











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El nogal pecanero (Carya illinoensis) es una planta de la familia Juglandaceae (Hurrell et al., 2011, 47) nativo del sur de Estados Unidos y del norte de México (SAGARPA, 2002, 01). Se tienen registros de árboles que han llegado a medir 30 m y alcanzar una edad superior a los 100 años, con una producción individual de más de 100 kg de nuez por año (Medina-Morales, 2004, 446). Según el estudio de Zermeño-González et al., (2014, 24) el 97.39 % de la producción de nuez del país se obtiene en Chihuahua, Nuevo León, Durango y Coahuila.
En la Comarca Lagunera se estimó que en 2016 la superficie sembrada fue de 9,208 ha-1, con un rendimiento aproximado de 1, 459 t de nuez ha-1 (SIAP-SAGARPA, 2016); representando una derrama monetaria de 728 mil millones de pesos aproximadamente (SIAP-SAGARPA, 2016). Las variedades que predominan en los huertos de la Comarca Lagunera son Wichita, presente en un 62 % y la variedad Western que se encuentra presente en el 100 % de las huertas (SAGARPA, 2002, 02).
Las plantas requieren gran cantidad de nutrientes y energía para mantener sus funciones principales (crecimiento, respiración, reproducción y protección) y en diversas ocasiones estas pueden llegar a ocurrir de manera simultánea (Uscola et al., 2015, 814). El nitrógeno (N) es uno de los elementos que tiene más demanda por las plantas, ya que es un elemento nutritivo muy importante para el crecimiento (Frak, 2002, 1043) y el desarrollo (El Zein et al., 2011, 1390). La asimilación del N desde el suelo se ve afectada principalmente por la etapa de desarrollo de la planta (Dawson et al., 2008, 90; Xu et al., 2012, 158) y su falta es un factor limitante para el crecimiento y desarrollo (Ferlian et al., 2017, 17; Frak, 2002, 1043). Cuando la cantidad de elementos nutritivos no pueden ser satisfechos por la absorción, los arboles dependen directamente de los compuestos almacenados en forma de reservas (como proteínas, aminoácidos y nitratos en el caso de compuestos nitrogenados) (Villar-Salvador et al., 2015, 814).
Existen compuestos que se pueden almacenar en todos los órganos de la planta o en órganos específicos y se movilizan para apoyar el crecimiento y el funcionamiento (Face, 2013, 1096). En los arboles caducifolios existen compuestos que se almacenan anualmente durante la caída de las hojas y se movilizan en primavera (Bazot et al., 2013, 1024; Gilson et al., 2014, 117; Rennenberg et al., 2010, 276).
Los estudios realizados sobre las reservas de N se han llevado a cabo en plantas herbáceas anuales (Valenzuela-Núñez et al, 2010, 76), mientras las investigaciones en especies leñosas son escasas, resaltando Populus spp. (Cooke y Weih, 2005, 20), Hevea brasilensis (Tian et al., 1998, 153), Swietenia macrophylla (Tian et al., 2003, 492), Picea sitchensis (Proe y Millard, 1994, 76), Prunus avium (Millard et al., 2006, 528), Quercus robur (Valenzuela-Núñez et al., 2011, 142) y Pyrus pyrifolia (Marafon et al., 2016, 1137), sin embargo, la mayoría de los trabajos se han realizado en condiciones controladas (Millard y Grelet, 2010, 1083).
Los estudios que abordan el almacenamiento de compuestos nitrogenados en el tallo (El Zein et al., 2011, 1391) y la movilización de nutrientes hacia los tejidos de nuevo crecimiento son escasos (Villar-Salvador et al., 2015, 819). En múltiples ocasiones los elementos nutritivos satisfacen la demanda de las plantas, ya sea por factores ambientales o por su disponibilidad en el medio ambiente (Millard y Grelet, 2010, 1091). La longevidad de los árboles no permite una adaptación rápida a los cambios ambientales, propiciando que sean susceptibles a los efectos del cambio climático (Sellin et al., 2017, 2136; Zwieniecki y Secchi, 2015, 4).
Los cambios ambientales pueden aumentar o retrasar el crecimiento de los árboles (Sellin et al., 2017, 2136). La baja humedad relativa puede causar un descenso en la productividad y la biomasa de los árboles (Lendzion y Leuschner 2008,649; Sellin et al., 2017, 2143), de igual forma la radiación puede reducir el crecimiento de las plantas (Dengel et al., 2009, 548) y se ha comprobado que las temperaturas altas influyen sobre el N en los árboles (Coder, 2016, 03). Debido a que el nitrógeno es un elemento nutritivo que se almacena en los órganos perenes durante el otoño e invierno y se moviliza durante la primavera hacia el nuevo crecimiento, se pretende tener conocimiento de la influencia de las variables ambientales (temperatura, irradiación y humedad relativa) sobre el porcentaje de nitrógeno almacenado en la biomasa de las dos variedades cultivadas en la zona de estudio (Western o Wichita) con la finalidad de contribuir al manejo agronómico. Por lo anterior, el objetivo del presente trabajo fue determinar la variación del contenido de N en nogal durante un ciclo anual productivo de las variedades Wichita y Western y desarrollar una función matemática que describa el comportamiento del contenido de N con respecto a variables ambientales (temperatura, humedad relativa, irradiación).
Método
Área de estudio
El estudio se realizó en el campo experimental de la Universidad Autónoma Agraria Antonio Narro Unidad Laguna en Torreón, México en las coordenadas 25º 33’22.63’’ LN y 103º 22’07.77’’ LW. El clima del área de estudio es muy seco semiárido (BWh) (Briceño-Contreras et al., 2018, 4163), con una precipitación promedio anual de 189.6 mm, una temperatura media anual de 22.64 ºC (SAGARPA-INIFAP, 2017) y con una altitud media de 1,120 m.
Muestreo
El muestreo realizado fue sistemático para cada una de las variedades (Western y Wichita), de acuerdo a la metodología desarrollada por (Briceño-Contreras et al., 2018, 4164; Espino-Castillo et al., 2018, 311; Valenzuela-Núñez et al., 2010, 76; Valenzuela-Núñez et al., 2011, 142; Valenzuela Núñez et al., 2014, 35). Cuatro árboles jóvenes de una edad aproximada de 30 años fueron seleccionados de manera intercalada en el centro de la huerta, para minimizar el efecto borde. Se extrajeron muestras de manera mensual durante un ciclo anual productivo desde julio de 2016 hasta junio de 2017. Para cada uno de los árboles se extrajo una viruta en el tallo a una altura de 1.3 m con la ayuda de un taladro Pressler (Haglof BS072 ®). Las muestras se introdujeron en una hielera para ser transportadas al laboratorio siguiendo la metodología de Briceño-Contreras et al. (2018, 4164).
Procesamiento de las muestras
Las muestras fueron procesadas en el Laboratorio de Biología y Ecología Forestal en la Facultad de Ciencias Biológicas de la Universidad Juárez del Estado de Durango; se introdujeron y se etiquetaron en bolsas de papel aluminio previamente perforadas para su conservación. Las muestras se congelaron con N líquido, después se almacenaron en un ultracongelador (Revco Value Plus® Thermo Scientific®) a una temperatura de -70 °C, para posteriormente someterlas a un proceso de liofilización (Liofilizador Labconco® Freezone Triad Freeze Dry Systems®) durante siete días a -40 °C con el propósito de deshidratar las muestras y evitar toda actividad enzimática. Una vez liofilizadas las muestras fueron molidas en un equipo Pulverisette 15 Fritsch® para obtener un polvo fino.
Determinación de N total
La determinación de N total se realizó en el Laboratorio de Genética INRA-URP3F Génétique en el Institut National de la Recherche Agronomique (INRA) del Centre de Recherche Nouvelle-Aquitaine- Poitiers, Francia y se determinó la cantidad de N total expresado en kg y en porcentaje en la muestra por medio de un analizador elemental de gases (Thermo Scientific® Fash 2000 Organic Elemental Analizer ®).
Estimación de la biomasa del tallo del árbol
Para calcular el volumen de madera de cada uno de los tallos de los arboles fue necesario utilizar los valores de densidad de la madera determinados para nogal por el (INTI-CITEMA, 2003, 8) además del uso de ecuaciones alométricas en Carya según (Rodríguez-Laguna et al., 2006, 180). La biomasa del tallo se determinó mediante la siguiente ecuación:
Una vez calculado el contenido de N (kg) en el tallo, se utilizó una regla de tres simple para calcular el porcentaje de nitrógeno contenido en el tallo del nogal. La radiación, la humedad relativa y la temperatura media mensual fueron obtenidas de la base de datos del INIFAP campo experimental La Laguna (INIFAP, 2018).
Análisis estadístico
Se realizó un análisis de varianza de dos factores (variedad, mes e interacción) para observar la influencia sobre el porcentaje de N en el tallo del árbol; asimismo se realizó un análisis de regresión lineal múltiple para observar la influencia de las variables ambientales (temperatura, humedad relativa e irradiación) sobre el porcentaje de N en el tallo del árbol. Una vez determinada la variable que más influyó sobre el porcentaje de N se realizó un análisis de regresión lineal simple para observar su comportamiento con respecto al porcentaje de N en el tallo de nogal. Los procedimientos fueron realizados con la ayuda del software SPSS 18, con un nivel de significancia de 0.05.
Resultados
Las variedades (Wichita y Western) de nogal (F=0.846, g.l.= 1,72, P = 0.361), así como los meses del año (F=0.682, g.l.= 11, 72, P= 0.751) y la interacción (mes y variedad) (F=0.273, g.l. = 11, 72, P=0.989) (Figura 1) no influyeron significativamente en el porcentaje de N en el tallo de los árboles. La variedad Wichita estadísticamente fue igual a la variedad Western.
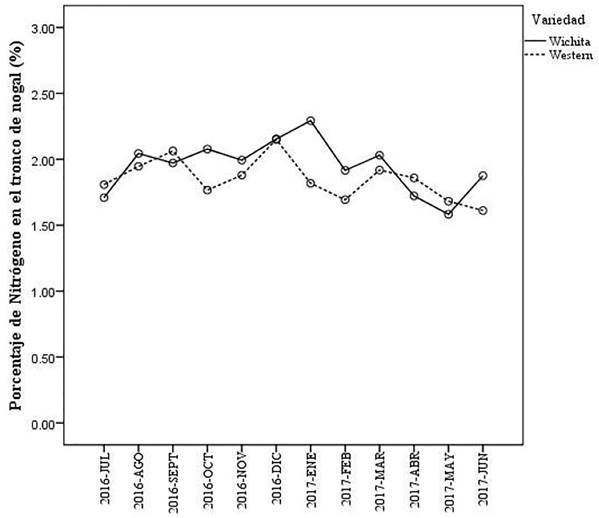
Figura 1 Porcentaje mensual de N en las variedades de nogal Wichita y Western durante un ciclo anual productivo en Torreón, Coahuila
El resultado del análisis de regresión lineal múltiple de la variedad Western indicó que la temperatura, la irradiación y la humedad relativa no predicen de manera significativa la acumulación de N en el tallo del árbol (F = 1.99, g.l. = 3,8, P = 0.193, R2= 0.42).
El resultado del análisis de regresión lineal múltiple de la variedad Wichita indicó que la temperatura, la irradiación y la humedad relativa predicen de manera significativa la acumulación de N en el tallo del árbol para la variedad Wichita (F = 5.81, g.l. = 3,8, P = 0.02, R2= 0.68). La ecuación de regresión obtenida para la acumulación del porcentaje N fue la siguiente, siendo la temperatura media mensual la variable que más influyó en el porcentaje de N en el tallo de nogal de esta variedad.
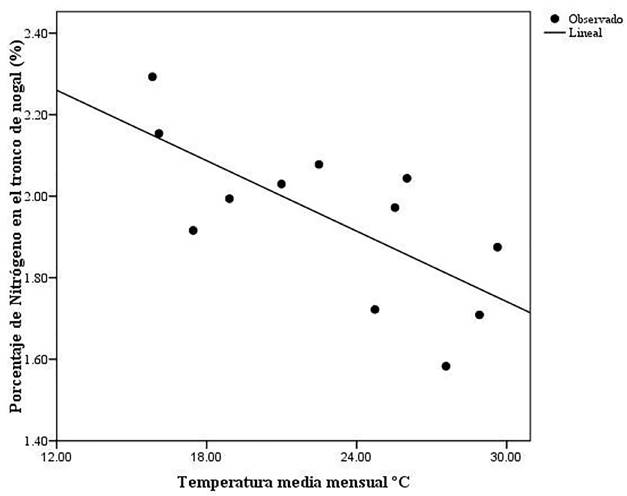
El resultado del análisis de regresión simple indicó que la temperatura predijo de manera significativa la acumulación de N en el tallo del árbol en la variedad Wichita (F = 10.07, g.l. = 1,10, P = 0.010, R2= 0.50). La ecuación de la regresión obtenida fue la siguiente:
Se observó que la relación entre la temperatura y el porcentaje de N fue inversamente proporcional, ya que si la temperatura es mayor el porcentaje de N en el tallo tiende a decrecer, y si la temperatura es menor el porcentaje de N aumenta como se puede observar en la Figura 2.
Discusión o Conclusiones
Los resultados obtenidos no concuerdan con lo observado por Muñoz et al. (1993, 268) ya que reportaron un aumento en el N en los tejidos desde agosto a noviembre en Prunus persica. Además Jordan et al., (2014, 61) encontró que en Prunus persica el N aumenta a finales de otoño. En las especies leñosas caducifolias, la reabsorción de N normalmente tiene lugar en otoño, ya que el follaje envejece en respuesta a las bajas temperaturas o a la disminución del fotoperiodo (Yasumura et al., 2005, 29). Sin embargo Cheng y Fuchigami, (2002, 1299) y Jordan et al., (2014, 62) reportaron que las observaciones en el contenido de N pueden alterarse debido a la movilización de este elemento, antes de la caída de las hojas hacia los órganos perenes.
Los resultados en la variedad Western son similares a lo reportado por Northup et al., (2005, 35) ya que presentó contenidos muy similares en C. pallida, C. taxensis, E. texana y S. cuneifolia (1.69 %. 1.37 %, 1.27 % y 1.29 % respectivamente). Los resultados en la variedad Wichita coinciden con lo reportado con Northup et al. (2005, 35) en G. angustifolia donde se encontró que los porcentajes de N fueron de 2.70 %.
El presente estudio difiere con lo reportado por Gérant (2017, 793) en Eucalyptus urophylla S.T. Blake en el que se comparó el porcentaje de N en la materia seca a diferentes alturas del tallo en septiembre y en abril, observándose un ligero aumento en el porcentaje de N en septiembre. Asimismo, los resultados de este estudio de las dos variedades concuerdan con lo reportado por Jordan et al. (2014, 68) en árboles de Prunus persica para el mes de mayo donde se reportó 1.39 % de contenido de N.
Por otra parte el porcentaje de N en las dos variedades (Western y Wichita) coincide con los reportados por Windels et al., (2003, 529), en los tallos de Acacia berlandieri (1.8 a 6.0 %). El porcentaje de N en el tallo en las dos variedades es superior a lo reportado por Gérant et al., (2017, 793) en Eucalyptus urophylla S.T. y Northup et al. (2005, 35) en el tallo de Acacia farnesiana. Los resultados de este estudio también concuerdan con lo reportado por Ledgard y Smith (1992, 62) en kiwi (Actnidia deliciosa) ya que no se reportaron cambios en el porcentaje de N (p>0.05) en el tallo.
Los compuestos almacenados tienen tendencia a fluctuar a corto plazo (horas-días) porque muestran plasticidad en la planta y varían conforme a los factores ambientales (Villar-Salvador et al., 2015, 815). El N se ve afectado por el frío y la sequía, pero no existe un consenso claro sobre la magnitud o la dirección de su efecto (Villar-Salvador et al., 2013, 222).
Sin embargo Zhu et al., (2001, 250) reportan que un alto contenido de N puede reducir la tolerancia a las heladas de las plántulas o promover la resistencia al frío. La baja concentración de N en los tejidos generalmente dificulta la aclimatación en frío (Taulavuori et al., 2014, 135).
En el presente estudio se puede observar que el porcentaje de N es mayor cuando la temperatura es baja. Taulavuori et al., (2014, 134) realizaron una revisión de los estudios publicados de 1990 hasta el 2014 y reportaron que en 40 % de los casos un alto contenido de N aumenta la resistencia a heladas, también comprobando además que cuando existe un mayor contenido de N en los tejidos también aumenta la tolerancia a bajas temperaturas. Villar-Salvador et al., (2015, 823) establecen que la movilización de los recursos almacenados muestra plasticidad y varía según los factores ambientales y endógenos en las plantas. Gérant et al. (2017, 796) asociaron las fuertes fluctuaciones de N a las estaciones secas y lluviosas. Lendzion y Leuschner (2008, 654) señalan que una combinación de precipitación, veranos calurosos, heladas tardías, frialdad extrema en el invierno y un corto período de vegetación probablemente influyen en el contenido de N, aunque los estudios sobre los efectos de alta temperatura en el crecimiento de árboles de hoja caduca han reportado que el calentamiento promueve el crecimiento (Way y Oren, 2010, 669).
En el presente estudio se observa una tendencia de disminución en el porcentaje de N contenido en el tallo del árbol cuando aumenta la temperatura. Coder (2016, 03) reportó que las temperaturas altas pueden dañar e inducir la muerte de los tejidos en los árboles. El estrés por calor afecta la función de la planta desde la escala celular hasta la planta completa (Marias et al., 2017, 302). El aumento de la temperatura actúa directamente sobre la membrana celular y deshidrata el tejido, dentro de los meristemos (puntos o áreas de crecimiento), la división y expansión celular se inhiben y la regulación del crecimiento se interrumpe (Coder, 2016, 04). En el estudio realizado por Way y Oren (2010, 6787) en Quercus rubra también se observó un efecto negativo de la temperatura elevada sobre el crecimiento lineal y sustancial, lo que concuerda con los resultados obtenidos en el presente estudio.
McClaran et al., (2013, 39) utilizaron ecuaciones para estimar el contenido de N en Prosopis velutina, obteniendo una correlación positiva con una R2 = 0.90 utilizando una función de potencia, lo que difiere en el presente estudio ya que una vez realizada la regresión lineal múltiple se identificó la variable que más influía en el porcentaje de N en el tallo (temperatura); la ecuación que se ajustó de mejor manera a los resultados fue de tipo lineal para la variedad Wichita con una R2 =0.50 para esta variedad. Northup et al. (2005, 29) encontraron que las predicciones de biomasa de tallos grandes son menos precisas que la de tallos pequeños (x>2.5), y se encontraron una baja relación, (R2= 0.32) no significativa en Z. fárgara (por debajo de los valores reportados en el presente estudio).
La respuesta de los árboles a la sequía depende de muchos factores ambientales y relacionados con el sitio, como la precipitación, la temperatura, la topografía y las características del suelo (Lévesque et al., 2014, 02; Weber et al., 2013, 179). Los resultados del presente estudio así como el de Wertin et al. (2011, 1287) indican que las plantas pueden responder de manera positiva o negativa a la temperaturas, además de que esto puede ser atribuido a la ubicación del sitio de crecimiento, dentro del rango geográfico de una especie en particular. El N es un elemento muy importante para las plantas, pero así como toda reserva vegetativa tiende a fluctuar con respecto a diferentes variables ambientales.

