











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El comportamiento productivo del ganado bovino en regiones tropicales generalmente es menor que en zonas templadas, debido parcialmente a las limitantes ambientales, como la menor calidad del alimento y la presencia de estrés por calor y humedad, lo que ocasiona mayor gasto de energía en el animal para el intercambio de calor hacia el exterior y la reducción de la ingesta de alimento (Finch, 1986). Los desafíos de varios de los sistemas de producción animal en las áreas tropicales, no sólo consisten en incrementar la producción de los animales sino también su resistencia a enfermedades y climas extremos, y a la escasez de agua y forraje. Los recursos genéticos bovinos con adaptación a climas cálidos son principalmente las razas de Bos indicus; sin embargo, algunas razas Bos taurus, como las denominadas criollas, pueden ser una alternativa viable en climas tropicales y su comportamiento productivo puede ser similar o superior que el de razas cebuínas (Hammond et al., 1996).
En general, las razas criollas de América Central, del Caribe y Sudamérica, incluyendo al Romosinuano, han mostrado adaptación a los retos ambientales de zonas tropicales y subtropicales del mundo (De Alba, 2011). Las razas criollas bovinas han evolucionado en condiciones adversas, lo que surgiere que son animales que poseen genes para adaptación en frecuencias distintas a las de razas exóticas, por lo que es conveniente caracterizar, conservar y utilizar de manera sostenible estos recursos (Núñez-Domínguez et al., 2016).
La raza Romosinuano es una raza criolla desarrollada en Colombia, que actualmente se encuentra en otros países de América, principalmente en Costa Rica, Estados Unidos, Venezuela y México (De Alba, 2011). Una característica sobresaliente en este ganado es su resistencia al estrés por calor y a la alta humedad, así como a contingencias que se presentan en las zonas tropicales de América, como la presencia de parásitos externos e internos, plagas y enfermedades (Scharf et al., 2010; Carroll et al., 2011, 2012).
Domínguez-Viveros et al. (2009) mencionan que es importante identificar animales superiores genéticamente, para lograr una mejor eficiencia productiva y de características deseables en el producto final. En México, existen ganaderos organizados dedicados a la cría de Romosinuano desde 1998 (De Alba, 2011), actualmente con hatos ubicados en Campeche, Michoacán, Tabasco, Tamaulipas y Veracruz; sin embargo, no se encontraron estudios que sirvan como base para establecer programas de mejoramiento genético. El conocimiento de parámetros genéticos es necesario para evaluar las posibilidades de mejoramiento genético de la raza Romosinuano en México. El objetivo de este estudio fue estimar parámetros y tendencias genéticas de variables de crecimiento en la población de ganado Romosinuano que se encuentra en México.
Método
Origen y características de los datos
La información genealógica y productiva utilizada en este estudio provino de la “Asociación Mexicana de Criadores de Ganado Romosinuano y Criollo Lechero Tropical A. C. (AMCROLET)”, donde se tomaron los datos de la identificación del animal y de sus padres, sexo, criador, fecha de nacimiento, y los datos productivos de peso al nacimiento y al destete. Los registros de comportamiento provienen de ranchos ubicados en áreas tropicales de México (estados de Veracruz y Tabasco), en donde los animales son alimentados únicamente con base en pastoreo de praderas (principalmente Brachiaria brizantha y Cynodon plectostachyus). Los becerros son destetados aproximadamente a los ocho meses de edad.
Las características estudiadas fueron los pesos al nacimiento (PN) y al destete ajustado a 240 días (PD). Los registros de pesos usados en los análisis fueron obtenidos de animales nacidos entre 2002 y 2016, provenientes de cuatro unidades de producción. Se depuró la base de datos, eliminando animales con errores en sus fechas de nacimiento, con padres desconocidos, muy alejados de la media general de cada característica (±3σ), que no pertenecieran a un grupo contemporáneo (GC = año, hato, época, sexo) y pertenecientes a GC que no presentaran conectividad genética entre ellos; esto último fue determinado usando el programa AMC (Roso y Schenkel, 2006). La información final utilizada incluyó 895 registros de PN y 517 de PD. Las edades de las madres al parto variaron entre 1.7 y 17.7 años. El archivo de pedigrí tuvo información desde 1950 hasta 2016, con un total de 4,537 animales. Una descripción general con los estadísticos obtenidos de los registros se muestra en el Cuadro 1.
Análisis estadístico
Las estimaciones de componentes de varianza fueron obtenidas por máxima verosimilitud restringida (REML), usando el programa ASReml (Gilmour et al., 2015). Con el propósito de establecer el modelo de mejor ajuste para análisis univariados de PN y PD, se realizaron pruebas de significancia para identificar efectos fijos y aleatorios importantes.
Para seleccionar los efectos fijos en las características evaluadas, los modelos iniciales incluyeron el grupo contemporáneo, la covariable edad de la madre al parto (efectos lineal y cuadrático), y la covariable lineal de la proporción de genes Romosinuano (0.5 a 1). La significancia de los efectos (P < 0.05) fue realizada mediante la prueba del Estadístico de Wald del programa ASReml (Gilmour et al., 2015).
Para seleccionar los efectos aleatorios a incluir en el modelo final, se utilizaron modelos iniciales univariados donde se incluyeron los efectos: genético aditivo directo, genético aditivo materno (ignorando y considerando la correlación entre efectos directos y maternos), ambiente permanente, grupo contemporáneo (en lugar de éste como efecto fijo), interacción de semental por grupo contemporáneo, e interacción de semental por hato. Para considerar significativo un efecto, se realizaron pruebas de proporción de verosimilitudes (−log L) que se basan en las diferencias entre los logaritmos de las funciones de verosimilitud estimadas en el programa y comparadas con el valor de Chi-cuadrada con p grados de libertad, donde p es igual a la diferencia en el número de parámetros entre los modelos (Domínguez-Viveros et al., 2009).
Con base en los efectos significativos de análisis preliminares, se realizó un análisis final con el modelo animal bivariado para PN y PD, con el fin de estimar los componentes de varianza y parámetros genéticos. La notación matricial para el modelo fue:
Donde y es vector de observaciones, b es vector de efectos fijos, u es vector de efectos aleatorios genéticos aditivos directos, m es vector de efectos aleatorios genéticos aditivos maternos (sólo en PD), s es vector de efectos aleatorios para la interacción semental por grupo contemporáneo, y e es vector de efectos residuales; X , Z , N , y W , son matrices de incidencia para su vector correspondiente; y los subíndices 1 y 2 corresponden a las características de PN y PD, respectivamente. Las suposiciones del modelo fueron:
Donde 𝑉 = 𝑍𝐺𝑍′ + 𝑁𝑀𝑁′ + 𝑊𝑆𝑊′ + 𝑅, 𝐺 = 𝐴 ⨂ 𝐺0, 𝑀 = 𝐴 ⨂ 𝑀0, 𝑆 = 𝐼𝑠 ⨂ 𝑆0, 𝑅 = 𝐼𝑒 ⨂ 𝑅0, 𝐴 es la matriz de relaciones genéticas aditivas entre los animales, G0es la matriz de (co)varianzas para los efectos genéticos aditivos directos entre PN y PD, M0 es la matriz de (co)varianzas para los efectos genéticos aditivos maternos, S0 es la matriz de (co)varianzas para la interacción semental por grupo contemporáneo entre PN y PD, R0 es la matriz de (co)varianzas para los efectos residuales entre ambas características, Is es la matriz identidad de orden igual al número de interacciones semental por grupo contemporáneo, Ie es la matriz identidad de orden igual al número de registros, y ⨂ es el producto Kronecker.
Las tendencias genéticas para las estimaciones de PN y PD directo, y de PD materno fueron calculadas mediante regresión lineal del promedio de los valores genéticos sobre el año de nacimiento de los bovinos, utilizando el procedimiento REG del paquete estadístico SAS (SAS, 2013). Los animales con registro en el pedigrí fueron nacidos desde el año 1950; sin embargo, dado que antes de 1998 (año del establecimiento legal de AMCROLET) se tuvieron pocos o ningún registro en años específicos, las tendencias genéticas se reportan sólo para animales nacidos durante el periodo de 1998 a 2016.
Resultados y discusión
En los modelos univariados, para ambas características, los estimadores de varianza genética aditiva (
Cuadro 2 Valores estimados de (co)varianzas, y parámetros genéticos y ambientales para pesos al nacimiento (PN) y al destete (PD) de la raza Romosinuano en México.
| Estimadores | PN | PD |
|---|---|---|
| Varianza genética aditiva directa, kg2 | 3.27 (1.25)* | 121.97 (76.23) |
| Varianza genética aditiva materna, kg2 | --- | 22.34 (30.61) |
| Varianza de la interacción SxGC** ( |
3.70 (0.87) | 68.25 46.74) |
| Varianza residual, kg2 | 10.39 (1.21) | 379.08 (57.35) |
| Varianza fenotípica ( |
17.37 (1.00) | 591.67 (51.10) |
| Heredabilidad directa | 0.19 (0.08) | 0.21 (0.12) |
| Heredabilidad materna | --- | 0.04 (0.05) |
|
|
0.21 (0.05) | 0.11 (0.08) |
* Valores dentro de paréntesis representan los errores estándar respectivos.
** SxGC = semental por grupo contemporáneo.
Heredabilidad directa
Para PN se estimó un valor de heredabilidad directa
En el presente estudio, la estimación de
Algunos valores estimados para este parámetro difieren más por la diferencia en los modelos utilizados, que por la diferencia en variabilidad genética que se encuentra en cada población, ya que la
Heredabilidad materna
La heredabilidad materna
La estimación de
Interacción semental por grupo contemporáneo
La inclusión de la interacción semental x grupo contemporáneo fue significativa tanto para PN como para PD, lo que coincide con diversos estudios (Gutiérrez et al., 2006; Gallegos-Ramírez et al., 2011; Neser et al., 2012) documentando la importancia de la inclusión de este efecto en la predicción de valores genéticos. Para el caso de PN, la proporción de la varianza fenotípica explicada por la interacción semental x grupo contemporáneo (s2) fue un valor muy alto (0.21). Esta estimación fue mayor que las publicadas para Asturiana de los Valles (Gutiérrez et al., 2006), Brangus (Neser et al., 2012) y Limousin (Van Niecker y Neser, 2006), con valores de 0.04, 0.07 y 0.13, respectivamente.
La estimación (Cuadro 2) de s2 para PD (0.11) fue menor que para PN. Sin embargo, este valor fue mayor que los publicados para Asturiana de los Valles (Gutiérrez et al., 2006), Angus (Berweger et al., 1999) y Brangus (Neser et al., 2012), con estimaciones de 0.07, 0.07 y 0.10, respectivamente. Aunque es conocido que el mecanismo de acción en esta varianza es impreciso y más de un factor puede intervenir en esta característica, algunas razones pueden ser el trato preferencial de sementales con hembras seleccionadas (Notter et al., 1992) o diferencias dentro de grupos contemporáneos no contabilizadas (Berweger et al., 1999). Una de las razones posibles por las cuales s2 explicó gran parte de la varianza fenotípica en este estudio, está dada por las restricciones sanitarias para importar animales de esta raza de su país de origen, ya que esta población se maneja de manera prácticamente cerrada, por lo cual los empadres no son de manera aleatoria, dando así un trato preferencial a los sementales en algunos casos.
Correlación genética y ambiental entre características de crecimiento
La estimación de coeficiente de correlación genética entre PN y PD (rg) fue 0.69 ± 0.28, la que puede ser considerada como una correlación alta. Lo anterior implica que si se realiza selección para incrementar el peso al nacer provocará también un aumento en el peso al destete y viceversa. Esta estimación fue mayor que las reportadas por Van Niecker y Neser (2006) en Limousin (0.41), y Meyer (1993) en Hereford (0.56). Valores superiores fueron reportados por Ossa et al. (2008) en Costeño con Cuernos (0.74), Meyer (1994) en Angus (0.76) y Neser et al. (2012) en Brangus (0.78). En el presente estudio, la estimación de la correlación de residuales (re) fue baja (0.07) y menor que los valores reportados por (Ossa et al., 2008) para la raza Costeño con Cuernos (0.13) y por Robinson (1996) en Angus (0.14), lo que puede sugerir que el efecto ambiental es un tanto independiente entre PN y PD. La correlación fenotípica (rp) también presentó un valor bajo y positivo (0.15). Dado el valor de
Tendencias genéticas
La tendencia de los valores genéticos anuales promedio para PN fue variable a través de los años (Figura 1), negativa (-11 g/año), pero con pendiente no diferente de cero (P > 0.05). Este resultado es favorable, pues generalmente no es conveniente que existan modificaciones importantes en el peso al nacimiento promedio de las crías para la población, excepto cuando algunas crías en particular nacen muy débiles o se tienen problemas de partos distócicos.
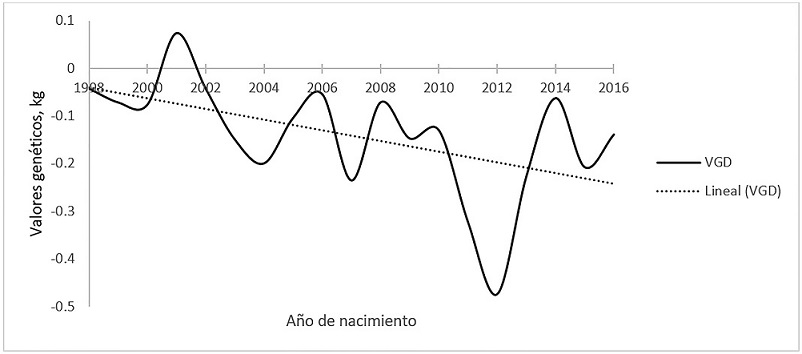
Figura 1 Tendencia de los valores genéticos directos (VGD) para peso al nacimiento en bovinos Romosinuano.
Estos resultados son similares a los valores publicados para otras razas criollas, Costeño con Cuernos (Ossa et al., 2008) y Sanmartinero (Martínez et al., 2009b), debido a que son poblaciones que tampoco se han sometido a programas de selección. Comparando los resultados con otras razas de bovinos para carne en México, en el caso tanto de Brangus como de Salers (Domínguez-Viveros et al., 2009) se obtuvo una ligera tendencia negativa, con -9 (P > 0.01) y -22 g/año (P > 0.05), respectivamente, lo que los autores explican como resultado de la búsqueda de los criadores por evitar partos distócicos en ambas poblaciones.
Tomando en cuenta la tendencia de los valores genéticos directos y maternos para PD (Figura 2), se puede observar, que en forma similar a lo ocurrido con PN, no presentaron tendencias constantes, además que las pendientes tanto de los valores genéticos directos (P > 0.5) como los maternos (P > 0.1), aunque negativas (-9 y -18 g/año) no fueron diferentes de cero. En forma similar que para PN, la razón de que no se encuentre una tendencia en aumento que muestre un mejoramiento en estas características, puede ser debido a que no se ha realizado ningún programa de mejoramiento en la población Romosinuano de México, ya que recientemente se produjo la primera evaluación genética de estos animales (AMCROLET, 2017).
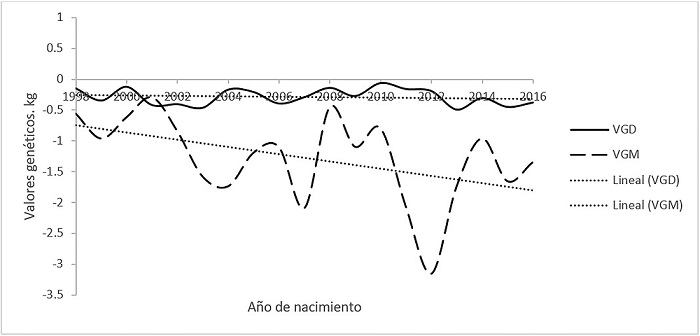
Figura 2 Tendencia de los valores genéticos directos (VGD) y maternos (VGM) para peso al destete en bovinos Romosinuano.
En otras razas que se encuentran en México y en las que se han realizado programas de selección, como en Brangus y Salers, se muestran tendencias genéticas para PD de 329 (P < 0.01) y 46 (P > 0.10) g/año, respectivamente (Domínguez-Viveros et al., 2009). Así mismo, Parra-Bracamonte et al. (2007) reportaron una tendencia positiva (P < 0.05) en ganado Brahman, con incrementos del 0.05% de la media de esa población. Martínez y Pérez (2006) reportaron para la raza Romosinuano en Colombia una tendencia de aumento (161 g/año), aunque no fue diferente de cero (P > 0.05). Lo anterior sugiere la necesidad de un programa de selección en la población de Romosinuano en México, para el mejoramiento en estas características.
Conclusiones
Aunque las estimaciones de las heredabilidades para peso al nacimiento y peso al destete son relativamente bajas en la población de bovinos Romosinuano en México, muestran suficiente variabilidad genética para mejorar el crecimiento predestete por medio de selección con base en valores genéticos predichos. Se recomienda continuar con el registro y análisis de información en la raza, para realizar evaluaciones genéticas con una mayor cantidad de datos productivos y genealógicos.

