]
Lanauskas, J.; Uselis, N.; Valiuskaite, A. y Viskelis, P. 2006. Effect of foliar and soil applied fertilizers on strawberry healthiness, yield and berry quality. Agron. Res.4:247-250. [ Links ]
Ogweno, J. O.; Zhou, Y. H. and Yu, J. Q. 2009. Changes in activities of antioxidant enzymes and photosynthesis in detached leaves of tomato after exposure to different temperatures. Afr. J. Hort. Sci. 2(1):124-137. [ Links ]
Rohacek, K. 2002. Chlorophyll fluorescence parameters: the definitions, photosynthetic meaning, and mutual relationship. Photosynthetica. 40(1):13-29. [ Links ]
Steiner A. A. 1961. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil. 15(2): 134-154. [ Links ]
Xu, Z.; Zhou, Z. G.; Han, G. and Li, Y. 2011. Photosynthetic potential and its association with lipid peroxidation in response to high temperature at different leaf ages in maize. J. Plant Growth Regul. 30(1):41-50. [ Links ]

