










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.2 no.5 Texcoco sep./oct. 2011
Artículos
Anatomía de la lámina de hoja en ocho cultivares de aguacate*
Leaf lamina anatomy of eight avocado cultivars
Víctor Manuel González-Calderón1, Alejandro Facundo Barrientos-Priego2§, Carlos Alberto Núñez-Colín3, Sweetia Paulina Ramírez-Ramírez2, Reuben Hofshi4 y Mary Lu Arpaia5
1 Campo Experimental Valle del Fuerte. INIFAP. Carretera Internacional México-Nogales, km 1609. Los Mochis, Sinaloa. C. P. 81110. (gcvictorm@hotmail.com).
2 Instituto de Horticultura. Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5. Chapingo, Estado de México. C. P. 56230. (sweetia.ramirez@gmail.com).§Autor para correspondencia: abarrien@correo.chapingo.mx.
3 Campo Experimental Bajío. INIFAP. Celaya, Guanajuato. A. P. 112. C. P. 38110. (lit007a@gmail.com).
4 The Hofshi Foundation. 1260 S. Main St. Falbrook, CA 92028. USA. (rhofshi@avocadosource.com).
5 Department of Botany and Plant Sciences. University of California, Kearney Agricultural Center. 9240 S. Riverbend Avenue. Parlier, CA 93648. USA. (Arpaia@uckac.edu).
* Recibido: abril de 2011
Aceptado: octubre de 2011
Resumen
Actualmente en el mundo existen programas de mejoramiento genético en aguacate, para la obtención de nuevos cultivares y portainjertos, con el fin de incrementar productividad y problemas relacionados con estrés en el suelo. El caso de 'Lamb Hass' se ha encontrado que presenta características en hojas como mayor tasa de asimilación de CO2 y resistencia al ácaro persea. Estas características pueden estar relacionadas directamente con la anatomía de la hoja. Por lo que es importante conocer dichas características en actuales y nuevos cultivares: 'Hass', 'Reed', 'Gwen', 'GEM', 'Lamb Hass', 'Harvest', BL667 ('Nobel') y BL516 ('Marvel'). Durante 2007 en Chapingo, México, se realizaron cortes transversales de lámina foliar, donde se evaluaron características anatómicas. Se encontraron variables que no presentaron diferencias entre cultivares, que puede adjudicarse a que todos los genotipos utilizados en este estudio excepto 'Reed', provienen de un ancestro común 'Hass'. La agrupación de los cultivares es posible de acuerdo a características anatómicas de su lámina foliar, donde existe variación en las mismas que pueden interpretarse desde el aspecto fisiológico y sus semejanzas por su ancestría. 'Gwen', 'Hass', 'Marvel' y 'GEM', presentaron una cutícula más ancha y un menor ancho de células de parénquima en empalizada, mientras que 'Reed', 'Nobel' y 'Lamb Hass', presentaron características contrastantes con los primeros, 'Harvest' presentó valores intermedios. El cultivar 'Lamb Hass' presentó anatomía de la lámina foliar con características para llevar a cabo más eficientemente la fotosíntesis, debido a los mayores valores que presentó en su parénquima en empalizada I.
Palabras clave: Persea americana Mill., cutícula, epidermis, parénquima en empalizada.
Abstract
Currently in the world, there are breeding programs in avocado, aiming to obtain new cultivars and rootstocks to increase productivity and stress-related problems on the soil. In the case of 'Lamb Hass'; characteristics in leaves have been found such as increased CO2 assimilation rate and persea mite resistance. These features can be directly related to the leaf anatomy. It is important to know these features in current and new cultivars: 'Hass', 'Reed', 'Gwen', 'GEM', 'Lamb Hass', 'Harvest', BL667 ('Nobel') and BL516 ('Marvel'). During 2007 in Chapingo, Mexico, transverse sections of leaf blades were performed, evaluating anatomical features. There were found variables with no differences between cultivars, which may be because all genotypes used in this study except 'Reed', come from a common ancestor 'Hass'. The cultivars groupings are possible according to the anatomical characteristics of its leaf blade, where, there is a variation that can be interpreted from the physiological aspect and its likenesses by their ancestry. 'Gwen', 'Hass', 'Marvel' and 'GEM' had a wider cuticle and less wide palisade parenchyma cells, while 'Reed', 'Nobel' and 'Lamb Hass', had contrasting features, 'Harvest' had intermediate values. The 'Lamb Hass' cultivar presented a leaf blade anatomy with features that allow it to carry out photosynthesis more efficiently, due to the higher values of its palisade parenchyma I.
Key words: Persea americana Mill., cuticle, epidermis, palisade parenchyma.
INTRODUCCIÓN
La producción comercial del aguacate en México, ha sido afectada por el bajo rendimiento en comparación con otros países (República Dominicana, Israel y Sudáfrica). Existen varios factores que influyen en el rendimiento, entre los más importantes está el cultivar y el portainjerto a utilizar, así como su interacción (Ben-Ya'acov et al., 1993; Ben-Ya'acov y Michelson, 1995). Por lo que a nivel mundial se tienen programas de investigación para la obtención de nuevos cultivares y portainjertos, con el fin de solucionar este problema.
En el caso de cultivares existen varios programas de mejoramiento genético, donde han obtenido individuos con características particulares, siendo el programa de la Universidad de California Riverside, EE.UU. el más importante (Barrientos et al., 2000), el cual en los últimos 30 años ha liberado varios cultivares y puesto a prueba varias selecciones de aguacate con grandes expectativas (Kremer-Khöne, 1998; Arpaia, 2002).
La eficiencia fisiológica de nuevos cultivares de aguacate es de gran importancia, la cual puede estar relacionada con la anatomía de la hoja. Los espacios de aire de la hoja y los arreglos internos de las células, se asocian a la eficacia con la cual las plantas pueden asimilar el CO2 o intercambiar otros gases (Black et al., 1975).
Según Black et al. (1975) se ha demostrado con genotipos de Lolium, que el tamaño de la célula del mesófilo es inversamente proporcional a la asimilación de CO2; es decir, que la fotosíntesis aumenta mientras el tamaño de la célula disminuye. En el caso del manzano se ha probado que los mayores índices de potencial fotosintético están asociados a un mayor grosor de la hoja, peso específico y el número de capas de parénquima en empalizada (Barden, 1978). En manzano se puede observar que el diámetro de las células en el plano de la superficie de la hoja, es tres a cuatro veces más grande que las células en empalizada, pero el eje vertical de las células en empalizada es dos a tres veces más largo que el grosor de las células epidérmicas (Ryugo, 1988).
Se ha evaluado el porcentaje que ocupa el espacio intercelular en hojas de aguacate, ya que puede estar relacionado con la asimilación de CO2. Se encontró que dos capas de parénquima en empalizada conformaron aproximadamente dos tercios del grueso total de la hoja, y se determinó también que hay poco espacio intercelular en el parénquima esponjoso (Sharon et al., 2001). De acuerdo con Chartzoulakis et al. (2002), la anatomía foliar del aguacate es influenciada por condiciones ambientales, como la sequía, donde se observan grandes cambios, sobre todo en el espacio intercelular del parénquima esponjoso y en general en el grosor de la hoja.
Las diferentes cantidades de cera en la hoja de manzano, puede explicar parcialmente diferencias observadas en el intercambio gaseoso en distintos cultivares, debido a la variación en la limitación del movimiento de gases dentro y fuera de la hoja (Black et al., 1975; Mickelbart et al., 2000). Es de esperarse que la resistencia a algunos insectos y ácaros esté relacionada con el grosor de la cutícula y epidermis de las hojas, así como tricomas y ceras (Mickelbart et al., 2000).
Robbins et al. (1974), explicó que hay caracteres estructurales de la hoja que influyen en la cantidad de bióxido de carbono que llega a los cloroplastos. Estos factores incluyen tamaño, posición y comportamiento de estomas y la cantidad de espacio intercelular. También la intensidad y la calidad de la luz que alcanzan a los cloroplastos, son influidas por el grosor de la cutícula y la epidermis, por la presencia de pelos epidérmicos, la disposición de las células del mesófilo y la posición de los cloroplastos en las células.
En 'Lamb Hass' se ha encontrado que presenta algunas características únicas en sus hojas como son: mayor tasa de asimilación de CO2 y resistencia al ácaro persea (Hoddle, 1998). Estas características son de gran importancia para la selección de nuevos cultivares y pueden estar relacionadas directamente con la anatomía de las hojas, ya que en otras especies se ha encontrado que las dimensiones del mesóflo en empalizada de la hoja, está relacionada con su capacidad fotosintética (Wilson y Cooper, 1970); es de esperarse que la resistencia a algunos insectos y ácaros esté también relacionada con la cutícula y epidermis de sus hojas, así como tricomas y ceras (Mickelbart et al., 2000). Por lo anterior es importante conocer las características anatómicas de la hoja en nuevos cultivares de aguacate.
Por lo anterior se planteó como objetivo estudiar las características anatómicas de la lámina foliar de ocho cultivares de aguacate y agruparlos por dichas características.
MATERIALES Y MÉTODOS
Material vegetal
En el estudio se utilizaron los cultivares: 'Hass', 'Reed', 'Gwen', 'GEM', 'Lamb Hass', 'Harvest', 'Nobel' (BL667) y 'Marvel' (BL516). Los dos primeros cultivares fueron obtenidos por particulares y el resto son producto del programa de mejoramiento genético de la Universidad de California Riverside, EE.UU.
Toma de muestras
Las muestras se tomaron durante el mes de junio del año 2006, en un lote de evaluación localizado en el Rancho "ACW" en Fallbrook, California, EE.UU. Para lo cual se seleccionaron cuatro hojas completamente desarrolladas pero las más jóvenes de brotes completamente expuestos al sol, lo cual se realizó en tres árboles. Las hojas se cortaron y se utilizó un rectángulo de 1*3 cm de la parte media de la lámina, las cuales se fijaron en una solución FAA (50% etanol 96° + 5% ácido acético glacial + 10% formaldehído 40% de pureza + 35% agua).
Cortes anatómicos
Las muestras fijadas en FAA se procesaron durante 2007, con ayuda de en un cambiador automático de tejidos (Tissuematon Fisher) con cellosolve y xileno, para transferirse a parafina (55 °C) permaneciendo 72 h dentro de la estufa. Se elaboró el taquete y pirámide de parafina de acuerdo a la metodología propuesta por Sass (1968). En un micrótomo rotatorio (American Optical modelo 820) se realizaron cortes transversales en hoja abarcando la vena central con un grosor de 10 µm, los cuales se montaron con adhesivo Haup y formol al 10% y su tinción fue con safranina y verde fijo (Sass, 1968).
Imágenes digitales
A partir de las preparaciones anatómicas realizadas, se obtuvieron imágenes digitales con una resolución a 40x de objetivo y 10x de ocular en un microscopio marca Leica modelo Galen III, con adaptación para una cámara de vídeo marca Leica modelo ET-8800N conectado a una tarjeta de captura de imágenes GrabIT Pro para su posterior análisis (Figura 1 y 2).

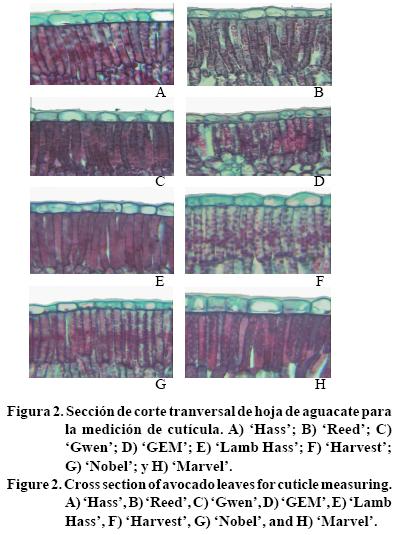
En cortes de lámina foliar se evaluó: grosor de la lámina; grosor de cutícula (Figura 2); grosor de la epidermis superior e inferior; grosor de parénquima en empalizadas I y II; grosor de parénquima esponjoso; área de espacio intercelular en parénquima esponjoso; relación longitud/diámetro de parénquima en empalizada I; diámetro, longitud, y área de células de parénquima en empalizada I.
Se realizaron cinco observaciones por repetición, para posteriormente utilizar el promedio de éstas en el análisis estadístico.
Análisis de imágenes
Las mediciones en las imágenes digitales se realizaron mediante el analizador de imágenes Image Tool ver. 3.00 (Wilcox et al., 2002).
Análisis estadístico
Se realizó un análisis discriminante canónico (ADC), para comparar los diferentes cultivares y determinar las variables que expliquen de mejor manera la variabilidad entre grupos formados, así como obtener la distancia de Mahalanobis y verificar si existen diferencias entre estos grupos (Jonhson, 1998). Adicionalmente se realizó un análisis de varianza entre cultivares y se llevaron a cabo pruebas de Tukey a una p< 0.05 para cada variable.
RESULTADOS Y DISCUSIÓN
Análisis discriminante canónico
Las tres primeras funciones canónicas aportaron 87.01%, de éstas la primera aportó 60.79% de la varianza, la segunda 15.45% y la tercera 10.77% (Cuadro 1).
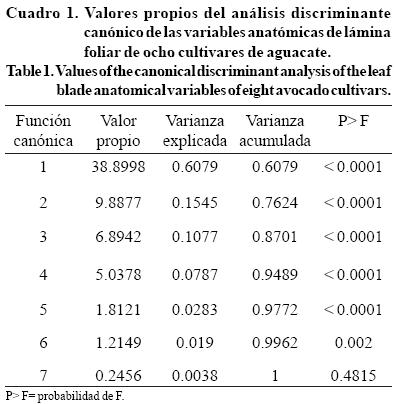
Se observó en la estructura canónica, que las variables más importantes de la primera función canónica (FC1) fueron la epidermis del haz y el espacio intercelular; y en el caso de las variables más importantes de la segunda función canónica (FC2) fueron la epidermis del haz, la cutícula y el ancho de célula de parénquima en empalizada y en la tercer función canónica (FC3) fueron epidermis del envés y la relación largo/ ancho de las células de parénquima en empalizada (Cuadro 2).
La primera función canónica fue definida por la epidermis del haz y el espacio intercelular, que se ha asociado con la eficiencia que se asimila el CO2 y el intercambio de otros gases (Black et al., 1975), además de la resistencia a algunos insectos y ácaros (Mickelbart et al., 2000).
La segunda función canónica estuvo definida por la epidermis del haz, la cutícula y ancho de la hoja, esta última se puede interpretar como índice de potencial fotosintético, ya que está muy relacionada con este proceso de acuerdo a Barden, (1978). Por otra parte puede explicar diferencias en el intercambio gaseoso (Black et al., 1975; Mickelbart et al, 2000), principalmente por diferencias en cutícula y epidermis.
La representación gráfica de los grupos en un plano factorial, dio como resultado la distribución de estos mismos en el espacio en las dos primeras funciones canónicas (FC1 y FC2), donde 'Nobel' y 'Reed' se encontraron distribuidos juntos (Figura 3), lo que indica que son genotipos anatómicamente muy similares de acuerdo a las variables evaluadas en este estudio, esto se debe posiblemente a que algún individuo de 'Reed' presentó el ancho de hoja mayor a los otros dos y muy semejante a 'Nobel'. 'Harvest' presentó el menor espacio intercelular, aunque fue similar a 'GEM', y 'Gwen', 'Nobel' y 'Reed' presentaron el mayor espacio intercelular. Por otra parte, 'Hass', 'Marvel' y 'Lamb Hass' se comportaron como intermedios.
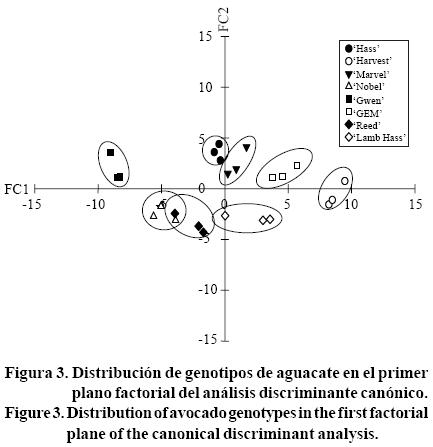
Se observó que el cultivar 'Gwen' presentó un grosor mayor de hoja y 'Harvest' presentó un menor grosor, de acuerdo a la primera función canónica (FC1). 'Gwen', 'Hass', 'Marvel' y 'GEM' presentaron un mayor grosor de cutícula y un menor ancho de células de parénquima en empalizada, mientras que 'Reed', 'Nobel' y 'Lamb Hass' presentaron características contrastantes con los primeros y 'Harvest' presentó valores intermedios. Los genotipos que presentaron mayor similitud fueron los cultivares 'Nobel' y 'Reed', así como 'Hass' y 'Marvel'.
Al calcular la distancia de Mahalanobis (Cuadro 3), entre los cultivares se obtuvieron las probabilidades con el fin de determinar la presencia de similitudes entre ellos. Se encontraron varios valores que no presentaron diferencia significativa, que se puede adjudicar a todos los genotipos utilizados en este estudio excepto 'Reed', provienen de un ancestro común que es 'Hass', pero a diferente nivel.
A continuación se explican las distancias menores obtenidas: el cultivar 'Reed' no presentó diferencias significativas con 'Nobel', esto se debió que un individuo de 'Reed' presentó el espesor de su hoja similar a 'Nobel', por lo que se puede considerar dentro del grupo.
El cultivar 'Reed' no presentó diferencias significativas con 'Lamb Hass', tal vez debido que un individuo de 'Lamb Hass' mostró un espesor de su hoja similar a 'Reed'; de tal manera, que puede estar considerado dentro de este grupo.
Los cultivares que presentaron mayor diferencia entre sí fueron 'Harvest' con 'Gwen', estas diferencias se observaron principalmente en el ancho de cutícula y en la relación largo/ ancho de células de parénquima en empalizada I.
Cabe indicar como se mencionó anteriormente, todos los cultivares, excepto 'Reed' están emparentados, por lo que sus semejanzas son fuertes como lo indicó la distancia de Mahalanobis (Cuadro 3). Donde 'Hass' es el ancestro directo de 'Gwen', y a partir de este último se derivó 'GEM', 'Harvest', 'Lamb Hass', 'Marvel' y 'Nobel'.
Análisis de varianza y prueba de medias
El análisis de varianza (Cuadro 4), indicó que en todas las variables existe un cultivar que difiere estadísticamente con otro.

Al comparar los datos obtenidos sobre 'Hass' en el estudio realizado por Chartzoulakis et al. (2002), con los obtenidos en el presente estudio en epidermis de haz y envés, parénquima en empalizada I y II y parénquima esponjoso, se pudo constató que en forma general son similares. Aunque hay diferencias en cuanto a grosor, donde las hojas de California, EE. UU, fueron más gruesas que las reportadas por Chartzoulakis et al. (2002) en Creta, Grecia; cuyas proporciones difirieron principalmente en la epidermis del envés (6.89 vs 4.24 µm: Grecia vs EE. UU).
El cultivar 'Hass' presentó la cutícula más gruesa y el menor grosor en la epidermis del envés (Cuadro 5), lo que puede indicar que para una mayor eficiencia de los productos que se asperjan al cultivar, es mejor que la aplicación sea dirigida al envés de la hoja (Beakbane, 1967).
En el espacio intercelular no existieron diferencias significativas entre los ocho cultivares (Cuadro 6), por lo que se podrá prescindir de esta variable en estudios posteriores, aunque el coeficiente de variación fue casi 27%, que explica el porqué no se encontraron diferencias y probablemente se requieran más repeticiones para encontrar diferencias.
'Harvest' presentó la cutícula más delgada, por lo que se puede considerar que tenga una mayor capacidad para el intercambio gaseoso (Black et al., 1975; Mickelbart et al., 2000). El cultivar Marvel tiene un mayor grosor en la epidermis del haz, por lo que puede presentar una mayor resistencia a plagas insectiles y algunos ácaros (Mickelbart et al., 2000), mientras que 'Nobel' tuvo un comportamiento contrario.
Se observó que el cultivar Reed, está dentro de las medias de todas las variables evaluadas, a pesar de provenir de ancestros distintos a los que provienen el resto de los cultivares. 'Lamb Hass' presentó en el parénquima en empalizada I, mayor largo de célula en empalizada, ancho de célula en empalizada y área de célula en empalizada, probablemente porque este cultivar es más eficiente fotosintéticamente (Wilson y Cooper, 1970), tal como lo indicó (Hoddle, 1998). El cultivar GEM presentó el menor valor en ancho de célula, que puede estar reflejado por la eficiencia fotosintética (Black et al., 1975).
CONCLUSIONES
La agrupación de los cultivares es posible de acuerdo a características anatómicas de su lámina foliar, donde existe variación en las mismas que pueden interpretarse desde el aspecto fisiológico y las semejanzas por los ancestros.
El cultivar Reed se ubicó dentro de las medias de todas las variables evaluadas, a pesar de provenir de cultivares distintos ('Nabal'*'Anaheim'), a los que provienen el resto de los cultivares (descendientes de 'Hass').
El cultivar 'Lamb Hass' realiza más eficientemente la fotosíntesis, ya que los valores que presenta en sus características del tejido el parénquima en empalizada I, son los de mayor valor.
LITERATURA CITADA
Arpaia, M. L. 2002. Enhancement of avocado productivity, plant improvement-selection and evaluation of improved varieties and roots tocks. In: Witney, G. (ed.). Proceedings of the California Avocado Research Symposium, 2002. Riverside California Avocado Commission. University of California Riverside. Riverside, USA. 43-53 pp. [ Links ]
Barden, J. A. 1978. Apple leaves their morphology and photosynthetic potential. HortScience. 13:644-645. [ Links ]
Barrientos-Priego, A. F.; Muñoz-Pérez, R. B.; Borys, M. W. y Martínez-Damián, M. T. 2000. Cultivares y portainjertos del aguacate. In: Téliz, D.; González, H.; Rodríguez, J. y Dromundo, R. (eds.). El aguacate y su manejo integrado. Mundi-Prensa S. A. de C. V. México. 35-54 pp. [ Links ]
Beakbane, A. B. 1967. A relationship between leaf structure and growth potential in apple. Ann. Appl. Biol. 60:67-76. [ Links ]
Ben-Ya'acov, A.; Michelson, E. and Sela, I. 1993. Rootstock effect on avocado vigor and productivity. Acta Hortic. 349:191-195. [ Links ]
Ben-Ya'acov, A. and Michelson, E. 1995. Avocado rootstocks. Horticultural Reviews. 17:381-429. [ Links ]
Black, C. C.; Goldstein, L. D.; Ray, T. B.; Kestler, D. P. and Mayne, B. C. 1975. The relationship of plant metabolism to internal leaf and cell morphology and the efficiency of CO2 assimilation. In: Burris, R. H. and Black, C. C. (eds.). CO2 Metabolism and plant productivity. University Park Press. Baltimore, USA. 113-139 pp. [ Links ]
Chartzoulakis, K.; Patakas, P.; Kofidis, G.; Bosabalidis, A. and Nastou, A. 2002. Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars. Scientia Hortic. 95:39-50. [ Links ]
Hoddle, M. S. 1998. Biology and management of the persea mite. California Avocado Society Yearbook. 82:75-85. [ Links ]
Johnson, D. E. 1998. Métodos multivariados aplicados al análisis de datos. International Thomson Editores. Distrito Federal, México. 566 p. [ Links ]
Kremer-Khöne, S. 1998. Maintenance and evaluation of avocado cultivars and selections. South African Avocado Growers' Association Yearbook. 21:35-37. [ Links ]
Mickelbart, M. V.; Miller, R.; Parry, S.; Arpaia, M. L. and Heath, R. 2000. Avocado leaf surface morphology. California Avocado Society Yearbook. 84:139-150. [ Links ]
Robbins, W. W.; Weier, T. E. y Stocking, C. R. 1974. Botánica. Editorial Limusa. D. F., México. 608 p. [ Links ]
Ryugo, K. 1988. Fruit culture, its science and art. John Wiley & Sons. New York, USA. 344 p. [ Links ]
Sass, J. E. 1968. Botanical microtechnique. The Iowa State University Press. Iowa, USA. 227 p. [ Links ]
Sharon, Y.; Bravdo, B. and Bar, N. 2001. Aspects of the water economy of avocado trees (Persea americana, cv. Hass). South African Avocado Growers' Association Yearbook. 24:55-59. [ Links ]
Wilcox, C., D.; Dove, S. B.; Doss, W. M. and Greer, D. B. 2002. UTHSCSA Image Tool. IT version 3.0. Department of Dental Diagnostic Science. University of Texas Health Science Center. San Antonio, USA. 57 p. [ Links ]
Wilson, D. and Cooper, J. P. 1970. Effect of selection for mesophyll cell size on growth and assimilation in Lolium perenne L. New Phytologist. 69:233-245. [ Links ]

