










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.3 no.1 Texcoco ene./feb. 2012
Artículos
Relación nitrato/amonio/urea y concentración de potasio en la producción de tomate hidropónico*
Relation nitrate / ammonium / urea and potassium concentration in hydroponic tomato production
Saúl Parra Terraza1§, Guadalupe Mendoza Palomares1 y Manuel Villarreal Romero1
1Facultad de Agronomía. Universidad Autónoma de Sinaloa. Carretera Culiacán-El Dorado, km 17.5. Culiacán, Sinaloa, México. A. P. 726, C. P. 80000. Tel. 01 667 7543693. (pipo_m_p@hotmail.com), (manuelvillarreal2@yahoo.mx).§Autor para correspondencia: psaul@uas.uasnet.mx.
*Recibido: abril de 2011
Aceptado: noviembre de 2011
Resumen
Plantas de tomate (Solanum lycopersicum L. cv. IB-9) fueron cultivadas en un sistema hidropónico con el objetivo de evaluar el efecto de la relación nitrato, amonio, urea (100/0/0, 85/15/0, 85/0/15y85/7.5/7.5) y la concentración de potasio (7 y 9 mol m-3) en el rendimiento de fruto y composición mineral de las plantas. El rendimiento de fruto no fue afectado por los tratamientos, por lo que se puede sustituir 15% del total de nitrógeno nítrico de la solución nutritiva con un porcentaje igual de nitrógeno amoniacal o ureico, o una mezcla de amonio y urea, cada una a 7.5%. El contenido de fósforo en las hojas aumentó significativamente con la interacción de 85/15/0 y 9 mol m-3 de potasio, mientras que la concentración de calcio en las hojas disminuyó estadísticamente con 100/0/0 y 9 mol m-3 de potasio.
Palabras clave: Solanum lycopersicum L., contenido nutrimental, hidroponía, soluciones nutritivas.
Abstract
Tomato plants (Solanum lycopersicum L. cv. IB-9) were grown in ahydroponic system in orderto evaluate the effect of the ratio nitrate, ammonium, urea (100/0/0, 85/15/0, 85 / 0/15 and 85/7.5/7.5) and the concentration of potassium (7 and 9 mol m-3) in fruit yield and mineral composition of plants. The fruit yield was unaffected by the treatments, so that it can substitute 15% of the total nitrate nitrogen to the nutrient solution with an equal percentage of nitrogen or urea, or a mixture of ammonia and urea, each at 7.5 %. The phosphorus content in leaves was significantly increased by the interaction of 85/15/0 and 9 mol of potassium m-3, while the calcium concentration in leaves 100/0/0 statistically decreased and 9 mol m-3 potassium.
Key words: Solanum lycopersicum L., nutrient content, hydroponic, nutrient solutions.
Introducción
El nitrógeno es un elemento esencial para el crecimiento y el metabolismo de las plantas; puede suministrarse en tres diferentes formas: nítrica, amoniacal y ureica (Parra et al., 2010). La forma nítrica es absorbida preferentemente por la mayoría de las plantas (Mengel y Kirkby, 2000; Miller y Cramer, 2004), por lo que es la más utilizada en los cultivos hidropónicos. La respuesta a la nutrición amoniacal varía entre especies y las condiciones ambientales (Kotsiras et al., 2005), en ciertas concentraciones puede resultar tóxica para muchas especies vegetales (Salsac et al., 1987), por lo que generalmente se recomienda aplicar el amonio en pequeñas concentraciones, discrepando los investigadores en las concentraciones a usar (Steiner, 1984; Sandoval et al., 1995;Portree, 1997).
La urea es una de las principales fuentes de fertilizantes nitrogenados para la producción de plantas (Vavrinay Obreza, 1993); no obstante, rara vez se usa en sistemas hidropónicos (Ikeda y Xuewen, 1998), debido a la escasa investigación relacionada con la absorción y la utilización de dicha fuente (Khan et al., 2000). El uso potencial de la urea como sustituto de los fertilizantes nítricos en hidroponía, podría reducir la acumulación excesiva de nitratos en las plantas (Luo et al., 1993) y los costos de producción, ya que la urea es un fertilizante más barato que los fertilizantes nítricos. Está documentado que varias especies de plantas pueden incrementar su crecimiento, con aportes combinados de nitrato y amonio a diferencia de cuando se aporta cualquiera de las dos formas de nitrógeno por separado (Lips et al., 1990; Errebhi y Wilcox, 1990); sin embargo, hay diferencias en la tolerancia de especies y cultivares respecto ala concentración de amonio en la relación nitrato/amonio (Claussen y Lenz, 1999).
El tomate está considerado como una especie sensible al amonio (Gerendas et al., 1997), por lo que la concentración de N amoniacal debe ser entre tres y diez porciento del total de nitrógeno suministrado (Portree, 1997; Steiner, 1984). La absorción de amonio está influenciada por la presencia de nitratos (Britto y Kronzucker, 2002) y potasio (Szczerba et al., 2006) en la solución, los cuales pueden reducir la toxicidad del amonio. La absorción de nitratos incrementa la concentración de algunas hormonas del crecimiento en el xilema (Rahayu et al., 2005), regulando la división y la expansión celular (Francis y Sorell, 2001), mientras que el potasio tiene similitudes con el amonio, en cuanto a la valencia, el diámetro del ion hidratado y su efecto sobre el potencial eléctrico de la membrana (Wang et al., 1996; Xu et al., 2002) por lo que hay interacción entre estos dos iones.
El amonio puede influir en la absorción y acumulación de potasio posiblemente por que compite con el para ingresar a las células (Szczerba et al., 2006b); mientras que el potasio puede inhibir la absorción de amonio en cultivos de maíz y cebada (Wang et al., 1996) o proteger a las plantas de toxicidad debido a altas concentraciones de amonio (Zhu et al., 1998; Santa-María et al., 2000). El tomate absorbe grandes cantidades de potasio, mayores que cualquier otro nutrimento (Chen y Gabelman, 1999) por lo que un adecuado suministro de potasio puede favorecer el uso del amonio, si se considera que la toxicidad por amonio puede atribuirse a la disminución en la absorción de cationes (Marschner, 1995), a la alteración del balance osmótico (Gerendas et al., 1997) y a la síntesis limitada de aniones orgánicos (Salsac et al., 1987). El objetivo del presente estudio fue evaluar el efecto de la relación porcentual de nitrato/amonio/urea y la concentración de potasio en la solución nutritiva sobre el rendimiento de fruto y la composición mineral de tomate en hidroponía.
Materiales y métodos
El estudio se realizó bajo malla sombra al 50% (porcentaje de luz que bloquea la malla), en la Facultad de Agronomía de la Universidad Autónoma de Sinaloa, México. Plántulas de tomate (Solanum lycopersicum L. cv. IB-9), de crecimiento indeterminado, tipo bola, crecieron hasta su trasplante en charolas de poliestireno de 200 cavidades con un volumen unitario de 30 cm3 regadas con soluciones nutritivas con las mismas composiciones químicas (Parra et al., 2010), que las aplicadas en el presente trabajo. El diseño experimental fue completamente al azar con arreglo factorial 4*2, con cuatro repeticiones por tratamiento. La unidad experimental estuvo compuesta por dos recipientes de plástico con capacidad individual de 20 L interconectados entre sí con manguera de plástico de 1.25 cm de diámetro.
Uno de los recipientes contenía 18 L de tezontle negro como sustrato con una granulometría de 2 a 5 mm de diámetro (Cuadro 1), con una planta de tomate por recipiente, y el otro recipiente, 10 L de solución nutritiva del tratamiento correspondiente para efectuar los riegos por subirrigación. Se aplicaron dos riegos diarios, el primero a las 8:00 am y el segundo a las 13:00 pm; diariamente se repuso el agua evapotranspirada aforando a 10 L con agua destilada y el pH se ajustó a 5.5 ± 0.1 con HCl o NaOH 1N; las soluciones nutritivas se renovaron cada 15 días. Los tratamientos fueron ocho soluciones nutritivas resultantes de la combinación de cuatro relaciones porcentuales de NO3-NH4+/CO(NH2)2( 100/0/0, 85/15/0, 85/0/15 y 85/7.5/7.5) y dos concentraciones de K+ (7 y 9 mol m-3).
Los tratamientos se diseñaron a partir de la solución universal de Steiner(1984) (tratamiento 1, Cuadro 2); esta solución no incluye al amonio ni a la urea como fuentes de N, por lo que las modificaciones fueron: 1) adición de una concentración de NH4+ y reducción equivalente de la concentración de NO3-; 2) disminución de las concentraciones relativas de los aniones (NO3-, H2PO4-, SO42) e incrementos de las concentraciones de H2PO4- y SO42-, hasta igualar la concentración de NH4+ añadido y mantener la relación mutua de H2PO4- y SO42- (1:3.5); 3) la inclusión de NH4+ incrementó las concentraciones relativas de los cationes (K+, Ca2+, Mg2+ y NH4+), por lo que la concentración de NH4+ añadida se restó a las concentraciones de K+, Ca2+ y Mg2+, considerando su relación mutua, que permitió mantener la relación mutua entre Ca2+y Mg2+(2.25:1); 4) las modificaciones anteriores provocaron un cambio en el potencial osmótico de la solución nutritiva, por lo que se ajustaron las concentraciones de los iones mediante el factor de corrección (-0.0024), propuesto por Steiner (1984).
Las soluciones nutritivas se prepararon con sales inorgánicas grado reactivo y agua d estilada y se les añadieron las concentraciones de micronutrimentos (enmg L-1) siguientes: Fe 2.5, Mn 0.5, B0. 5, Cu0 .02 y Zn 0.05.El Fese proporcionó como Fe-EDTA (Steiner y Van Winden, 1970).
El periodo de cosecha inició el 23 de enero y terminó el 23 de marzo de 2009. Se evaluó rendimiento (kg planta-1) de frutos rojos y frutos verdes; rendimiento total (frutos rojos + frutos verdes), número de frutos rojos por planta y el peso promedio de frutos rojos (g planta-1). En el último corte de frutos las plantas se cosecharon y separaron en hojas, tallos, frutos (rojos y verdes); estos órganos, con excepción de los frutos verdes, se secaron en estufa con circulación forzada de aire a una temperatura de 70 °C durante 72 h, se molieron en un molino eléctrico y tamizaron con malla 40.
Al material tamizado se le realizó una digestión seca en una mufla a 550 °C durante 5 h y se determinaron las concentraciones de P, K, Ca y Mg. El P se cuantificó por el método amarillo vanadato molibdato (Rodríguez y Rodríguez, 2002), Kporflamometría (Alcántar y Sandoval, 1999), Ca y Mg por titulación con EDTA (Chavira y Castellanos, 1987); mientras que el N se determinó por el procedimiento semi-micro Kjeldhal (Bremner, 1965) modificado para incluir nitratos. El análisis de varianza de las variables consideradas se realizó para los factores principales y su interacción con el programa SAS (1999) versión ocho. Para la comparación de medias se utilizó la prueba de Tukey (a< 0.05).
Resultados y discusión
Rendimiento
El peso de frutos totales (rojos + verdes), número de frutos rojos y el peso promedio de frutos rojos, no presentaron diferencias significativas por efecto de los tratamientos. El peso de frutos totales varió de 4.372 a 4.578 kg planta-1, el número de frutos rojos por planta de 17.3 a 19.5 y el peso promedio de frutos rojos varió de 148.3 a 155.8 g planta-1 (Cuadro 3). Estos resultados contrastan con los de Siddiqi et al. (2002) y Bialczyk et al. (2007), quienes al añadir 10% y 20% del nitrógeno total en forma de NH4+ a la solución nutritiva, obtuvieron incrementos de 15% y 20% respectivamente en el rendimiento de tomate cultivares Trust F1y Perkoz F1 comparado con la solución de nitrato al 100%, lo cual puede atribuirse a que los cultivares de tomate responden diferente a las fuentes de nitrógeno (Ben-Oliel et al., 2004), a variaciones en las condiciones ambientales (Kotsiras et al., 2005) y experimentales prevalecientes en ambos estudios.
La adición de 15% de nitrógeno total en forma de urea a la solución nutritiva, tampoco afectó estadísticamente el peso de frutos totales de tomate, cultivar IB-9 (Cuadro 3). Lo anterior, puede explicarse considerando que en hidroponía, igual que en el suelo la urea puede ser hidrolizada a amonio por la enzima ureasa (Bundy y Bremner, 1974) con la subsecuente conversión de amonio a nitrato mediante el proceso de nitrificación (Aminuddin et al., 1993; Luo et al., 1993). Luo et al. (1993) al evaluar las transformaciones de la urea en condiciones de hidroponía, encontraron que en 28 días se realizó la transformación de urea a amonio y nitrato, por ello mencionan que la urea en solución puede ser considerada como una mezcla de nitrógeno inorgánico y orgánico. Aminuddin et al. (1993) indican que en un sistema hidropónico de técnica en película nutritiva (NFT) la hidrólisis de la urea inicia el séptimo día y finaliza el día 20.
En el presente estudio la solución nutritiva fue reemplazada cada 15 días, por lo que posiblemente se presentó el tiempo suficiente para que gran parte de la urea fue transformada a amonio y nitrato, lo que sugiere que la aplicación de un cierto porcentaje de urea en la solución nutritiva, puede reemplazar al nitrato en la solución nutritiva sin afectar el rendimiento, además de reducir la acumulación de nitratos en frutos de vegetales como el tomate (Tan et al., 2000). Guins et al. (1994); Zhu et al. (1997) reportaron que la sustitución de 20 y 25 % de nitratos por urea en la solución nutritiva, no afectó el crecimiento de lechuga tampoco en pak choi (Brassicca pequinensis L.).
Concentración de nutrimentos en hojas
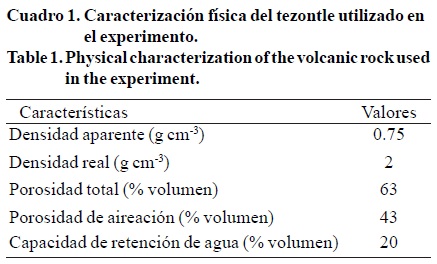
Las concentraciones de fósforo y de calcio fueron significativamente afectadas por la interacción relación porcentual nitrato/amonio/urea*concentración de potasio (Cuadro 4). Con las relaciones 100/0/0, 85/0/15 y 85/7.5/7.5 los contenidos de fósforo en las hojas fueron estadísticamente iguales para ambas concentraciones de potasio, mientras que el contenido de fósforo (0.383%) para la relación 85/15/0 con 9 mol m-3 de potasio, fue significativamente mayor comparada con esa misma relación y 7 mol m-3 de potasio (Figura 1).
Marschner (1995) menciona que las plantas suministradas con nitrógeno amoniacal, generalmente contienen altos contenidos de aniones; por ejemplo, el fósforo para mantener el equilibrio de aniones y cationes en las plantas, lo que explica este aumento significativo de fósforo en las hojas. Parra et al. (2010) reportaron que con una relación 85/15/0 y 9 mol m-3 de potasio en la solución nutritiva, la concentración de P en hojas de plántula de tomate, fue significativamente mayor con respecto a los valores de Pobtenidos con las relaciones 100/0/0, 85/0/15 y 85/7.5/7.5, lo que coincide con los resultados de este estudio e indica un efecto similar de la relación 85/15/0 sobre el contenido de fósforo en una plántula joven y una planta madura.
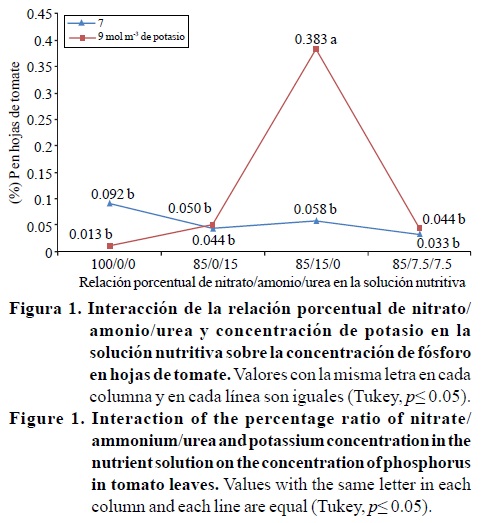
En la Figura 2 se observa que la relación porcentual nitrato/ amonio/urea no afectó estadísticamente los contenidos de calcio en las hojas para ambas concentraciones de potasio; con 9 mol m-3 de potasio en la solución nutritiva el contenido de calcio (3.92%) para la relación 100/0/0, fue significativamente mayor comparado con las otras relaciones, las cuales fueron estadísticamente iguales entre sí (2.28, 2.43 y 2.10% de calcio para las relaciones 85/0/15, 85/15/0 y 85/7.5/7.5). Este resultado coincide con lo reportado por diversos autores (Marschner, 1995; Mengel y Kirkby, 2000), quienes mencionan que las plantas que absorben preferencialmente los nitratos en lugar del amonio contienen niveles altos de cationes, entre los cuales está el calcio. Siddiqi et al. (2002) reportaron una reducción en la concentración de calcio en hojas de tomate debido al amonio, lo cual coincide con el resultado obtenido en este estudio (Cuadro 4).
Concentración de nutrimentos en tallos
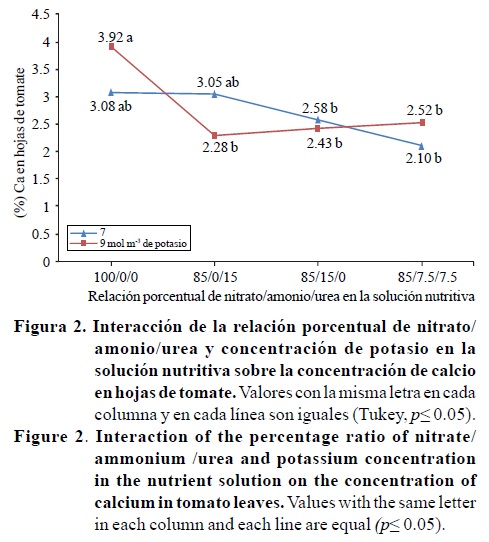
Las concentraciones de nitrógeno y magnesio en los tallos, fueron significativamente afectados por la interacción de los factores relación porcentual nitrato/amonio/urea*concentración de potasio; mientras que el contenido de fósforo presentó diferencias estadísticas por efecto del factor relación porcentual nitrato/amonio/urea (Cuadro 4). Con 7 mol m-3 de potasio en la solución nutritiva el contenido de nitrógeno fue de 2.1 % para las relaciones 85/0/15 y 85/15/0, valores significativamente mayores al obtenido con 100/0/0 y estadísticamente igual a 85/7.5/7.5 (Figura 3). La Figura 4 indica que con la relación 85/7.5/7.5 y 9 mol m-3 de potasio el contenido de magnesio fue de 0.8%, valor estadísticamente mayor al obtenido con esa misma relación y 7 mol m-3 de potasio (0.2%).
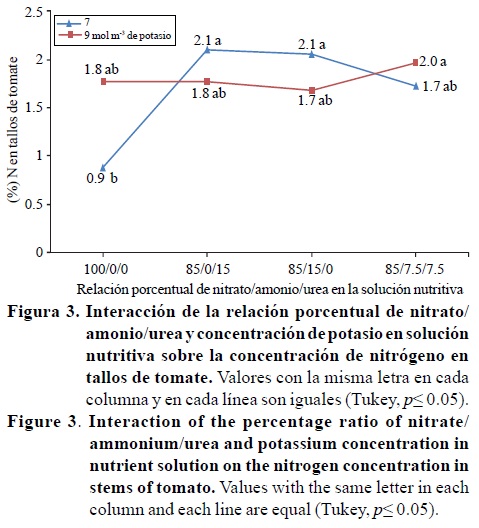
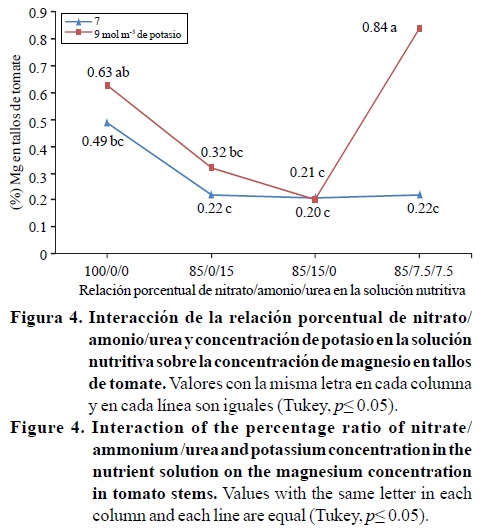
También se observa en la Figura 4 que al incrementar el contenido de amonio (relación 85/15/0) o de urea (relación 85/0/15) los contenidos de magnesio bajan significativamente a0.2y 0.3%, respectivamente, lo que es similar a lo reportado por otros investigadores (Mengel y Kirkby, 2000; Marschner, 1995), quienes mencionan que el amonio puede inhibir la absorción y acumulación de cationes, como el magnesio. Los contenidos de fósforo en los tallos (0.3 7 y 0.3 %) obtenidos con las relaciones 85/15/0 y 85/0/15, fueron significativamente mayores al obtenido con 100/0/0 (0.18%), y estadísticamente igual a 85/7.5/7.5 (0.29%) (Cuadro 4), que coincide con lo reportado por Mengel y Kirkby (2000); Marschner (1995), quienes mencionan que las plantas suministradas con nitrógeno amoniacal, generalmente contienen altas concentraciones de aniones, por ejemplo fósforo, ya que el amonio favorece su absorción y acumulación para mantener la electroneutralidad a nivel celular (Marschner, 1995).
Concentración de nutrimentos en frutos rojos
La concentración de magnesio en los frutos rojos, fue significativamente afectada por la interacción entre la relación porcentual nitrato/amonio/urea y la concentración de potasio; mientras que la concentración de calcio fue estadísticamente diferente en la relación porcentual nitrato/amonio/urea (Cuadro 4). Con la relación 85/7.5/7.5 y 9 mol m-3 de potasio el contenido de magnesio fue 0.081 %, valor estadísticamente mayor al obtenido con esa misma relación y 7 molm-3 de potasio (0.025%) (Figura 5).
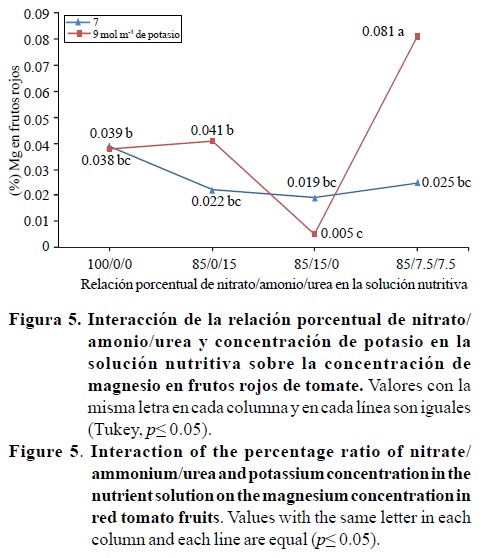
El contenido de calcio obtenido con la relación 100/0/0 fue 0.13%, valor significativamente menor a los obtenidos con las relaciones 85/15/0 y 85/7.5/7.5 (0.16 y 0.17%) (Cuadro 4). Este resultado es difícil de explicar, ya que generalmente la absorción de calcio es limitada por la competencia del amonio (Marschner, 1995; Mengel y Kirkby, 2000); por lo tanto, se esperaría que los frutos de tomate provenientes de las relaciones 85/15/0 y 85/7.5/7.5, tuvieran una concentración menor de calcio comparado con la relación 100/0/0.
Este resultado difiere de lo reportado por Siddiqi et al. (2002), quienes encontraron que la concentración de amonio en la solución nutritiva redujo las concentraciones de calcio en los frutos. Probablemente, estas diferencias en los contenidos de calcio en los frutos del presente estudio con el de ellos, pueden ser atribuidas a variaciones en los genotipos o condiciones ambientales diferentes prevalecientes en ambos trabajos.
Conclusiones
Los resultados indican que la sustitución de 15% del total de nitrógeno nítrico en la solución nutritiva, por un porcentaje similar de nitrógeno amoniacal y ureico o una mezcla 7.5% N-amoniacal-7.5% N-ureico, pueden ser utilizadas sin afectar el rendimiento de tomate cultivar IB-9 en hidroponía. La interacción de la relación porcentual nitrato/amonio/urea y la concentración de potasio en la solución nutritiva, afectaron estadísticamente las concentraciones de fósforo y calcio en hojas, nitrógeno y magnesio en tallos, y magnesio en frutos rojos.
Agradecimientos
A la Universidad Autónoma de Sinaloa por su apoyo financiero, a través del Programa de Fortalecimiento y Apoyo a Proyectos de Investigación (PROFAPI-2008).
Literatura citada
Alcántar, G. G. y Sandoval, V. M. 1999. Manual de análisis químico de tejido vegetal. Publicación Especial 10. Sociedad Mexicana de la Ciencia del Suelo. Chapingo, México. 156 p. [ Links ]
Aminuddin, H.; Khalip, R.; Norayah, K. y Alias, H. 1993. Urea as the nitrogen source in NFT hydroponic system. Pertanika J. Trop. Agric. Sci. 16:87-94. [ Links ]
Ben-Oliel, G.; Kant, S.; Naim, M.; Rabinowitch, H. D.; Takeoka, G. R.; Buttery, R. G. andKafkafi, U. 2004. Effects of ammonium to nitrate ratio and salinity on yield and fruit quality of large and small tomato fruit hybrids. J. Plant Nutr. 27:1795-1812. [ Links ]
Bremner, J. M. 1965. Total nitrogen. In: Black, C. A. (ed.). Methods of soil analysis. (Part 2). (Agronomy 9). American Society of Agronomy. Madison, WI. USA. 1149-1178 p. [ Links ]
Bialczyk, J.; Lechowski, Z.; Dziga, D. and Mej, E. 2007. Fruit yield of tomato cultivated on media with bicarbonate and nitrate/ammonium as the nitrogen source. J. Plant Nutr. 30:149-161. [ Links ]
Britto, D. T. and Kronzucker, H. J. 2002. NH4+ toxicity in higher plants. J. Plant Physiol. 159:567-584. [ Links ]
Bundy, L. G. and Bremner, J. M. 1974. Effects of nitrification inhibitors on transformation of urea N in soils. Soil Biochem. 6:369-376. [ Links ]
Chavira, R. J. C. y Castellanos, R. J. Z. 1987. Sales solubles. In: Aguilar, S. A.; Etchevers, J. D. y Castellanos, R. J. Z. (eds.). Análisis químico para evaluar la fertilidad del suelo. Publicación Especial Núm. 1. Sociedad Mexicana de la Ciencia del Suelo. 109-124p. [ Links ]
Chen, J. and Gabelman, W. H. 1999. Potassium transport rate from root to shoot unrelated to potassium-use efficiency in tomato grown under low-potassium stress. J. Plant Nutr. 22:621-631. [ Links ]
Claussen, W. and Lenz, F. 1999. Effect of ammonium or nitrate nutrition on net photosynthesis, growth, and activity of the enzymes nitrate reductase and glutamine synthetase in blueberry, raspberry and strawberry. Plant Soil. 208:95-102. [ Links ]
Errebhi, M. and Wilcox, G. E. 1990. Plant species response to ammonium-nitrate concentration ratios. J. Plant Nutr. 13:1017-1029. [ Links ]
Francis, D. and Sorell, D. A. 2001. The interface between the cell cycle and plant growth regulators: a mini review. Plant Growth Regulation. 33:1-12. [ Links ]
Gerendas, J.; Zhu, Z.; Bendixen, R.; Ratcliffe, R. G. and Sattelmacher, B. 1997. Physiological and biochemical processes related to ammonium toxicity in higher plants. Z. Pflanzenernaehr. Bodenkd. 160:239-251. [ Links ]
Guins, A.; Post, W.N. K.; Kirkby, E. A. and Aktas, M. 1994. Influence of partial replacement of nitrate by amino acid nitrogen or urea in the nutrient medium on nitrate accumulation in NFT grown winter lettuce. J. Plant Nutr. 17:1929-1938. [ Links ]
Ikeda,H. and Xuewen,T. 1998. Urea as an organic nitrogen source for hydroponically grown tomatoes in comparison with inorganic nitrogen sources. Soil Sci. Plant Nutr. 44:609-615. [ Links ]
Khan, N. K.; Watanabe, M. and Watanabe, Y. 2000. Effect of partial urea application on nutrient absorption by hydroponically grown spinach. Soil Sci. Plant Nutr. 46:199-208. [ Links ]
Kotsiras, A.; Olympios, C. M. and Passam, H. C. 2005. Effects of nitrogen form and concentration on yield and quality of cucumbers grown on rockwool during spring and winter in southern Greece. J. Plant Nutr. 28:2027-2035. [ Links ]
Lips, S. H.; Leidi, E. O.; Silberbush, M.; Soars, M. I. M. and Lewis, O. E. M. 1990. Physiological aspects of ammonium and nitrate fertilization. J. Plant Nutr. 13:1271-1289. [ Links ]
Luo, J.; Lian, Z. and Yan, X. 1993. Urea transformation and the adaptability of three Leary vegetables to urea as a source of nitrogen in hydroponic culture. J. Plant Nutr. 16:797-812. [ Links ]
Marschner, H. 1995. Mineral nutrition of higher plants. Second edition. Academic Press. San Diego, California. USA. 231-254 pp. [ Links ]
Mengel, K. y Kirkby, E. A. 2000. Principios de nutrición vegetal. Traducción al español de la 4ta edición de Melgar, R. J. y Ruíz, M. Internacional Potash Institute. Basel, Switzerland. 135-146 p. [ Links ]
Miller, A. J. and Cramer, M. D. 2004. Root nitrogen acquisition and assimilation. Plant Soil. 274:1-36. [ Links ]
Parra, T. S.; Salas, N. E.; Villarreal, R. M.; Hernández, V. S. y Sánchez, P. P. 2010. Relaciones nitrato/amonio/ urea y concentraciones de potasio en la producción de plántulas de tomate. Revista Chapingo. Serie Horticultura. 16:37-46. [ Links ]
Portree, J. 1997. Greenhouse vegetable production guide for commercial growers. British Columbia Ministry of Agriculture, Fisheries and Food. [ Links ]
Rahayu, Y. S.; Walch-Liu, P.; Neumann, G.; Romheld, V.; Von Wiren, N. and Bangerth, F. 2005. Root-derived cytokinins as long-distance signals for NO3- induced stimulation of leaf growth. J. Exp. Bot. 56:1143-1152. [ Links ]
Rodríguez, H. y Rodríguez, J. 2002. Métodos de análisis de suelos y plantas: criterios de interpretación. Editorial Trillas, S. A. de C. V. D. F., México. 155-157pp. [ Links ]
Salsac, L.; Chaillou, S.; Morot-Gaudry, J.; Lesaint, C. and Jolivert, E. 1987. Nitrate and ammonium nutrition in plants. Plant Physiol. Biochem. 25:805-812. [ Links ]
Sandoval, V. M.; Alcántar, G. G. and Tirado, T. J. L. 1995. Use of ammonium in nutrient solutions. J. Plant Nutr. 18:1449-1457. [ Links ]
Santa-María, G. E.; Danna, C. H. and Czibener, C. 2000. High-affinity potassium transport in barley roots: Ammonium-sensitive and-insensitive path-ways. Plant Physiol. 123:297-306. [ Links ]
Statistical Analysis System (SAS) Institute. 1999. User's guide. Versión 8. SAS Institute Inc. Cary, N. C. USA. [ Links ]
Siddiqi, M. Y.; Malhotra, B.; Min, X. and Glass, A. D. M. 2002. Effects of ammonium and inorganic carbon enrichment on growth and yield of a hydroponic tomato crop. J. Plant Nutr. Soil Sci. 165:191-197. [ Links ]
Steiner, A. A. and van Winden, H. 1970. Recipe for ferric salts of ethylenediaminetetraacetic acid. Plant Physiol. 46:862-863. [ Links ]
Steiner, A. A. 1984. The universal nutrient solution. Sixth International Congress on Soilless Culture. ISOSC Proceeding. The Netherlands. 633-649 pp. [ Links ]
Szczerba, M. W.; Britto, D. T. and Kronzucker, H. J. 2006. Rapid, futile K+ cycling and pool-size dynamics define low-affinity potassium transport in barley. Plant Physiol. 141:1494-1507. [ Links ]
Szczerba, M. W.; Britto, D. T. and Kronzucker, H. J. 2006b. The face value of ion fluxes: the challenge of determining influx in the low-affinity transport range. J. Exp. Bot. 57:3293-3300. [ Links ]
Tan, X. W.; Ikeda, H. and Oda, H. 2000. The absorption, translocation, and assimilation of urea, nitrate or ammonium in tomato plants at different plant growth stages in hydroponic culture. Hortic. Sci. 84: 275-283. [ Links ]
Vavrina, C. S. and Obreza, T.A.1 993. Response of chinese cabbage to nitrogen rate and source in sequential plantings. HortScience. 28:1164-1165. [ Links ]
Wang, M. Y.; Siddiqui, M. Y. and Glass, A. D. M. 1996. Interaction between K+ and NH4+: effects onion uptake by rice roots. Plant Cell Environ. 19:1037-1046. [ Links ]
Xu, G.; Wolf, S. and Kafkafi, U. 2002. Ammonium on potassium interaction in sweet pepper. J. Plant Nutr. 25:719-734. [ Links ]
Zhu, Z.; Gerendas, J. and Sattelmacher, B. 1997. Effects of replacing of nitrate with urea or choride on the growth and nitrate accumulation in pak-choi in the hydroponics. In: Ando, T.; Fujita, K.; Mae, T.; Matsumoto, H.; Mori, S. and Sekiya, J. (eds.). Plant nutrition for sustainable food production and environment. Kluwer Academic Publishers, Dordrecht. 963-964 pp. [ Links ]
Zhu, J. K.; Liu, J. and Xiong, L. 1998. Genetic analysis of salt tolerance in Arabidobsis thaliana: evidence for a critical role of potassium nutrition. Plant Cell. 10:1181-1191. [ Links ]

