











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El maíz (Zea mays L.) es el cultivo agrícola más importante de México, por ser la fuente principal de alimentación humana y animal, así como en la elaboración de múltiples productos para la industria; se siembra en la mayoría de los estados en una amplia gama de condiciones climáticas (SIAP, 2017).
En el estado de Coahuila, en 2017 se sembraron 30,728 ha de maíz para grano, de las cuales, en el sureste del estado se sembraron 27,061 (88.1 %) y de estas, el 95.4 % fue bajo condiciones de temporal (SIAP, 2017), principalmente con poblaciones nativas. Las poblaciones nativas están estrechamente relacionadas con las condiciones del ambiente de producción y adaptación (temperatura, precipitación, altitud) y a los usos, principalmente del grano (Wellhausen et al., 1951; Ruiz et al., 2008). En el sureste del estado de Coahuila, se han identificado ocho grupos raciales de maíz: Celaya, Cónico Norteño, Elotes Cónicos, Elotes Occidentales, Olotillo, Ratón, Tuxpeño y Tuxpeño Norteño en una amplitud de altitudes entre los 887 a 2557 m (Wellhausen et al., 1951; Rincón et al., 2010; Rincón y Ruiz, 2015).
Las consecuencias del cambio climático conllevan modificaciones de las condiciones del ambiente, determinada por los cambios en la temperatura, ocurrencia errática de la precipitación, presencia de heladas y etapas prolongadas de sequía. En México, producción de maíz en áreas agrícolas de temporal está expuesta a un alto riesgo debido al incremento de las temperaturas y a la reducción de la precipitación como consecuencia del cambio climático (Hellin et al., 2014). En un análisis de los efectos del cambio climático, Ruiz et al. (2011), para el periodo 2051-2060 proyectan un incremento promedio en la temperatura de 1.9 ºC en las regiones productoras de maíz en México, lo que repercutirá en la disponibilidad del área cultivable, una reducción del ciclo de madurez en el cultivo y por lo tanto una reducción en el rendimiento de grano. Ureta et al. (2012) enfatizan en la influencia del cambio climático en los patrones de distribución de cultivos, particularmente en los sistemas de agricultura tradicional y perciben una reducción del 30 % en las áreas apropiadas para siembra de maíz.
Los efectos en la agricultura se manifiestan en bajos rendimientos en la producción de alimentos, así como en la erosión genética, causada por la pérdida de materiales genéticos debido a los periodos prolongados de condiciones que limitan la productividad. La presión ambiental por efectos del cambio climático sugiere el análisis de la diversidad genética y su interacción con las modificaciones ambientales, como alternativa para identificar alelos con potencial para mejorar poblaciones locales o para ampliar tolerancia a factores de estrés biótico y abióticos (Hellin et al., 2014). Mercer y Perales (2010) mencionan a la plasticidad (respuesta adaptativa a diferentes condiciones ambientales), la selección (variación intrínseca de alelos favorables) y el flujo genético (infiltración génica) como mecanismos de respuesta de las poblaciones nativas a los cambios ambientales. El conocimiento de la diversidad del maíz en un ámbito regional permite definir estrategias para su conservación, determinar su potencial genético con el propósito de ser incorporados a esquemas de selección per se o en combinaciones genéticas, pero también, la respuesta y adaptación a las condiciones del cambio climático. Debido a la importancia de las siembras de temporal, la diversidad genética de maíces existentes en el sureste de Coahuila, y la presión por efecto del cambio climático, se realizó el presente estudio con la finalidad de conocer el potencial de rendimiento y la respuesta a factores del ambiente de 63 poblaciones nativas de maíz representativas de la variación en el sureste de Coahuila. Lo anterior se basa en el supuesto de que la evaluación agronómica en ambientes representativos y contrastantes de la región de estudio permitirá determinar el potencial genético de poblaciones dentro de grupos raciales, que, de acuerdo con su área de adaptación, puedan ser incluidas como material base para el desarrollo de germoplasma capaz de responder a las modificaciones ambientales.
Método
El material genético estuvo constituido por 63 poblaciones nativas de maíz adaptadas al sureste de Coahuila, clasificadas en ocho grupos raciales (Cuadro 1).
Cuadro 1 Clasificación racial y área de adaptación de 63 poblaciones nativas de los maíces del Sureste de Coahuila.
| Clasificación racial † | Bajo (B) (< 1000) |
Intermedio (I) (1001 - 1800) |
Transición (T) (1801 - 2000) |
Altura (A) (> 2000) |
Total |
|---|---|---|---|---|---|
| Celaya | 3 | 3 | |||
| Cónico Norteño | 5 | 17 | 22 | ||
| Cónico Norteño _ Chalqueño | 1 | 1 | |||
| Cónico Norteño _ Elotes Cónicos | 2 | 2 | |||
| Cónico Norteño _ Ratón | 1 | 1 | |||
| Elotes Cónicos | 1 | 1 | 1 | 3 | |
| Elotes Cónicos _ Ratón | 1 | 1 | |||
| Elotes Occidentales | 1 | 1 | |||
| Olotillo | 1 | 1 | |||
| Olotillo _ Ratón | 2 | 2 | |||
| Ratón | 3 | 9 | 1 | 13 | |
| Ratón _ Elotes Cónicos | 1 | 1 | |||
| Ratón _ Tuxpeño | 2 | 2 | |||
| Tuxpeño | 3 | 3 | |||
| Tuxpeño Norteño | 3 | 1 | 4 | ||
| Tuxpeño _ Celaya | 2 | 2 | |||
| Tuxpeño _ Ratón | 1 | 1 | |||
| Total | 3 | 30 | 8 | 22 | 63 |
† El símbolo “_” entre los grupos raciales indica algún grado de combinación genética o formas intermedias entre razas.
Un grupo de las poblaciones evaluadas fueron recolectadas y descritas como parte del estudio y distribución de los maíces nativos de Coahuila (Rincón et al., 2010); el resto de las poblaciones fueron obtenidas y descritas en colectas posteriores (datos no publicados). Las poblaciones nativas provienen de áreas ecológicas del sureste de Coahuila, que oscilan entre 887 y 2557 m. Con propósitos de interpretación de la respuesta a los ambientes, las poblaciones fueron agrupadas de acuerdo con el área de adaptación: Bajo (≤ 1000 m), Intermedio (1001 - 1800 m), Transición (1801 - 2000 m) y Altura (> 2000 m), en referencia a la clasificación que se propone para la descripción varietal de maíz (SNICS, 2005), así como en grupos raciales de acuerdo con la clasificación propuesta por Wellhausen et al. (1951) (Cuadro 1). En el estudio se incluyeron siete genotipos como testigos, variables en madurez y tipo de grano: una variedad experimental (POBAM), dos variedades mejoradas (VAN210 y JAGUAN) y cuatro variedades sintéticas proporcionadas por el Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) (6221, 6222, Pool31 y Pool32).
La evaluación agronómica de las 63 poblaciones y los siete testigos se llevó a cabo en el ciclo agrícola primavera-verano de 2013, en dos localidades contrastantes y representativas de las condiciones ambientales de la región en el sureste del estado de Coahuila: El Mezquite, Galeana, N. L., localizada a 10 km de los límites de Arteaga, Coah. (25° 05’ 22’’ N; 100° 42’ 31’’ O; temperatura media anual de 16.3 ° C; precipitación anual 372.8 mm; altitud de 1910 m); y General Cepeda, Coah. (25° 23’ 00.91’’ N; 101° 27’ 15.3’’ O; temperatura media anual de 18.4 ° C; precipitación anual 414.6 mm; altitud de 1457 m), bajo condiciones de riego en las dos localidades de estudio.
Los genotipos fueron establecidos en un diseño experimental de bloques incompletos 7 × 10, con arreglo α-látice (0,1), con dos repeticiones (bloques) por localidad. La siembra en la localidad, El Mezquite fue el 11 de mayo de 2013, en donde se estableció cada bloque en área independiente dentro del mismo predio (MEZ1 y MEZ2, respectivamente); en General Cepeda, cada repetición fue establecida en diferente área y fecha de siembra; la primera, el 25 de Mayo y la segunda, el 15 de Junio de 2013 (GC1 y GC2, respectivamente). De esta manera, los bloques representan un muestreo de las condiciones del ambiente diferentes dentro de cada localidad y, por lo tanto, se consideran ambientes diferentes, particularmente en la localidad de General Cepeda, donde la primera fecha fue afectada por el periodo de la canícula. La unidad experimental consistió en dos surcos de 4 m de largo, con una distancia entre plantas de 0.2 m y distancia entre surcos de 0.85 m en General Cepeda, Coah., y 0.92 m en El Mezquite, Galeana, N. L.
La respuesta ambiental se analizó con base en el rendimiento de grano (t ha-1) al 15 % de humedad y los días a floración masculina. Se realizó un análisis de varianza de acuerdo con el diseño experimental (SAS Institute, 2004) para probar los efectos de los genotipos y la interacción con las dos localidades de evaluación.
Los genotipos y las localidades fueron considerados efectos fijos y el resto de los
efectos en el modelo, aleatorios. Se realizó la descomposición de la suma de
cuadrados para probar los efectos de las poblaciones nativas, los testigos y la
interacción de ambos con las localidades. Para identificar genotipos sobresalientes,
en cada una de las variables en estudio se calculó un valor de decisión determinado
por la media más uno y dos veces el error estándar de la media (
Resultados
En el análisis de varianza se encontró significancia (p ≤ 0.01) entre las localidades de evaluación sólo en los días a floración masculina (Cuadro 2).
Cuadro 2 Cuadrados medios del análisis de varianza del rendimiento de grano y días a floración de las poblaciones evaluadas en 2013.
| FV | GL | Rendimiento de grano (t ha-1) |
Floración masculina (d) |
||
|---|---|---|---|---|---|
| Localidades (Loc) | 1 | 673.546 | 44,201.16 | ** | |
| Bloques (Blo) / Loc | 2 | 116.889 | ** | 548.61 | ** |
| Blo incompletos / Loc × Blo | 24 | 2.267 | 12.17 | ** | |
| Genotipos (Gen) | 69 | 5.074 | ** | 178.73 | ** |
| Poblaciones (Pob) | 62 | 4.362 | ** | 164.42 | ** |
| Testigos (Tes) | 6 | 9.782 | ** | 289.77 | ** |
| Pob vs Tes | 1 | 15.742 | ** | 191.20 | ** |
| Loc × Gen | 69 | 3.251 | ** | 22.80 | ** |
| Pob × Loc | 62 | 3.378 | ** | 20.94 | ** |
| Tes ×Loc | 6 | 1.224 | 27.45 | ** | |
| (Pob vs Tes) × Loc | 1 | 6.989 | * | 101.86 | ** |
| Error | 113 | 1.752 | 6.05 | ||
| CV (%) | 21.819 | 3.04 |
* y **=Significativo al 0.05 y 0.01 niveles de probabilidad; FV = Fuentes de Variación; GL=Grados de Libertad.
Las dos localidades son contrastantes en clima y altitud, y también, son representativas de las condiciones ambientales del sureste del estado de Coahuila, consideradas para efectos del estudio en áreas de adaptación intermedia (General Cepeda, Coah.) y de transición (El Mezquite, Galeana, N. L.), respectivamente. Sin embargo, debido a las condiciones del establecimiento del experimento, como el establecimiento de los bloques en espacios independientes, y en el caso de la localidad de General Cepeda, las dos fechas de siembra, no se encontraron diferencias significativas en el rendimiento de grano. En los genotipos evaluados se encontró diferencias estadísticas (p ≤ 0.01) en los dos caracteres en estudio, lo que se explica por la amplia variación entre las poblaciones nativas (Cuadro 1), así como la variación entre los testigos. La interacción poblaciones × localidades (p ≤ 0.01), refleja el cambio relativo en la respuesta a las condiciones del ambiente de evaluación, además de la respuesta determinada por la procedencia de las poblaciones nativas asociadas a los diferentes grupos raciales en estudio. En la interacción testigos × localidades no se encontraron diferencias en el rendimiento de grano, en tanto que en floración masculina la diferencia fue significativa (p ≤ 0.01). Lo anterior se atribuye a que los testigos han sido expuestos a mejoramiento genético, y, por lo tanto, muestran capacidad de respuesta en el rendimiento de grano en las dos localidades, al ser comparados con las poblaciones nativas, aunque estos, son sensibles a las condiciones ambientales en el ciclo vegetativo y respuesta debido a la procedencia y clasificación racial (Cuadro 1).
Potencial de rendimiento
Con base en el promedio del rendimiento de grano a través de localidades, se identificaron las mejores 20 poblaciones con referencia a los genotipos usados como testigos (Cuadro 3).
Cuadro 3 Medias de rendimiento de grano y floración masculina de las 20 poblaciones superiores y los testigos en la evaluación agronómica realizada en 2013.
| Genotipos | Rendimiento de grano (t ha-1) | Floración masculina (d) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Promedio | MEZ† | GC | MEZ | GC | ||||||||
| Poblaciones‡ | ||||||||||||
| I13R | 8.495 | * | 10.417 | ** | 6.572 | * | 92 | 68 | ||||
| I52T_C | 8.308 | * | 11.317 | ** | 5.299 | 107 | ** | 78 | ** | |||
| I38T | 8.222 | * | 7.682 | 8.762 | ** | 103 | ** | 71 | ** | |||
| I40R_T | 8.190 | * | 9.455 | * | 6.925 | * | 106 | ** | 79 | ** | ||
| I54T_C | 7.817 | * | 10.757 | ** | 4.877 | 107 | ** | 75 | ** | |||
| I34TN | 7.785 | * | 8.533 | 7.038 | * | 100 | * | 82 | ** | |||
| I49T | 7.486 | * | 7.104 | 7.867 | ** | 103 | ** | 72 | ** | |||
| I45R | 7.388 | * | 9.381 | * | 5.395 | 99 | * | 69 | ||||
| T17EC | 7.349 | 8.104 | 6.594 | * | 94 | 71 | ** | |||||
| I39R_T | 7.059 | 8.077 | 6.042 | * | 112 | ** | 79 | ** | ||||
| I33EO | 6.915 | 9.619 | * | 4.211 | 94 | 72 | ** | |||||
| I18TN | 6.859 | 8.245 | 5.473 | 100 | * | 69 | ||||||
| I02R | 6.847 | 7.776 | 5.918 | 100 | * | 72 | ** | |||||
| I16T | 6.806 | 6.822 | 6.789 | * | 100 | * | 69 | |||||
| I23C | 6.663 | 8.040 | 5.285 | 97 | 71 | ** | ||||||
| I59TN | 6.565 | 7.529 | 5.600 | 112 | ** | 76 | ** | |||||
| T11R | 6.530 | 8.471 | 4.588 | 99 | * | 72 | ** | |||||
| B15R | 6.386 | 6.605 | 6.166 | * | 101 | * | 73 | ** | ||||
| I01O | 6.364 | 6.817 | 5.910 | 95 | 67 | |||||||
| A43CN_EC | 6.332 | 8.870 | * | 3.794 | 85 | 63 | ||||||
| Testigos | ||||||||||||
| Pool31 | 8.437 | * | 9.141 | * | 7.734 | ** | 105 | ** | 72 | ** | ||
| 6222 | 8.333 | * | 8.888 | * | 7.777 | ** | 108 | ** | 76 | ** | ||
| 6221 | 8.037 | * | 8.456 | 7.619 | ** | 112 | ** | 76 | ** | |||
| POBAM | 7.444 | * | 8.430 | 6.458 | * | 96 | 67 | |||||
| Pool32 | 7.400 | * | 9.218 | * | 5.582 | 99 | * | 71 | ** | |||
| JAGUAN | 6.154 | 7.737 | 4.570 | 91 | 65 | |||||||
| VAN210 | 4.159 | 5.840 | 2.478 | 79 | 58 | |||||||
| Media | 6.066 | 7.617 | 4.515 | 94 | 68 | |||||||
| Máximo | 8.495 | 11.317 | 8.762 | 112 | 82 | |||||||
| Mínimo | 3.685 | 3.891 | 1.526 | 79 | 58 | |||||||
| EE | 1.316 | 1.217 | 1.406 | 3.567 | 1.251 | |||||||
* y ** :=Valores superiores a la media más uno y dos veces el
error estándar (
De las 20 poblaciones, mostraron un potencial de rendimiento promedio sobresaliente con base en el valor superior a μ + EE, representativas principalmente de las razas Tuxpeño, Ratón y Tuxpeño Norteño, procedentes de áreas de adaptación intermedias (Cuadro 3), y cuatro de estas poblaciones I13R, I52T_C, I38T y I40R_T, con valores similares al rendimiento de los mejores testigos (Pool31, 6221 y 6222). También, resalta el comportamiento de las poblaciones I13R y I40R_T con rendimientos superiores en las dos localidades de evaluación.
En la localidad El Mezquite las ocho poblaciones nativas de maíz (I13R, I52T_C, I40R_T, I54T_C, I45R, I33EO, T08CN y I35R) mostraron rendimientos superiores al mejor testigo Pool32 (Cuadro 2); de éstas, las poblaciones I13R, I52T_C y I54T_C, con valores superiores al valor calculado de μ + 2EE. Con excepción de T08CN (población adaptada al área de transición, información referida en el Cuadro 1), el resto de las poblaciones, de acuerdo con el sitio de procedencia, están adaptadas a áreas intermedias. En la localidad de General Cepeda, las poblaciones I38T y I49T, de la raza Tuxpeño, adaptadas a áreas intermedias fueron superiores en rendimiento de grano al mejor testigo (variedad sintética 6222).
La diferencia promedio en los días a floración de las poblaciones adaptadas a las áreas de transición y altura, evaluadas en las dos localidades de estudio fue de 18 a 25 días, en tanto que las poblaciones procedentes de áreas intermedias, la diferencia en floración alcanza valores superiores a los 33 días para las poblaciones I38T, I54T_C, I39R_T, I59TN. Las poblaciones adaptadas a áreas de transición y altura (A43CN_EC, T08CN, A28CN y A03CN) de la raza Cónico Norteño, en la localidad de General Cepeda tuvieron días a floración similar al promedio, lo que puede considerarse como poblaciones con capacidad de tolerar temperaturas superiores en relación con su área de adaptación.
Respuesta ambiental
En la Fig. 1 se muestran las medias del rendimiento de grano en las dos localidades de evaluación de los dos grupos de poblaciones de mayor frecuencia en el estudio, las 22 poblaciones adaptadas a áreas de Altura (Fig. 1a) y 30 de áreas intermedias (Fig. 1b).
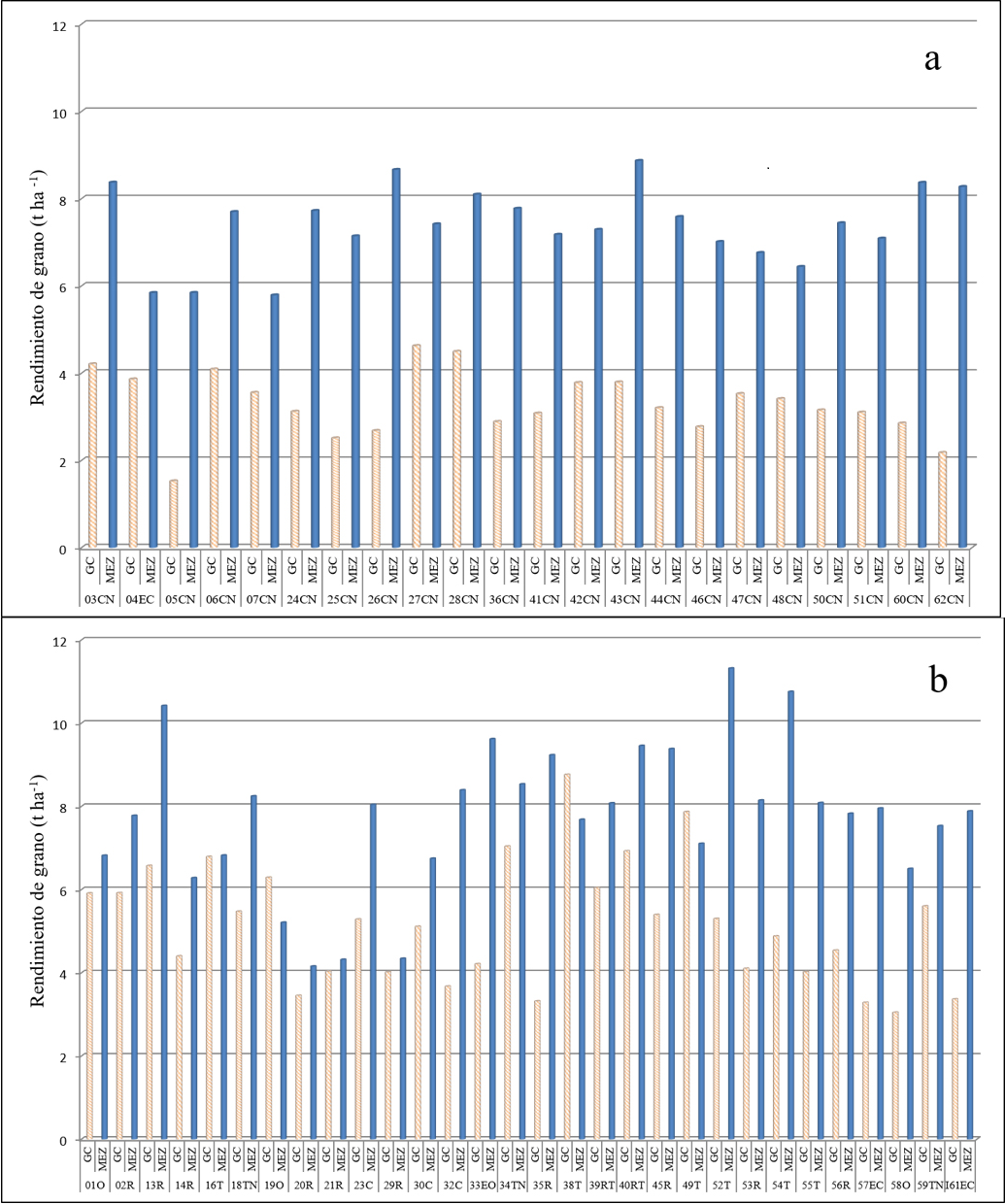
Fig. 1 Rendimiento de grano de poblaciones nativas de maíz evaluadas en dos localidades del sureste de Coahuila. GC=General Cepeda, Coah.; MEZ=El Mezquite, Galeana, N.L.; a) Poblaciones adaptadas a áreas de Altura (> 2000 m); b) Poblaciones adaptadas a áreas Intermedias (1001 - 1800 m); C=Celaya; CN=Cónico Norteño; EC=Elotes Cónicos; EO=Elotes Occidentales; O=Olotillo; R=Ratón; T=Tuxpeño; TN=Tuxpeño Norteño.
En el primer caso, los materiales de Altura (Fig. 1a), en general tuvieron un rendimiento promedio inferior en 4.1 t ha-1 (55.0 %), al ser evaluadas en la localidad de General Cepeda, con un rendimiento promedio de 3.29 y 7.39 t ha-1 para las localidades de General Cepeda y El Mezquite, Galeana, N. L., respectivamente. Con respecto al comportamiento de las 30 poblaciones de áreas intermedias (Fig. 1b), se encontró una respuesta en el rendimiento promedio de 2.6 t ha-1 (30.5 %) en la localidad El Mezquite, con medias de 5.15 y 7.75 t ha-1 para las localidades de General Cepeda, Coah. y El Mezquite, Galeana, N. L., respectivamente. Con excepción de las poblaciones 19O (Olotillo), 38T y 49T (Tuxpeño) con rendimientos superiores en General Cepeda, el resto de las poblaciones obtuvieron rendimientos superiores en la localidad El Mezquite, de las cuales 14 con rendimiento de grano superior a 8.0 t ha-1 (Fig. 1b) y ocho de estas, con rendimientos sobresalientes a través de los ambientes de evaluación (Cuadro 3).
Interacción genotipo por ambiente
La Fig. 2 presenta la dispersión gráfica de los genotipos (poblaciones nativas y testigos) de acuerdo con el modelo GGEbiplot para el análisis gráfico de la interacción genotipo × ambiente del rendimiento de grano en dos localidades (Yan et al., 2000).
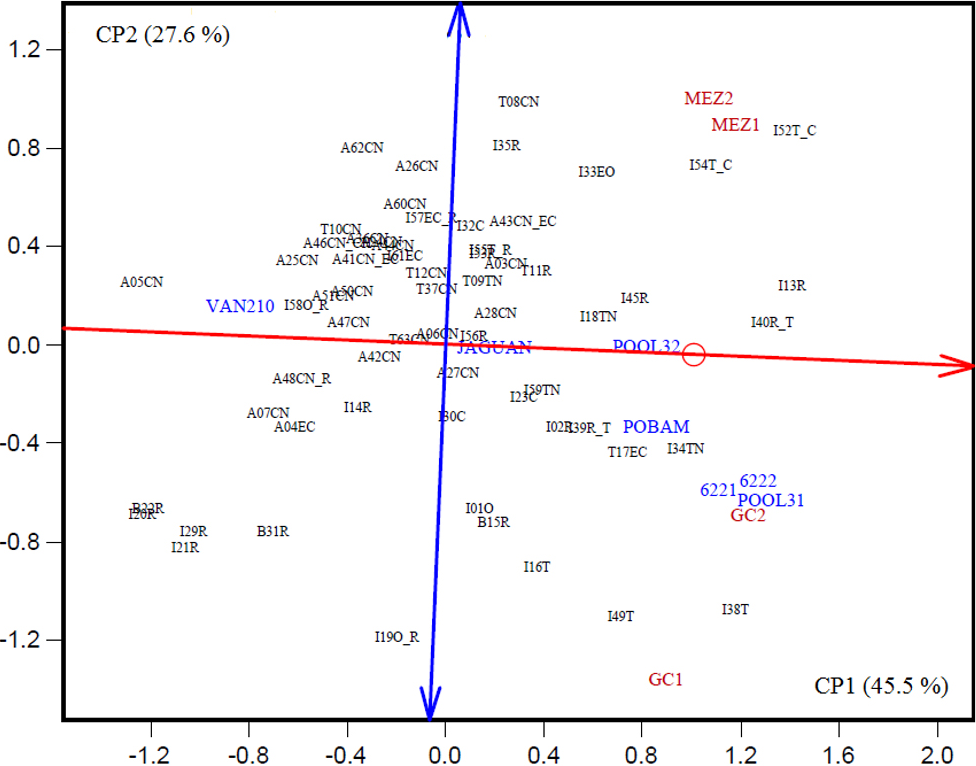
Fig. 2 Dispersión gráfica de la interacción genotipo × ambiente del rendimiento de grano en dos localidades (GC1, GC2, MEZ1 y MEZ2). El primer dígito en los genotipos indica el área de adaptación. A=Altura; B=Bajo; I=Intermedio; y T=Transición. Los dos dígitos siguientes indican el número de la entrada y el complemento, el grupo racial. C=Celaya; CN=Cónico Norteño; EC=Elotes Cónicos; EO=Elotes Occidentales; O=Olotillo; R=Ratón; T=Tuxpeño; TN=Tuxpeño Norteño.
El modelo GGEbiplot se basa en el análisis de componentes principales, que analiza la información de los efectos de los genotipos (G) y de la interacción genotipo × ambiente (GE), proporcionando un despliegue gráfico del comportamiento promedio y estabilidad de los genotipos de acuerdo con los ambientes de evaluación (Yan et al., 2000; Yan y Kang, 2003). En este análisis, los dos componentes principales (CP1 y CP2) explicaron el 73.1 % de la variación total acumulada de los genotipos y la interacción genotipo × ambiente del rendimiento de grano. De acuerdo con Yan y Kang (2003) si los ambientes de evaluación son los probadores de los genotipos, el eje de la abscisa indica los rendimientos promedio, en tanto que la ordenada al origen, valora la respuesta específica en los ambientes de prueba.
En la Fig. 2, el CP1 indica el comportamiento promedio en el rendimiento de grano de los genotipos, en tanto que, los valores positivos del CP2 y alejado del origen, se identifica las poblaciones con adaptación a la localidad El Mezquite, Galeana, N. L., las que se adaptan a General Cepeda se localizan con los valores negativos. Con el propósito de identificar a los genotipos con un comportamiento promedio a través de las variantes ambientales, se consideró a aquellos que se encuentran localizados cerca y paralelos a la abscisa (rango aproximado de -0.3 ≤ 0 ≤ 0.3 del CP2), lo cual señala como una forma de estabilidad de los genotipos en los ambientes de prueba (Yan y Tinker, 2006).
Las poblaciones con mayor rendimiento de grano a través de ambientes (Cuadro 3) se ubicaron con valores positivos del CP1 en la Fig. 2; de estas poblaciones, tres I52T_C, I54T_C y I33EO tuvieron un comportamiento y adaptación a la localidad El Mezquite, en tanto que, en la localidad de General Cepeda, Coah., se identificaron cinco poblaciones: I38T, I16T y I49T de la raza Tuxpeño; I34TN y T17EC de las razas Tuxpeño Norteño y Elotes Cónicos, y tres testigos (Pool31, 6221 y 6222). Por otro lado, se identificaron a nueve poblaciones con un comportamiento promedio en el rendimiento de grano a través de ambientes: I11R, I13R, I02R, I40R_T, I45R y I39R_T de la raza Ratón; I18TN y I59TN de Tuxpeño Norteño y I23C de Celaya, y dos testigos (Pool32 y POBAM).
Discusión o Conclusiones
Los experimentos fueron diseñados para establecer las repeticiones (bloques principales), siguiendo el principio de la experimentación en bloques, y en el caso de la localidad en General Cepeda, los dos bloques fueron separados por fechas de siembra. Lo anterior permitió que los bloques fueran diferentes, con un valor alto y significativo (p ≤ 0.01) del cuadrado medio de bloques dentro de localidad; valor utilizado para probar los efectos de localidades (Cuadro 2), que, bajo esta premisa, no se encontraron diferencias entre localidades, aun cuando son estas contrastantes en clima y altitud, además, representativas de las condiciones ambientales del área de estudio. Por lo tanto, las combinaciones de localidades × bloques (MEZ1, MEZ2, GC1 y GC2) fueron considerados ambientes diferentes para explorar e interpretar la interacción genotipo × ambiente mediante el análisis de dispersión gráfica, basado en el análisis de componentes principales (CP) con el modelo GGEbiplot, un diagrama de dispersión bi-espacial que permite visualizar las relaciones entre factores, en este caso, los genotipos y ambientes (Yan y Kang, 2003).
Las poblaciones que tuvieron un potencial de rendimiento promedio sobresaliente (Cuadro 3), son de las razas Tuxpeño, Ratón y Tuxpeño Norteño, procedentes de áreas de adaptación intermedias. En la evaluación de poblaciones nativas de maíz de Coahuila, Nájera et al. (2010), identificaron a las razas Ratón, Tuxpeño y Tuxpeño Norteño como las de mayor potencial de rendimiento. Al igual que en el presente estudio, diversos investigadores han identificado poblaciones nativas con potencial de rendimiento similares a las variedades mejoradas e híbridos comerciales (Pecina et al., 2011; Herrera-Cabrera et al., 2013); lo anterior destaca la importancia del germoplasma nativo y su capacidad de respuesta en ambientes contrastantes como resultado del proceso continuo de selección que realizan los agricultores (Hellin et al., 2014). El comportamiento promedio superior de las poblaciones de maíz de diferente grupo racial, así como la identificación de poblaciones con rendimiento aceptable en las dos localidades, determina el potencial genético, además, es posible identificar características adicionales para mejorar poblaciones locales en la tolerancia a factores adversos como la temperatura, estrés por sequía y adaptación, para mitigar las variaciones en los factores ambientales (Mercer y Perales, 2010; Hellin et al., 2014). De las 20 poblaciones con mayor rendimiento promedio de grano (Cuadro 3), 11 fueron las más tardías, con una diferencia promedio entre las dos localidades de 25 días, debido a las diferencias intrínsecas de las mismas, pero también, a la respuesta a las condiciones de los ambientes de evaluación, particularmente a la temperatura y fotoperiodo, factores donde se ha reportado correlación positiva en ambientes de Valles Altos (Giauffret et al., 2000).
En el caso de las poblaciones con adaptación a áreas de altura, aun cuando las poblaciones tuvieron una respuesta inferior en la localidad de General Cepeda, la reducción en el rendimiento tuvo una amplitud del 33.9 (04EC) al 73.9 % (05CN), lo que indica la respuesta diferencial entre las poblaciones, principalmente de la raza Cónico Norteño. Estos resultados muestran la vulnerabilidad de los materiales genéticos adaptados a Valles Altos a las condiciones extremas de temperatura como lo señalan Bellon et al. (2011). Las razas Cónico Norteño y Elotes Cónicos se encuentran adaptadas a las áreas de transición-altura en altitudes superiores a los 1700 m; en tanto que las razas Ratón, Tuxpeño y Tuxpeño Norteño su área de adaptación es muy amplia: Ratón (84 - 1300 m), Tuxpeño (0 - 1950 m) y Tuxpeño Norteño (1400 - 1701) (Ruiz et al., 2008). Lo anterior explica la respuesta en el rendimiento de grano de las poblaciones incluidas en el presente estudio (Figs. 1a y 1b) a los diferentes ambientes, donde destacan las poblaciones adaptadas a áreas intermedias como alternativa para identificar alelos favorables que ayudan a mitigar los efectos del cambio ambiental en materiales adaptados a Valles Altos, en selecciones per se o a través de combinaciones genética como lo señalan Mercer y Perales (2010). La respuesta ambiental de los dos grupos de poblaciones indica que los materiales genéticos adaptados a áreas de Altura (Fig. 1a) mostraron un abatimiento promedio de 55.0 % del potencial de rendimiento al ser expuestas a condiciones ambientales de la localidad intermedia, como el caso de la localidad de General Cepeda, Coah. Por otro lado, las poblaciones adaptadas a áreas intermedias (Fig. 1b), al ser expuestas a condiciones de la localidad de transición, como el Mezquite, Galeana N. L., muestran un comportamiento en general, favorable, con un abatimiento de 30.5 % al comparar la respuesta en las dos localidades de evaluación. Este patrón de respuesta coincide con resultados encontrados por diversos investigadores en evaluaciones de poblaciones nativas en ambientes contrastantes (Giauffret et al., 2000; Pecina et al., 2011).
Las poblaciones sobresalientes a través de ambientes en el rendimiento de grano (Cuadro 3, Fig. 2) corresponden principalmente a las razas Ratón, Tuxpeño y Tuxpeño Norteño, las cuales, con excepción de Tuxpeño Norteño, cuentan con una amplia adaptación (Ruiz et al., 2008). También se nota la variación entre las poblaciones de los diferentes grupos raciales como respuesta a la interacción con los ambientes de prueba, así como las poblaciones con estabilidad de rendimiento. Lo anterior puede explicarse por las combinaciones entre grupos raciales y adaptación específica (Cuadro 1), promovido por la forma tradicional de intercambio de semilla entre agricultores en la región de estudio (Aguirre et al., 2011; Rincón y Ruiz, 2015). Con base en la interpretación de la Fig. 2, se identificaron 22 poblaciones con adaptación a la localidad El Mezquite, que representa a las condiciones de transición-altura, 14 adaptadas a la localidad General Cepeda (condiciones de área intermedia) y 27 poblaciones con un comportamiento promedio (estable) a través de los ambientes. El concepto de estabilidad en esta interpretación se refiere a la respuesta promedio de los genotipos en cada ambiente, es decir, responden bien en ambientes favorables y viceversa (Weber et al., 1996).
Se identificaron ocho poblaciones de maíz con un potencial de rendimiento promedio sobresaliente a través de ambientes, principalmente de las razas Ratón, Tuxpeño y Tuxpeño Norteño. Las poblaciones adaptadas a áreas de Altura tuvieron un abatimiento promedio del 55.0 % en el rendimiento de grano al ser expuestas a condiciones de una localidad intermedia; en tanto que las poblaciones procedentes de áreas intermedias, la reducción en el rendimiento de grano, bajo las mismas condiciones fue de 30.5 %. La interpretación gráfica de la interacción genotipo × ambiente permitió identificar y agrupar a los genotipos con adaptación específica en las localidades de evaluación, así como aquellos con un comportamiento promedio a través de ambientes. Se identificaron y agruparon 22 poblaciones con adaptación a las condiciones de Altura, 14 a condiciones representativas de área intermedia y 27 poblaciones con un comportamiento promedio a través de ambientes.

