










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.2 no.6 Texcoco nov./dic. 2011
Artículos
Cinética de crecimiento foliar y desarrollo de brotes en selecciones injertadas de zapote mamey*
Leaf growth kinetics and shoot development in grafted selections of mamey sapote
Alejandro Ledesma Miramontes1, Ángel Villegas Monter1§, Víctor Arturo González Hernández2, Lucero del Mar Ruiz Posadas3 y Antonio Mora Aguilera4
1Fruticultura. Colegio de Postgraduados. Carretera México-Texcoco, km 36.5. C. P. 56230. Montecillo, Texcoco, Estado de México. Tel. 01 595 9520200. Ext. 1565. (aledesma@colpos.mx). § Autor para correspondencia: avillega@colpos.mx.
2Fisiología Vegetal. Colegio de Postgraduados. Tel. 01 595 9520200. Ext. 1565. (vagh@colpos.mx).
3Botanica. Colegio de Postgraduados. Tel. 01 595 9520200. Ext. 1300. (lucpo@colpos.mx).
4Fitopatología. Colegio de Postgraduados. Tel. 01 595 9520200. Ext. 1611. (aguilera@colpos.mx).
* Recibido: enero de 2011
Aceptado: agosto de 2011
RESUMEN
El objetivo de este trabajo fue evaluar el crecimiento de órganos vegetativos en seis selecciones de zapote mamey (Pouteria sapota (Jacq.) H. E. Moore & Stearn), injertadas de tres y cuatro años en las condiciones edafoclimáticas de Alpoyeca, Guerrero; para conocer el comportamiento de árboles procedentes de Yucatán y Alpoyeca, Guerrero, se evaluaron: cinética del crecimiento foliar, crecimiento total del brote, índice de área foliar, peso específico de la hoja y grosor de la hoja. Los resultados mostraron que las hojas de mamey alcanzan el máximo desarrollo a 42 días, pero a 23 días tienen más de 80% del tamaño total y pueden ser fotosintéticamente autosuficientes. El crecimiento total de brotes durante un año en árboles de 3 años, fue de 62.9 a 72 cm, y el periodo de mayor crecimiento de junio a septiembre, mientras que en los de 4 años, fue de 40.7 a 44.1 cm y el periodo de mayor crecimiento de abril a agosto. 'El Regalo' seleccionado en Alpoyeca, Guerrero; presentó valores mayores en índice de área foliar, peso específico y grosor de la hoja, mientras que en 'Dany 66' de Yucatán fueron los menores. El peso específico de la hoja es una variable que se modifica con la fenología de la planta, por lo que se debe evaluar mínimo en tres etapas para tener resultados confiables.
Palabras claves: Pouteria sapota, grosor de hoja, índice área foliar, peso específico de hoja, Sapotaceae.
ABSTRACT
The aim of this study was to evaluate the growth of vegetative organs in six selections of mamey sapote (Pouteria sapota (Jacq.) H. E. Moore & Stearn), grafted of three and four years in the soil's and climatic conditions of Alpoyeca, Guerrero; in order to understand the behavior of trees from Alpoyeca Yucatán, Guerrero, were evaluated: kinetics of leaf growth, total growth of shoot, leaf area index, leaf unit weight and thickness of the leaf. The results showed that mamey's leaves reach their maximum development at 42 days but, at 23 days they have 80% of their total size and can be photosynthetically self-sufficient. The total growth of buds on the 3 year old trees during one year was 62.9 to 72 cm, and the period of highest growth was from June to September, while in the 4-year old trees was of 40.7 to 44.1 cm and the period of highest growth from April to August. 'El Regalo' selected in Alpoyeca, Guerrero, presented the highest values of leaf area index, specific leaf weight and thickness of the leaf, while 'Dany 66' from Yucatán had the lowest. The specific weight of the leaf is a variable that changes with the phenology of the plant, which should be evaluated at least three stages to get reliable results.
Key words: Pouteria sapota, leaf area index, leaf thickness, Sapotaceae, specific leaf weight.
INTRODUCCIÓN
El zapote mamey (Pouteria sapota (Jacq.) H. E. Moore & Stearn), es un frutal nativo de México y Centroamérica con alto potencial económico, ya que existe demanda de la fruta en varios países. Pero debido que la superficie cultivada en México no rebasa 1 500 ha, son pocos los trabajos que se han realizado para conocer la fenología y respuesta fisiológica a diversas prácticas de manejo. En general, cuando se evalúan arboles injertados, se analizan: crecimiento, rendimiento, calidad de la fruta, vigor, incidencia de plagas y enfermedades, adaptación al medio y contenido de elementos minerales en hojas (Figueiredo et al., 2002; Quijada et al., 2002).
Sin embargo, en pocas ocasiones se considera el efecto del portainjerto en el crecimiento de la planta (Ayala-Arreola et al., 2010; Arrieta-Ramos et al., 2010; Berdeja-Arbeu et al., 2010). La edad de la hoja y su estado de desarrollo son importantes para determinar algunos procesos morfológicos y fisiológicos en las plantas (Horsley y Gottschalk, 1993). De igual forma, conocer el área foliar es fundamental en estudios de nutrición y crecimiento vegetal, porque se puede determinar acumulación de materia seca, metabolismo de carbohidratos, rendimiento y calidad de la cosecha, además es necesaria para evaluar la intensidad de asimilación de las plantas (Bugarin et al., 2002).
El peso específico de la hoja (PEH), determina la cantidad relativa de tejido fotosintético tomando como base el peso de materia seca por unidad de área, el cual es afectado por la intensidad y duración de la luz, puede ser utilizada como una forma sencilla de medir fotosíntesis en plantas leñosas (Marini y Sowers, 1990). Además el grosor de la hoja puede relacionarse con el PEH debido al número de capas de células en empalizada y tamaño de estas, además el sombreado incrementa el tamaño de la hoja, disminuye el grosor y altera la estructura interna (Marini y Sowers, 1990).
Aun cuando el grosor de la hoja es afectado por la especie (Reyes-Santamaría et al., 2000), el ambiente (Ibarra, 2005), y uso de portainjerto (Ayala-Arreola et al., 2010), es importante conocerlo en zapote mamey, para que pueda ser empleado como referencia en futuras investigaciones. Con base en lo anterior, el objetivo de este trabajo fue evaluar el crecimiento de hojas y brotes en selecciones de zapote mamey, en las condiciones edafoclimáticas de Alpoyeca, Guerrero; para conocer el comportamiento fisiológico de árboles injertados de zapote mamey, procedentes de Yucatán y Alpoyeca, Guerrero.
MATERIALES Y MÉTODOS
El trabajo fue realizado en una huerta comercial con fertirrigación, en el municipio de Alpoyeca, Guerrero, México; ubicada entre las coordenadas 17° 31' y 17° 40' de latitud norte y 98° 01' y 98° 31' de longitud oeste y 1 030 m de altitud. El clima de la región es del tipo (A) C(w), clima semicalido subhúmedo con lluvias en verano con temperatura media anual de 22 °C y precipitación de 698 mm (García, 1973).
Se utilizaron árboles injertados de zapote mamey de tres años y cuatro años de seis selecciones: 'El Regalo', 'Alfonso 15' y 'Nidito de Amor', procedentes de Guerrero, México; mientras que 'Magaña 1', 'Magaña 2' y 'Dany 66' de Yucatán, México. Establecidos a 68 m, el manejo agronómico que el productor da a la huerta, consistió en riegos ligeros cada tercer día con aproximadamente 40 L de agua por árbol, fertilización semanal (10 g planta-1) con la fórmula 18-46-00 o bien con 17-17-17.
Los árboles no fueron podados para conocer su vigor. Los deshierbes se realizaron en forma manual alrededor del árbol, pero cuando la maleza fue abundante se aplicó glifosato (Faena®) a dosis de 1.5 mL por litro de agua. Las variables evaluadas en los brotes y hojas de las selecciones de zapote mamey fueron las siguientes características.
Cinética de crecimiento foliar. Para conocer el tiempo que requieren las hojas desde que aparecen hasta que alcanzan el máximo crecimiento, en cinco árboles de la selección 'Nidito de Amor' se marcaron 10 brotes uniformes (30 a 40 cm de largo) y dos hojas por brote (n= 20 hojas). Debido que las hojas de mamey emergen en grupos a partir del ápice, no fue posible medirlas en los primeros cinco días debido a que se dañarían; por esta razón, se eligieron hojas que tenían de cinco a seis días de emergidas. Con una regla graduada en milímetros, las hojas fueron medidas semanalmente en largo y ancho. El ancho se midió en la parte media de la hoja. Esta variable se evaluó durante enero de 2009. Con los datos se graficó la cinética de crecimiento foliar hasta alcanzar el tamaño máximo.
Crecimiento de brotes. Se midieron mensualmente durante un año 7 brotes (35 a 60 cm), distribuidos en la periferia del dosel de cuatro árboles de cada selección. Los brotes se marcaron con listones para identificarlos fácilmente. Con un flexómetro se midió la longitud del brote (cm), desde la base hasta el ápice, el dato de crecimiento en cada mes se obtuvo restando el dato de cada mes, al del mes anterior. Con los datos se graficó el crecimiento de los brotes durante el año.
Peso específico de la hoja. Se midió en los meses de junio, agosto y octubre (2009), en 12 hojas completamente desarrolladas, sanas y maduras provenientes de cuatro árboles (3 hojas por árbol), de cada selección. Se envolvieron en papel húmedo, se colocaron en bolsas de plástico y se trasladaron al laboratorio. El área foliar se midió con un integrador LICOR-3100® (LICOR, Inc. Lincoln, NE, USA), y el peso de materia seca (PMS) se determinó después de secar por 48 h a 80 °C. Con los datos de área foliar y peso de materia seca, el PEH se calculó con la ecuación: PEH= peso seco (mg)/área foliar (cm2).
Grosor de la hoja. Se midieron 10 hojas completamente sanas, desarrolladas y maduras, en cada una de las selecciones evaluadas. Las hojas se lavaron con agua destilada, se realizó un corte en la parte media, de aproximadamente 1 cm2. Cada muestra se colocó en un micrótomo de congelación, se le agregó agua y se le aplicó CO2para solidificar la muestra. Posteriormente, el material se cortó con un micrótomo rotatorio de congelación (Spencer, Mod 820® American Optical Company; Massachusetts, USA).
En cada hoja se hicieron seis cortes, los cuales se colocaron en agua, se extendieron y se pusieron en portaobjetos; las mediciones de estos cortes se realizaron en la parte media, con un microscopio óptico (Carl Zeiss® SMT, Inc., New York, USA) con el objetivo de 10X y con un micrómetro ocular. El grosor se calculó con la fórmula: grosor de la hoja= unidad ocular*7.5µm; dónde: unidad ocular= valor observado en campo de 10X; 7.5 µm= factor para hacer los cálculos en campo de 10X.
Se realizaron análisis de varianza simple con base en un modelo completamente al azar. La comparación de medias se hizo con base a Tukey a nivel de significancia de α≤ 0.05. Los análisis se realizaron con el paquete estadístico SAS (Statistical Analisis System) versión 9.0 (SAS, 2002).
RESULTADOS Y DISCUSIÓN
Cinética de crecimiento foliar. En Alpoyeca, Guerrero, las hojas de zapote mamey alcanzan el tamaño máximo entre 35 y 42 días después de la emergencia. En la selección 'Nidito de Amor', a 7 días las hojas ya tenían 48% del tamaño final y a 28 días el 93% (Figura 1). El tamaño máximo se alcanzó a cuatro semanas, a partir de ese momento no presentó diferencias (p≤ 0.05). El promedio de largo de las hojas evaluadas fue de 15.4 cm y el ancho de 3.9 cm. Las ecuaciones polinómicas de tercer orden que relacionan el largo y ancho con la edad de la hoja presentaron alto valor de bondad de ajuste (R2≥ 0.99 para largo y R2≥ 0.97 para ancho).

Si consideramos lo indicado por Kennedy y Johnson (1981) en manzano y por Whiley (1990) en aguacate, se espera que las hojas de zapote mamey sean autosuficientes fotosintéticamente a 23 días cuando alcanzan 80% del tamaño. Es importante mencionar que esta variable se evaluó en enero, donde las temperaturas son menores que en la época de mayor crecimiento; por lo que, es posible que en otras épocas del año requiera de menos tiempo.
Crecimiento de brotes. El zapote mamey tiene crecimiento vegetativo principalmente en la parte terminal del brote, lugar donde se ubican las hojas, las cuales caen al final de cada ciclo en árboles adultos; por lo que, esta especie se considera caducifolia en condiciones de Alpoyeca, Guerrero. En la Figura 2 se observa que el crecimiento vegetativo de los árboles de tres años es constante durante el año. La selección 'Magaña 1' fue la que presentó mayor crecimiento (72 cm), seguida de 'Magaña 2' (65 cm) y finalmente 'El Regalo' con 62 cm. Los meses donde los brotes crecieron más fue de junio a septiembre para los tres materiales, esto coincide con la época de lluvias y con incremento de temperatura.

Como el crecimiento vegetativo no había sido evaluada en esta especie, se consideró la información de otros frutales. En mango, Avilan et al. (2000) observaron que el crecimiento de brotes es rítmico con tres o cuatro flujos vegetativos de 30 a 40 días de duración cada uno, en un año; pero esta característica puede ser modificada por la poda, ya que arboles podados, desarrollaron hasta cuatro flujos de crecimiento en nueve meses, mientras que los árboles sin poda presentaron de uno a dos flujos. En aguacate el crecimiento también ocurre en forma de flujos vegetativos y dependiendo de las condiciones climáticas, así como de la carga de fruto en el árbol, se pueden presentar de dos a cuatro flujos al año (Salazar-García et al., 2006).
Al igual que en las especies anteriores, en zapote mamey el crecimiento de brotes ocurre en dos flujos de crecimiento al año, presentándose: uno en junio-septiembre y el otro de diciembre a febrero y las plantas no se defolian. Sin embargo, Ibarra (2005) reportó que en Alpoyeca, Guerrero. El crecimiento vegetativo se presenta de enero a mayo una vez que los árboles renovaron sus hojas.
Las selecciones de 4 años, tuvieron comportamiento diferente, 'Nidito de Amor' creció más (44 cm), seguido de 'Alfonso 15' (43 cm) y finalmente 'Dany 66' (40 cm), estas selecciones detuvieron el crecimiento entre enero y marzo, posteriormente fue constante hasta diciembre (Figura 3). Esto se debió que el crecimiento de los brotes es afectado por otros órganos que demandan carbohidratos como los frutos, siendo los competidores más fuertes, al reducir el crecimiento vegetativo, tal como lo observaron Sanz et al. (1987) en hojas de naranjo 'Navel' donde ocurren cambios en elementos minerales y carbohidratos durante el crecimiento vegetativo.
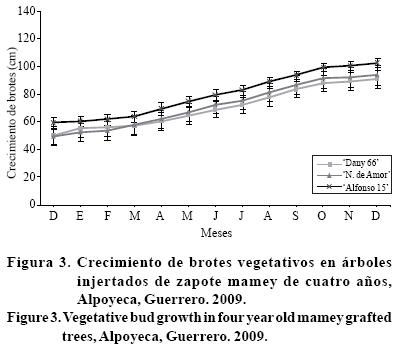
Otros factores, pueden ser las reservas de la planta, agua y nuevos fotoasimilados, además del ambiente, temperatura, luz y fotoperiodo (Sanz et al., 1987). Generalmente hay una etapa de crecimiento acelerado que después decae y finalmente cesa, por fenómenos correlativos como competencias e inhibiciones relacionadas con hojas, engrosamiento de tallo, brotes, tejidos de ramas, crecimiento de raíces y frutos o por condiciones ambientales desfavorables y acortamiento de los días (Forshey et al., 1987). Cabe indicar que en cítricos, durante la primavera se presenta floración y crecimiento vegetativo simultáneamente (Reyes et al., 2000).
En zapote mamey durante la primavera solo ocurre crecimiento vegetativo (Davenport y O'Neal, 2001), Cabe indicar que durante junio, julio y agosto, las plantas de zapote mamey presentan floración y tiene frutos del año anterior en desarrollo y desde septiembre hasta la cosecha, las plantas tienen frutos del año anterior (más de 14 meses) y frutos en desarrollo de 2 a 4 cm de diámetro, esta etapa fenológica en Alpoyeca, Guerrero, coincide con la detención del crecimiento y defoliación de las plantas, que dependiendo de la selección puede ocurrir desde diciembre hasta abril.
También se observó que los árboles de tres años presentaron mayor crecimiento de brotes que en árboles de cuatro años. En ambos casos se aprecia el efecto de la edad ya que Ibarra (2005), en árboles sin injertar de zapote mamey de 12 a 20 años, en Alpoyeca, Guerrero; encontró que el mayor crecimiento fue de enero a mayo, y puede ser de dos formas, el primero ocurre gradualmente mientras avanza la defoliación, y el segundo inicia hasta que la rama está completamente defoliada.
La longitud de brotes que observó fue de 3 cm hasta 22 cm durante seis meses de evaluación. Por otra parte, Granados (1995), en el Peten, Guatemala, observó longitudes entre 14 y 31.6 cm en la misma especie, esto puede deberse que las condiciones climáticas en donde se desarrollaron las plantas son más favorables para su crecimiento, ya que es uno de los sitios de origen del zapote mamey, mientras que Alpoyeca, Guerrero; se encuentra a mayor altitud (1 05 0 m) y latitud lo que puede afectar negativamente el desarrollo de las plantas.
El mayor crecimiento vegetativo en las plantas de tres años, se presentó en el periodo comprendido de junio a septiembre, mientras que las de 4 años en los meses de abril a agosto, lo cual puede estar relacionado con la floración que se presenta durante este mes. Damián-Nava et al. (2004) observaron en guayabo que la brotación de primavera (abril a junio) fue mínima, comparada con la de invierno que fue máxima. Además detectaron tres puntos importantes de brotación vegetativa en el año; dos en verano, y el mayor en invierno.
Las tasas más altas de alargamiento de brotes se registraron en septiembre (0.32 cm d-1) y de diciembre a febrero (0.28 cm d-1). Con base en lo anterior, para la evaluación de esta variable se deben considerar, edad de la planta, condiciones donde se desarrolla y manejo agronómico. En zapote mamey es importante que se evaluara en plantas injertadas con desarrollo en condiciones ambientales favorables, para definir número de flujos y tamaño de los mismos, dado que este aspecto es fundamental para el manejo de la especie.
Peso específico de la hoja. En el Cuadro 1 se observa que en el índice de área foliar (IAF), peso de materia fresca (PMF) y seca (PMS), 'El Regalo' fue superior (p≤ 0.01) a las demás selecciones. Para el PEH 'Magaña 2' fue igual a 'Magaña 1', 'El Regalo' y 'Alfonso 15' pero diferentes a 'Nidito de Amor' y 'Dany 66' (p≤ 0.01); el cual presentó el menor valor en todas las variables mencionadas. El IAF y PEH pueden ser modificadas por el portainjerto (Arrieta-Ramos et al., 2010; Berdeja-Arbeu et al., 2010) y por la época del año (Castillo-González et al., 1998), por lo que es conveniente conocer el comportamiento de la especie respecto a las etapas fenológicas, para poder realizar los muestreos en épocas más apropiadas.
Estos resultados difieren de los obtenidos por Ibarra (2005), en hojas de zapote mamey sin injertar de 12 a 20 años encontró PEH de 5.6 a 10.8 mg cm2, los cuales son inferiores a los obtenidos en esta investigación. Es importante señalar que Ibarra (2005) no menciona la época en que realizó las evaluaciones, debido que PEH es una variable que se modifica con la época del año, se debe evaluar de 3 a 4 veces para observar el comportamiento.
Reyes-Santamaría et al. (2000) en dos cultivares de naranjo ('Valencia' y 'Mars') observaron que ambas presentaron variación anual similar en el PEH, y que él mayor valor se observó en los meses de octubre a febrero, época en la cual la planta debió acumular mayor cantidad de carbohidratos para utilizarlos en la diferenciación floral, crecimiento vegetativo inicial y fructificación como lo sugieren, Marini y Sowers (1990) en manzano (Malus pumila Mill) y Castillo-González et al. (1998) en aguacate (Persea americana).
Grosor de hoja. En el Cuadro 2, se observa que 'El Regalo' fue igual (p≤ 0.01) a 'Magaña 2' y 'Magaña 1'pero diferentes a las demás selecciones, 'Dany 66' presentó el menor valor. En trabajos previos, en plantas de zapote mamey sin injertar, Ibarra (2005) encontró que el grosor de las hojas oscilaban entre 203.9 y 250.3 µm, en este caso el grosor fue de 249.6 a 291.6 µm, las diferencias pueden ser atribuidas a la edad de las plantas y al manejo, ya que los materiales utilizados en esta investigación fueron regados dos veces por semana y fertilizados con mayor frecuencia. Mientras que los utilizados por Ibarra (2005) se regaban cada 15 días y no eran fertilizados.
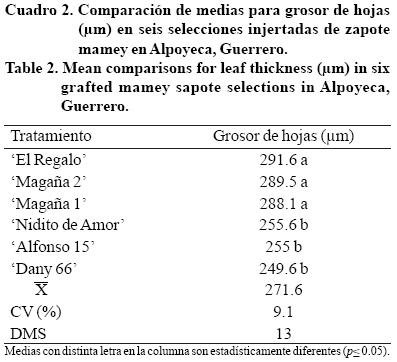
Reyes-Santamaría et al. (2000) mencionan que el grosor de la hoja varía entre especies (297.6 y 347.6 µm). Otro factor que modifica las características anatómicas de la hoja, es la injertacion, tal como lo observaron Ayala-Arreola et al. (2010) en aguacate, encontraron que el grosor de la epidermis superior e inferior y el parénquima en empalizada, se incrementó significativamente con el uso de portainjertos e interinjertos.
Si consideramos que el PEH puede relacionarse con el grosor de la hoja, debido al número de capas de células en empalizada y tamaño de estas (Marini y Sowers, 1990), entonces se puede decir que 'Dany 66' posee menor cantidad de tejido fotosintético por hoja, debido que es la selección que presentó menor grosor; en consecuencia, debería presentar menor tasa fotosintética, mientras que 'El Regalo' presenta el valor más alto, debería tener mayor tasa fotosintética, es necesario observar cómo se comportan estos materiales injertados en producción, ya que 'El Regalo' es un árbol muy productivo y 'Dany 66' ha tenido crecimiento lento y aun no entra en producción.
CONCLUSIONES
En Alpoyeca, Guerrero, las hojas de zapote mamey alcanzan el tamaño máximo a los 42 días; no obstante, después de los 23 días tienen más de 80% del tamaño y pueden ser autosuficientes fotosintéticamente. Los brotes de plantas juveniles tienen mayor crecimiento que las plantas en producción. El mayor crecimiento de brotes en plantas juveniles es de junio a septiembre, mientras que en plantas adultas es de abril a agosto. 'El Regalo' colectado en Alpoyeca, Guerrero; presentó los mejores valores de área foliar, peso de materia fresca y seca, así como grosor de hoja, 'Dany 66' de Yucatán, los valores menores. El PEH se modifica con la época del año, por lo que se debe evaluar de tres a cuatro veces para tener resultados confiables.
LITERATURA CITADA
Arrieta-Ramos, B. G.; Villegas-Monter, A.; Hernández-Bautisa, A.; Rodríguez-Mendoza, Ma.; Ruiz-Posadas, L. y García-Villanueva, E. 2010. Estomas y vigor de naranjo 'Valencia' injertado en portainjertos tolerantes al virus de la tristeza de los cítricos. Rev. Fitotec. Mex. 33:257-263. [ Links ]
Avilan, L.; Marín, C.; Rodríguez, M. y Ruíz, J. 2000. Comportamiento de los brotes de mango en plantas tratadas con diferentes intensidades de poda, paclobutrazol y nitrato de potasio. Agronomía Tropical. 50:347-360. [ Links ]
Ayala-Arreola, J.; Barrientos-Priego, A.; Colinas-León, M. T.; Sahagún-Castellanos, J. y Reyes-Alemán, J. C. 2010. Relaciones injerto-interinjerto y características anatómicas y fisiológicas de la hoja de cuatro genotipos de aguacate. Revista Chapingo. Serie Horticultura. 16:147-154. [ Links ]
Berdeja-Arbeu, R.; Villegas-Monter, A.; Ruíz-Posadas, L. M.; Sahagún-Castellanos, J. y Colinas-León, M. T. 2010. Interacción lima persa-portainjertos, efecto en características estomáticas de hoja y vigor de árboles. Revista Chapingo. Serie Horticultura. 16:91-97. [ Links ]
Bugarin, M. R.; Spínola, A. G.; García, P. S. y Paredes, D. G. 2002. Acumulación diaria de materia seca y de potasio en la biomasa aérea total de tomate. Terra. 20:401-409. [ Links ]
Castillo-González, A. M.; Colinas-León, M. T.; Ortega-Delgado, M. L.; Martínez-Garza, A. y Avitia-García, E. 1998. Variación estacional de carbohidratos en hojas e inflorescencias de aguacatero (Persea americana Mill.). Revista Chapingo. Serie Horticultura. 4:13-18. [ Links ]
Damián-Nava, A.; González-Hernández, V. A.; Sánchez-García, P.; Peña-Valdivia, C. B.; Livera-Muñoz, M. y Brito-Guadarrama, T. 2004. Crecimiento y fenología del guayabo (Psidium guajava L.) cv. 'Media China' en Iguala, Guerrero. Rev. Fitotec. Mex. 27:349-358. [ Links ]
Davenport, T. L. and O'Neal, J. 2001. Flowering and fruit development patterns of five mamey sapote cultivars in South Florida. Proc. Interamer. Soc. Trop. Hort. 44:56-59. [ Links ]
Figueiredo, J.; Sánchez, S.; Donadio, L.; Sobrinho, J.; Laranjeira, F.; Pio, M. y Sempionato, O. 2002. Porta-enxertos para a lima-ácida-Tahiti na regiao de Bededouro, sp. Revista Brasileira de Fruticultura. 24:155-159. [ Links ]
Forshey, C.; Weires, R. and Van-Kir, J. 1987. Seasonal development of the leaf canopy of 'Macspur Mcintosh' apple trees. HortScience. 20:881-883. [ Links ]
García, E. 1973. Modificaciones al sistema de clasificación climática de Köpen (para adaptarlo a las condiciones de la república mexicana). Instituto de Geografía. UNAM. D. F., México. 246 pp. [ Links ]
Granados, F. 1995. Algunas selecciones de zapote (Pouteria sapota) en Guatemala. Proc. Interamer. Soc. Trop. Hort. 39:115-118. [ Links ]
Horsley, S. B. and Gottschalk, K. 1993. Leaf area and net photosynthesis during development of Prunus serotina seedlings. Tree Physiology. 12:55-69. [ Links ]
Ibarra, E. M. E. 2005. Morfología de hojas y fenología en selecciones de zapote mamey (Pouteria sapota (Jacq.) H. E. Moore & Stearn) de Alpoyeca, Guerrero y Cazones Ver. Tesis de Maestría en Ciencias. Colegio de Postgraduados. Montecillo, Texcoco, Estado de México. 60 p. [ Links ]
Kennedy, R. A. and D. Johnson. 1981. Changes in photosynthetic characteristics during leaf development in apple. Photosynth. Res. 2:213-223. [ Links ]
Marini, R. P. and D. L. Sowers. 1990. Net photosynthesis, specific leaf weight and flowering of peach as influenced by shade. HortSience. 25:331-334. [ Links ]
Quijada, O.; Jiménez, O.; Matheus, M. y Monteverde, E. 2002. Evaluación de limero 'Tahití' sobre 10 portainjertos en la planicie de Maracaibo. Rev. Fac. Agron. 19:173-184. [ Links ]
Reyes-Santamaría, M.; Villegas-Monter, A.; Colinas-León, M. T. y Calderón-Zavala, G. 2000. Peso específico, contenido de proteína y de clorofila en hojas de naranjo y tangerino. Agrociencia. 34:49-55. [ Links ]
Salazar-García, S.; Cossio, L.; Lovatt, C.; González, J. and Pérez, M. H. 2006. Crop load affects vegetative growth flushes and shoot age influences irreversible commitment to flowering of 'Hass' avocado. HortScience. 41:1541-1546. [ Links ]
Sanz, A.; Monerri, C.; González-Ferrer, J. and Guardiola, J. L. 1987. Changes in carbohydrates and mineral elements in Citrus leaves during flowering and fruit set. Plant Physiol. 69:93-98. [ Links ]
Statistical Analysis System (SAS) Institute. 2002. SAS software release 9.0 SAS Institute Inc. Cary NC. USA. [ Links ]
Whiley, A. W. 1990. CO2 assimilation of developing shoots of cv. Hass avocado (Persea americana Mill.) a preliminary report. South African Avocado Growers Association. Yearbook. 13:28-30. [ Links ]

