










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.3 no.1 Texcoco ene./feb. 2012
Artículos
Adaptaciones morfológicas foliares en tomate cultivado en bioespacio de techo retráctil con clima cálido*
Morphological leaf adaptations in tomato grown in warm retractable-roof bio-space
Jorge Berni Medina Medina1, José de Jesús Luna Ruíz1§, Joaquín Sosa Ramírez1, Onésimo Moreno Rico2 y Catarino Perales Segovia3
1 Centro de Ciencias Agropecuarias. Universidad Autónoma de Aguascalientes. Av. Universidad 940. Ciudad Universitaria, Aguascalientes, México. C. P. 20131. Tel. 01 449 1393248. Ext. 8107 (jberni@bernilabs.com), (jsosar@correo.uaa.mx). §Autor para correspondencia: jjluna@correo.uaa.mx.
2 Centro de Ciencias Básicas. Universidad Autónoma de Aguascalientes. Tel. 01 449 9107400. Ext. 8118. (omoreno@correo.uaan.mx).
3 Instituto Tecnológico El Llano Aguascalientes. Carretera Aguascalientes-San Luis Potosí, km 18. Aguascalientes, México. Tel. 01 449 9161251 (cperaless@hotmail.com).
*Recibido: mayo de 2011
Aceptado: noviembre de 2011
Resumen
Se analizaron las condiciones microclimáticas y la morfologíafoliar de tomate indeterminado de dos bioespacios: invernadero de techo retractil y casa sombra en Culiacán, Sinaloa, México; durante el periodo 2009 a 2010. Los resultados muestran que el invernadero de techo retractil, genera condiciones más favorables para la fotosíntesis, como resultado de una menor fluctuación térmica al interior de este bioespacio, en comparación con el ambiente impuesto por la casa sombra. Las temperaturas extremas (máximas y mínimas) al interior del invernadero de techo retractil, se mantuvieron en un rango significativamente menor y por lo tanto, más favorable para la producción y la estabilidad reproductiva de tomate en el clima cálido de Culiacán, en comparación con casa sombra. Lo anterior fue corroborado por la magnitud de las variables morfológicas foliares analizadas y las diferencias detectadas entre foliolos desarrollados en ambos casos. Los valores de índice de área foliar, grosor de hoja, longitud del parénquima en empalizada y presencia de doble parénquima empalizada, observados en plantas desarrolladas en invernadero de techo retractil, superaron significativamente a los valores observados en casa sombra. Los resultados permiten concluir que los mayores rendimientos de tomate reportados recientemente para invernadero de techo retractil en climas cálidos, se deben en gran parte a la mayor eficiencia fotosintética, que resulta de las modificaciones y adaptaciones morfológicas foliares al microclima impuesto por el invernadero de techo retractil. Estos resultados apoyan la visión del manejo agronómico innovador, para la búsqueda por acondicionarlos espacios para la expresión de la vida. El concepto científico de bioespacio para la producción vegetal bajo sistemas de agricultura protegida, es fundamental para proponer medidas de adaptación ante el cambio climático.
Palabras clave: Solanum lycopersicim, casa-sombra, invernadero-sustentable, SAS.
Abstract
Microclimatic conditions and indeterminate tomato leaf morphology of two bio-spaces were analyzed: retractable-roof greenhouse and shade-house in Culiacán, Sinaloa, Mexico, during the period 2009 - 2010. The results show that, the retractable-roof greenhouse generates more favorable conditions forphotosynthesis, as a re sult of lower thermal fluctuation within this bio-space, compared with the environment imposed by the shade-house. Extreme temperatures (maximum and minimum) into the retractable-roof greenhouse remained in a range significantly smaller and therefore more favorable for the production and reproductive stability of tomato in the warm weather of Culiacán, comparedto shade-house s. This was corroborated by the magnitude of leaf morphological variables analyzed and the differences detected between leaflets developed in both cases. The values of leaf area index, leaf thickness, length of palisade parenchyma and presence of double palisade parenchyma observed in plants grown under retractable-roof made of glass were significantly higher than the values observed in the shade-house. The results suggested that higher tomato yields recently reported retractable-roof greenhouse in warm climates are due in large part to the higher photosynthetic efficiency, resulting from the amendments and foliar morphological adaptations imposed by the retractable-roof greenhouse' s microclimate. These results support the vision of innovative agricultural management to put the search space for the expression of life. The scientific concept of bio-space for crop production under protected agriculture systems is essential to propose adaptation measures to climate change.
Key words: Solanum ly copersicum, shade-house, sustainable glasshouse, SAS.
Introducción
La producción sostenible de tomate en sistemas de agricultura protegida, requiere de tecnología y conocimientos apropiados para lograr la máxima productividad, calidad y aprovechamiento racional de los recursos disponibles en el agroecosistema. Los esfuerzos tecnológicos se han encaminado preponderantemente al mejoramiento genético de variedades para condiciones de invernadero, a optimizar técnicas de nutrición y fertirriego, al uso de agroquímicos "de avanzada" (más amigables con el ambiente), a la incorporación de buenas prácticas de producción y poscosecha tales como el manejo integrado del cultivo y de poscosecha. El concepto de bioespacio en agricultura protegida, se entiende como aquella estructura construida por el hombre, para favorecer el desarrollo de plantas cultivadas en condiciones ambientales semirreguladas de humedad relativa, radiación y temperatura (Bustamante et al., 1999).
Bajo un enfoque sistémico, el bioespacio tiene como objetivo alinear la estructura foliar con otros sistemas celulares y variables ambientales, de tal manera que cada hoja funcione como una interface física para el flujo de energía solar y el optimo funcionamiento del sistema fotosintético de la planta. En un bioespacio las células del parénquima en empalizada y las cámaras estomáticas operan eficientemente para capturar energía lumínica y CO2, y mediante la evapotranspiración se favorece la disipación de calor y el movimiento de nutrientes (Laborit, 1973; Nobel et al., 1975; Boardman, 1977; Björkman, 1981; Anderson, 1986; Bakery McKiernan, 1988; Cui et al., 1991; Norman et al., 1996). Un buen sistema foliar permite: 1) la captura de la luz fotosintéticamente activa (PAR); 2) la captura de calor durante la temporada fría; y 3) la eliminación de calor excesivo durante la estación caliente (Hare et al., 1984; Cui et al., 1991; Critten y Bailey, 2002; Taiz y Zieger, 2002; Kumar et al., 2009).
Con la evolución tecnológica en la agricultura protegida, se ha logrado eludir gran parte de la variación climática y algunos eventos meteorológicos adversos como lluvias torrenciales, granizo, vientos, temperaturas extremas, etc. El uso de estructuras estáticas de soporte y protección, materiales plásticos de cubierta y sistemas de enfriamiento y calentamiento, forman parte de la gama de bioespacios para la producción vegetal (Arellano et al., 2006; Pascale y Marcellis, 2009). En invernaderos para latitudes mayores a 37° de latitud norte, se han alcanzado niveles de productividad sobresalientes. Por ejemplo en Almería, España bajo casa sombra la producción de tomate es de 18 a 20 kg m2, y en Holanda la producción bajo invernadero es de 58 a 60 kg m2.
El manejo tecnológico en estas regiones se ha enfocado a variables como luminosidad, flujo forzado de carbono y fertirrigación (Verhaegh y De Groot, 2000). Transferir y adaptar el conocimiento ecofisiológico puede beneficiar a los productores de otras regiones, quienes siempre están buscando cómo lograr la máxima producción y cómo penetrar las ventanas de oportunidad del mercado, para colocar su producción de manera rentable y competitiva (Baille, 2001; Cook y Calvin, 2005). A principios de los 90's se generaron diversos diseños de invernaderos con movilidad de techos y paredes, destacando el invernadero de techo retráctil (ITR).
El ITR surge de una búsqueda, para favorecer el desarrollo y producción de plantas cultivadas en condiciones semicontroladas de humedad relativa, radiación y temperatura. El ITR, considerado un modelo de producción de agricultura protegida, supero en rendimiento la producción de tomate en casa sombra (CS) e invernadero para climas templados y fríos (Schuch, 2003; Giacomelli y Suárez, 2004; Rogoyski, 2004; Vollebregt, 2004; Bartok, 2005; Mathers et al., 2007; Roberts et al., 2010; Vollebregt, 2010). Las experiencias con tomate en el Valle de Culiacán, México; durante los ciclos agrícolas otoño-invierno 2007-2008 y otoño-invierno 2008-2009, indican que los rendimientos en ITR fluctúan entre 13.8 y 16.3 kg m2 y de 6.4 a 7.9 kg m2 en CS (Figura 1).

A pesar de las ventajas en rendimiento y producción observadas con el ITR, el proceso de asimilación de esta innovación tecnológica ha sido lento en sus diferentes etapas -burla-ataque-aceptación (Kuhn, 1996). Lo anterior posiblemente se debe a la falta de fundamentos científicos, que ayuden a explicar los incrementos en rendimiento detectados en ITR. Los mayores rendimientos observados con tomate en ITR, podrían ser explicados por modificaciones en la morfología y anatomía foliar, bajo la influencia del microclima impuesto por el ITR, principalmente por la influencia de menores oscilaciones térmicas con respecto a otros bioespacios y al ambiente exterior.
Las modificaciones morfológicas y anatómicas en tejido foliar en respuesta al ambiente tienen relación directa con eficiencia fotosintética y por lo tanto, con los procesos de crecimiento, desarrollo, reproducción y rendimiento. El ambiente operacional del ITR está centrado en el funcionamiento optimo de la planta; el ITR opera mediante techos y paredes retractiles, que actúan como una máquina-herramienta que establecen ciclos microclimáticos de retroalimentación positiva y restrictiva (Salisbury y Ross, 1992; Connor y McDermott, 1998), mediante el cierre y apertura de techos y paredes durante lapsos cortos de tiempo, programables en minutos, de tal forma que es posible dispersar la luz, reducir las cargas térmicas, mejorar las condiciones de crecimiento y desarrollo, regular la condición y estructura del dosel, así como del conjunto de plantas bajo cultivo, que conduce a una mejor consistencia de hojas y frutos (Zhang et al. , 201 0), y una mejora de vida de anaquel (Roberts, 2000; Mathers et al., 2007; Roberts et al., 2010).
Por otra parte, el ambiente operacional del bioespacio CS; no obstante, que parece no restringir la fotosíntesis, se asocia a un elevado crecimiento vegetativo con una aparente restricción durante la etapa de formación de frutos y crecimiento de los mismos (Smith et al., 1984; Bustamante et al., 1999; Zhang et al., 2010). El carbono es el principal elemento constitutivo de la planta, representa 45% del peso seco (Salisbury y Ross, 1992), por lo que es esencial en plantas C4 mantener elevada la concentración y la disponibilidad de CO2, tanto en la cámara estomática como en el cloroplasto; de ahí que la adaptabilidad y plasticidad de las estructuras morfológicas foliares del mesófilo, involucradas en la penetración de la luz y en la asimilación del CO2, representan por su arreglo arquitectónico, formas y espacios críticos al interior de la hoja, para maximizar el proceso fotosintético en ambientes de agricultura protegida.
En agricultura protegida la eficiencia en el manejo de los insumos para la planta, no solo se deriva del flujo coheso-tenso-transpiratorio del agua y nutrientes provenientes del suelo ó sustrato, sino que también de la eficiencia con que se secuestre y se fije el carbono del aire, a través del propio arreglo y disposición estructural de las células al interior de la cámara estomática (Spomer, 1973; Stanhill, 1979; Hart, 1982; Nobel, 1983; Strohman, 1997; Philip, 2004; Terashima et al., 2006). Evaluar los atributos del ITR y su impacto en el desarrollo y producción vegetal, contribuye a definir el contenido innovador de este modelo tecnológico ante la búsqueda de sistemas sustentables de agricultura protegida, altamente productivos y de bajo consumo energético en regiones cálidas (Hare et al., 1984; Baille, 2001).
De ahí que los objetivos de este trabajo se centraron en: 1) determinar si los incrementos en rendimiento detectados a favor del ITR en el Valle de Culiacán, Sinaloa; pueden ser explicados por cambios favorables en el microclima al interior de cada bioespacio; y 2) determinar si las características morfológicas foliares: índice de área foliar (IAF), grosor de hoja (GH), longitud del parénquima empalizada (LPE) y doble parénquima empalizada (DPE) de plantas producidas bajo ITR, difieren favorablemente con respecto a CS. Las posible diferencias microclimaticas y en morfología foliar, podrían ayudar a explicar las diferencias en rendimiento de tomate reportadas recientemente a favor del ITR en el Valle de Culiacán.
Materiales y métodos
Variables microclimáticas. Para el primer objetivo se identificaron y evaluaron las condiciones de microclima, específicamente temperatura máxima (Tmax) y temperatura mínima (Tmin) al interior del ITR y CS para el periodo 2009 a 2010. Los registros del interior de cada bioespacio, fueron comparados con los registros del exterior para el mismo periodo y las diferencias medias (Di) del ITR, fueron contrastadas con las de CS mediante pruebas basadas en t de student para diferencias. Los datos diarios de Tmax y Tmin del interior de cada bioespacio, fueron registrados y almacenados por sensores automáticos, en tanto que los datos de Tmin y Tmax del exterior para el mismo periodo, fueron extraídos de la base de datos diarios de la estación meteorológica automática del INIFAP más cercana a cada bioespacio. Para el ITRlos datos microclimaticos provienen de la estación localizada en el Campo Experimental Culiacán (24° 38' 02.44" latitud norte y 107° 26'19.21" longitud oeste), y para CS los datos provienen del Campo La Primavera (24°29'54.82" latitud norte y 107°27'49.75" longitud oeste).
Variables morfológicas foliares. Para el segundo objetivo se analizaron variables de morfología foliar, provenientes de muestras de plantas de tomate indeterminado variedad Moctezuma, cultivadas en ITR y CS, ambos con manejo convencional. Las determinaciones de morfología foliar se realizaron con microscopio óptico y electrónico de barrido (JMS-35®) en el laboratorio de Biología, Departamento de Biología, Centro de Ciencias Básicas de la UAA. Para ello se colectaron hojas completamente desarrolladas tanto en ITR como en CS a los 160 días de trasplante. Cada planta muestreada se dividió en tres secciones: superior, media e inferior. En cada sección los foliolos fueron desprendidos del tallo principal y colocados en bolsas de plástico, etiquetadas, empacadas y almacenadas en un cuarto frío.
Índice de área foliar (IAF). Se obtuvo por diferencia entre la superficie cubierta por los foliolos de cada sección, en una superficie conocida de papel milimétrico de 3 000 cm2. El área foliar por planta (AFP) fue obtenida sumando las áreas foliares de las tres secciones de cada planta. El área foliar por m2 (AFM) se obtuvo multiplicando AFP por la cantidad de plantas en 1 m2 de bioespacio (tres plantas). El IAF equivale al valor de AFM expresado en m2.
Grosor de hoja (GH). Esta variable se obtuvo de dos maneras: 1) mediante la preparación de cortes transversales de 10 hojas frescas de cada sección, preservadas en formol-alcohol-acido acético (FAA), con montaje en para fina y corte con micrótomo tipo rotatorio; cada corte fue inspeccionado al microscopio y fotografiado a 20x; las mediciones de GH se realizaron con un software especializado para manejo y caracterización de tomas microscópicas; 2) mediante preparaciones derivadas de cortes transversales de 10 hojas frescas de cada sección preservadas en oro y observadas en dos campos con microcopia electrónica de barrido a 120x, 200x y 350x; las mediciones de GH en cada campo fueron registradas en micras desde el borde superior del corte de hoja (lamina) hasta el borde inferior (envés), ambos extremos delimitados por la cutícula.
Longitud del parénquima en empalizada (LPE). Se realizaron montajes en parafina a partir de muestras de tejido foliar de las tres secciones de la planta y estas fueron fijadas en FAA; de cada montaje se realizaron tres cortes transversales, y de cada corte se obtuvieron diez muestras con microtomo. Las muestras se procesaron en histoquinet T/P 8 000 durante 12 h, para después fijarlas en bloques de parafina. Los cortes histológicos se realizaron en un micrótomo rotatorio LEICA RM2125RT a un grosor de 10 micras; los tejidos de cada corte con micrótomo se colorearon con la técnica de Hematoxilina y Eosina (HE) (Curtis, 1968) y se fotografiaron; la medición de LPE fue realizada con el software Image-Pro Plus a 20x. La LPE también fue estimada con un microscopio electrónico de barrido modelo JFOL JSM-35C (JEOLLTD, Tokio, Japón) en la Universidad Autónoma de Aguascalientes. Para ello se prepararon cortes transversales de hoja procedentes de cada bioespacio y se observaron en una secuencia de resolución a 120x, 200x y 350x.
Doble parénquima en empalizada (DPE). Esta variable se obtuvo a partir de los cortes histológicos, realizados para estimar LPE en las tres secciones de las plantas muestreadas en ITR y CS (superior, media, inferior). Se registró la frecuencia de campos con DPE mediante observaciones al microscopio óptico a 20X. Los datos se organizaron por sección para determinar la frecuencia de muestras con DPE en cada sección de la planta. Los cortes histológicos que mostraron al menos tres campos con DPE, fueron considerados positivos para esta variable.
Análisis estadístico. Los datos de variables microclimaticas y de morfología foliar procedentes del ITR y CS, fueron organizados en diferentes bases de datos y analizados con el programa STATISTICA Ver. 8.0 (Stat Soft Inc. 1984-2007).
Resultados y discusión
Variables microclimáticas
Los resultados del análisis microclimatico se muestran en los cuadros 1 y 2. El análisis de temperaturas extremas indica que las condiciones al interior del ITR, fueron más estables que en CS a pesar que la condición de temperaturas externas del ITR fueron más extremosas que en CS.

Temperatura máxima (Tmax). Los resultados para Tmax se presentan en el Cuadro 1, el cual muestra los valores medios de Tmax registrados al interior y exterior del ITR y CS. También se presentan las diferencias medias observadas para Tmax al interior y exterior de cada bioespacio y la comparación para saber si hay diferencia significativa entre ambas diferencias, entre la temperatura al interior del bioespacio con respecto al exterior, no es igual para ITR y CS. En este sentido, la reducción de temperatura en el ITR fue de 8.9 °C (47.6°C a 38.7 °C) equivalente a 18.6% con respecto a la temperatura exterior. La CS redujo la temperatura en 2.1 °C (de 40.3 °C a 38.2 °C), equivalente al 5.1% respecto a la temperatura exterior. Los cambios en temperatura impuestos por el ITR, son significativamente diferentes a los cambios impuestos por la CS al interior de cada bioespacio (Cuadro 1).
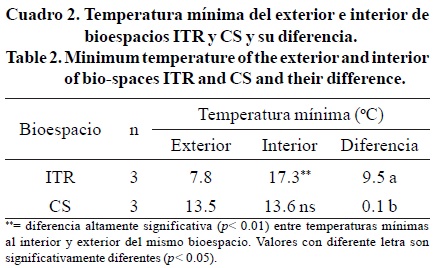
Temperatura mínima (Tmin). Los valores de Tmin exterior e interior para ambos bioespacios y sus diferencias se presentan en el Cuadro 2. La interacción del sistema helio-arquitectónico de cada bioespacio con el clima circundante, provoca diferencias de temperatura al interior de cada bioespacio en relación con la condición temperatura del exterior (Arellano et al., 2006). Al interior del bioespacio ITR se incrementó de manera favorable 9.5 °C la Tmin a promediar 17.3 °C al interior, contra 7.8 °C del exterior, diferencia equivalente a 54.9%. Esta diferencia no ocurrió en el bioespacio CS donde la Tmin del interior (13.6 °C), fue prácticamente igual que en el exterior (13.5 °C).
Variables morfológicas foliares
Los resultados del análisis de la morfología foliar se muestran del Cuadros 3 (4, 5) al Cuadro 6; Figuras 2 y Figura 3. Para cada variable se presentan las diferencias observadas entre ITR y CS y la significancia estadística.





Índice de área foliar (IAF). En el Cuadro 3 se puede observar que a 15 8 días del trasplante, el IAF en el bioespacio ITR presenta diferencia significativa superior 61%, contra el bioespacio CS. Considerando que esta diferencia en el IAF, se encuentra en una etapa de su ciclo fenológico con menor elasticidad de tiempo biológico, debido al desgaste y envejecimiento de tejidos impuestos por las condiciones microambientales del bioespacio en CS (Bradford y Trewavas, 1994; Hattrup et al., 2007).
Generalmente es aceptado que la cantidad de sustancias orgánicas que una plantación produce, está en función de: 1) la superficie de la hoja; 2) la estructura y desarrollo fotosintético, entre lo que estructuralmente destaca la conformación del parénquima en empalizada; y 3) índice de área foliar (IAF), es decir del área de los limbos foliares sobre la unidad de superficie del suelo (Fogg, 1967; Gómez et al., 1999); esto resulta particularmente importante en el cultivo de tomate, en variedades de crecimiento indeterminado y especialmente en agricultura bajo cubierta, ya que el manejo agronómico de las diferentes etapas de crecimiento, desarrollo y fructificación, se manipula en base a una serie de podas sistemáticas que se realizan, conforme el crecimiento del tallo y la estructura foliar que en su conjunto avanza y se va soportando sobre líneas cargadoras de la biomasa de la planta en su conjunto: tallo, foliolos y frutos en diferentes etapas.
El IAF en el bioespacio CS mostró valores inferiores respecto del ITR, debido probablemente a las bajas intensidades luminosas características de este bioespacio, y en parte también porque las hojas inferiores entran en senescencia más rápido e inactivan en consecuencia, su capacidad fotosintética (Cayón, 1992).
Grosor de hoja (GH). En las hojas del bioespacio ITR, las mediciones realizadas mostraron 28% de mayor grosor comparativamente contra el bioespacio CS. El comportamiento registrado en esta variable resultó de gran importancia para explicar las diferencias en producción de los dos bioespacios estudiados, ya que las hojas del tercio superior del bioespacio ITR fueron 77.7% más gruesas que las hojas del mismo tercio en el bioespacio CS). Esta diferencia significativa de GH en el tercio superior de las plantas de tomate del bioespacio ITR (Cuadro 4), concuerda con los estudios realizados en espina capor Cui et al. (1991) y por Taiz y Zeiger (2002), donde se confirma que el grosor y arreglo de la estructura interna de la hoja, crea una condición que optimiza el aprovechamiento de: 1) la radiación fotosintéticamente activa (PAR); y 2) de la difusión de CO2 al interior de la hoja a través de cámaras estomáticas mejor conformadas (Salisbury y Ross, 1992).
Lo anterior es una respuesta estructural adaptativa ante una mayor exposición a la radiación solar directa y que, como lo indica Müller et al. (2001), al ser la luz el parámetro ambiental desde el que las plantas han evolucionado en numerosos caminos bioquímicos y en respuestas morfológicas adaptativas alas intensidades y calidades lumínicas, se optimiza el proceso fotosintético y se incrementa el rendimiento.
Longitud de parénquima en empalizada (LPE).
Las estructuras de parénquima en empalizada, que se desarrollaron tanto en el bioespacio ITR, como en CS, corresponden con diferentes adaptaciones morfológicas, en los ambientes: regulado flexible de sol y sombra (ITR) y de sombra persistente (CS) descritas anteriormente, las cuales en el bioespacio ITR muestran una longitud promedio del parénquima en empalizada de 129 micras, considerando los tres tercios del follaje evaluados, la cual es ligeramente superior a la región de máximo aprovechamiento de la luz ubicada entre las 100 a 120 micras, para este grupo de células especializadas del parénquima en empalizada. El promedio de LPE del bioespacio de CS fue de 100.3 micras para los tres tercios del follaje evaluados (Figura 2 y Cuadro 5).
Los resultados del análisis morfológico confirman que las células del parénquima en empalizada del tomate corresponden a plantas C3. Las células del parénquima se organizan perpendicularmente al ángulo de incidencia de la luz. La mayor elongación y alta concentración de cloroplastos en este tejido son cruciales para la fotosíntesis; plantas y hojas con estas características pueden alcanzar hasta 90% de eficiencia fotosintética, si la radiación es captada por cloroplastos funcionales localizados a unos 100-120 micras de la cutícula foliar (Nobel et al., 1975; Boardman, 1977; Björkman, 1981; Anderson, 1986; Baker y McKiernan, 1988 citados por Cui et al., 1991).
Las condiciones de sol y sombra impuestas por el ITR y de sombra persistente en CS, conducen al desarrollo de diferentes adaptaciones morfológicas foliares, y en consecuencia a diferentes LPE (Cui et al., 1991; Stern, 1994; Taiz y Zeiger, 2002). Asimismo, las plantas expuestas a la luz directa, desarrollan hojas más gruesas y con superficies foliares mayores al interior del mesofilo esponjoso (Figura 2). Estas adaptaciones morfológicas foliares mejoran substancialmente la conductancia estomatal e intercambio gaseoso (CO2, O2 y vapor de agua) durante los procesos de fotosíntesis, transpiración y respiración.
Lo anterior conlleva un incremento en la tasa de fijación de carbono, sobre todo cuando se considera que esta superficie interna de exposición al intercambio gaseoso es de 10 a 40 veces mayor que la superficie externa de la hoja (Nobel, 1983), lo cual afecta positivamente la captura de CO2, y por consecuencia directa la generación de fotoasimilados y acumulación de biomasa. Una alta frecuencia de hojas provenientes de CS también mostraron el típico "síndrome de hoja de papel" (shade avoidance síndrome, SAS) descritas por Whitelam et al. (2004); Hattrup et al. (2007). Las hojas con SAS provenientes de CS presentaron gran fragilidad estructural, durante su manipulación y preparación de muestras para microscopia. La fragilidad detectada durante el manejo de foliolos provenientes de CS, coincidió con los datos morfológicos y las microfotografías provenientes del análisis.
Doble parénquima en empalizada (DPE). Los resultados para esta variable se muestran en el Cuadro 6, Cuadro 7 y Figura 3. De los 84 campos microscópicos observados en hojas provenientes de ITR, 44 (53%) mostraron DPE. Esta característica no fue observada en CS, ya que ninguno de los 87 campos microscópicos observados en foliolos de este bioespacio mostro DPE. La presencia de DPE en ITR apareció en todos los 25 campos observados de la sección superior de la planta y en 19 de los 30 campos de la sección media. Ninguno de los 29 campos inspeccionados de la sección inferior en ITR mostro DPE.

Estos resultaos indican que las condiciones microclimaticas del ITR promueven el desarrollo de DPE en las hojas medias y superiores de la planta. La presencia de DPE es indicativa de mayores tasas y eficiencia fotosintética (Salisbury and Ross, 1992; Taiz and Zeiger, 2002).
Conclusiones
Los resultados del presente estudio, permiten concluir que los mayores rendimientos de tomate reportados recientemente para ITR en climas cálidos, se deben en gran parte a la mayor eficiencia fotosintética, que resulta de las modificaciones y adaptaciones morfológicas foliares al microclima más favorable que impone el ITR. Estos resultados apoyan la visión del manejo agronómico innovador, para la búsqueda por acondicionar los espacios para la mejor expresión de la vida. El concepto científico de bioespacio para la producción vegetal bajo sistemas de agricultura protegida, es fundamental para proponer medidas de adaptación ante el cambio climático.
Literatura citada
Anderson, J. M. 1986. Photoregulation of the composition, function and structure of thylakoid membranes. Annu. Rev. Plant Physiol. 37:93-136. [ Links ]
Arellano, G. M.; Valera, M. D.; Urrestarazu, G. M.; García, G. S.; Sánchez, S. S. y Soria, R. J. 2006. Estudio microclimático en dos subtipos de invernaderos almeria. Agric. Téc. Méx. 32(2):225-234. [ Links ]
Baille, A. 2001. Trends in greenhouse technology for improved climate control in mild winter climates. ISHS. ActaHortic. 559:161-168. [ Links ]
Baker, N. R. and Mckiernan, M. 1988. Modifications of the photosynthetic apparatus of higher plants in response to changes in the light environment. Biol. J. Linnean Soc. 34:203. [ Links ]
Bartok, J. W. 2005. Retractable roof greenhouse and shadehouses. USDA Forest Service Proceedings. U. S. Department of Agriculture. 73-75 pp. [ Links ]
Björkman, O. 1981. Responses to different quantum flux densities. Encyclopedia of plant physiology. (Ed. Lange, O. L.; Nobel, P. S. and Ziegler, H.). New Series, Springer-Verlan, Berlin. 12.57-106. [ Links ]
Boardman,N. K. 1977. Comparative photosynthesis of sun and shade plants. Plant Physiol. 28:355-377. [ Links ]
Bradford, K. J. and Trewavas, A. J. 1994. Sensitivity thre sholds and variable time scales in plant hormone action. Plant Physiol. 105:1029-1036. [ Links ]
Bustamante, O. J.; Gonzáles, H. V.; Livera, M. M. y Zavaleta, M. E. 1999. Cambios fisiológicos y microclimáticos inducidos en jitomate por una cubierta flotante. Agrociencia. 33:31-39. [ Links ]
Cayón, G. 1992. Fotosíntesis y productividad de cultivos. Revista Comalfi. 19:23-31. [ Links ]
Cheema, D. S.; Kaur, P. and Kaur S. 2004. Off-season cultivation of tomato under net house conditions. ISHS. ActaHortic. 659:177-181. [ Links ]
Cook, R. and Calvin, L. 2005. Greenhouse tomatoes change the dynamics of the North American fresh tomato industry. USDA United States Department of Agriculture. Economic Research Report Number 2.68p. [ Links ]
Connor, J. y McDermott, I. 1998. Introducción al pensamiento sistémico. Recursos esenciales parala creatividad y la resolución de problemas. Ediciones Urbano. Argentina. 302 p. [ Links ]
Critten, D. L. and Bailey, B. J. 2002. Are view of greenhouse engineering developments during the 1990's. Elsevier. Agric. Forest Meteorol. 112:1-22 pp. [ Links ]
Cui, M.; Vogelmann, T. C. and Smith, W. K. 1991. Chlorophyll and light gradients in sun and shade leaves of Spinacia oleracea. Plant Cell Environ. 14:493-500. [ Links ]
Curtis, P. J. 1968. Manual de técnicas histológicas y citológicas. Escuela Nacional de Agricultura. Chapingo, Estado de México. 94 p. [ Links ]
Fogg, G. E. 1967. El crecimiento de las plantas. Editorial Universitaria de Buenos Aires. 327 p. [ Links ]
Giacomelli, G. and Suárez, R. A. 2004. Environmental and plant growth experiences in a retractable roof greenhouse under semi-arid consitions. ISHS International Symposium on Protected Culture in a Mild-Winter Climate. FL, USA. 23-27 pp. [ Links ]
Gómez, C. C.; Buitrago, M.; Cante y Huertas, B. 1999. Ecofisiología de la papa (Solanum tuberosum) utilizada para consumo fresco y para la industria. Revista Comalfi. 26:42-55. [ Links ]
Hare, J. G.; Norton, B. and Probert, S. D. 1984. Design of greenhouses: thermal aspects. Appl. Energy. 18:49-82. [ Links ]
Hart, D. R. 1982. Agroecosistemas, conceptos básicos. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, CostaRica. 2-50pp. [ Links ]
Hattrup, E.;Neilson, A. K.; Breci, L. andHaynes,A. P. 2007. Proteomic analiysis of shade-avoidance response in tomato leave. Agric. FoodChem. 55:8310-8318. [ Links ]
Kuhn, T. S. 1996. The structure of scientific revolutions. 3th edition. University of Chicago Press. USA. [ Links ]
Kumar, K. S.; Tiwari, K. N. and Madan, K. J. 2009. Design and technology for greenhouse cooling in tropical and subtropical regions: a review. Energy and Buildinds. 41:1269-1275. [ Links ]
Laborit, H. 1973. Del sol al hombre. 4ta edición. Editorial Labor S.A. Barcelona. 154 p. [ Links ]
Mathers, H.; Case, L. and Rivera, D. 2007. An ace up your sleeve. American Nurseryman. 36-46 pp. [ Links ]
Müller, P.; Xiao, P. L. and Niyogi, K. K. 2001. Non-photochemical quenching a response to excess light energy. Plant Physiol. 125:1558-1566. [ Links ]
Nobel, P. S. 1983. Biophysical Plant Physiol. Ecol. Freeman, USA. 608 p. [ Links ]
Nobel, P. S.; Zaragosa, L. J. and Smith, W. K. 1975. Relationship between mesophyll surface area, photosynthetic rate, and illumination level during development of leaves of Plectranthus parviflorus. Henekel. Plant Physiol. 55:1067-1070. [ Links ]
Norman, W. D.; Worman, F. D.; Siebert, J. D. y Modiakgotla, E. 1996. El enfoque de sistemas agropecuarios para el desarrollo y la generación de tecnología apropiada. FAO. Núm. 10. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Roma. 51-94 pp. [ Links ]
Pascale, S. D. and Marcellis, M. F. 2009. Crop management in greenhouses: adapting the growth conditions to the plant needs or adapting the plant to the growth conditions? Acta Hortic. 807:163-173. [ Links ]
Philip, N. 2004. Física biológica, energía, información y vida. Reverte. México. 631 p. [ Links ]
Roberts, J.W. 2000. Greenhouse technology -open roof design. Rutgers University, New Brunswick, ASP. USA. [ Links ]
Roberts, J. W.; Mears, D. R.; Reiss, E. and Both, A. J. 2010. Open-roof greenhouse design and operation, bioresource engineering. Department of Plant Biology and Pathology, Rutgers University. 12 p. [ Links ]
Rogoyski, M. 2004. Production of container-grown cliff rose plants in three environments using several irrigation methods. 5 p. [ Links ]
Salisbury, B. F. and Ross, C. W. 1992. 4th edition. Wadsworth Publishing Company. Belmont, California. Plant Physiol. 682 p. [ Links ]
Schuch, U. K. 2003. Forcing containerized roses in a retractable roof greenhouse and outdoors in a semi-arid climate. Plant Sciences Department. University of Arizona, Tucson. 6 p. [ Links ]
Smith, I. E.; Savage, M. J. and Mills, P. 1984. Shading effects on greenhouse tomatoes and cucumbers. Int. Soc. Hortic. Sci. 148:491-500. [ Links ]
Spomer, G. G. 1973. The concepts of "interaction" and "operational environment" in environmental analyses. Ecology. 54(1):200-204. [ Links ]
Stanhill, G. 1979. The energy cost of protected cropping: a comparison of six systems of tomato production. J. Agric. Eng. Res. 25:145-154. [ Links ]
Stern, R. K. 1994. Introductory plant biology. 6th edition. WCB.USA.94p. [ Links ]
Strohman, R. C. 1997. Epigenesis and complexity, the coming kuhnian revolution in biology. Nature Biotechnology. 15:194-200. [ Links ]
Taiz, L. and Zeiger, E. 2002. Plant Physiol. 3th edition. Sinaver Associates, Inc. Punlishers Sunderland, Massachusetts. USA. 175 p. [ Links ]
Terashima, I.; Hanba, T. Y.; Tazoe, Y.; Vyas, P. andYano, S. 2006. Irradiance andphenotype: comparative eco-development of sun and shade leaves in relation to photosynthetic CO2. J. Exp. Bot. 57:343-354. [ Links ]
Verhaegh, A. P. and de Groot, N. S. 2000. Chain production costs of fruits vegetables: a comparison between Spain and the Netherlands. Acta Hortic. 524:177-180. [ Links ]
Vollebregt, R. 2010. Retractable roof greenhouse. 2010. Re-writing the guidelines for greenhouse design and crop management. Cravo Canada. 91 p. [ Links ]
Vollebregt, R. 2004. The potential of retractable roof greenhouses to dominate greenhouse designs in the future. ISHS. ActaHortic. 633:43-49. [ Links ]
Whitelam, G. C. and Franklin, K. A. 2004. Light signals, phytochormes and crosstalk with otherenvironmental cues. J. Exp. Bot. 55:271-276. [ Links ]
Zhang, L.; Hao, X. M.; Li, Y. G. and Jiang, G. M. 2010. Response of greenhouse tomato to varied low pre-night temperatures at the same daily integrated temperature. HortScience. 45(11): 1654-1661. [ Links ]

