











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La irradiación gamma en semillas puede ser utilizada como agente mutagénico para inducir variabilidad genética en las plantas e inducir variabilidad fenotípica, que permita seleccionar plantas con características deseables. No obstante, antes de comenzar un programa de mejoramiento genético mediante mutagénesis, es necesario establecer las dosis de irradiación con mayor probabilidad de producir mutaciones efectivas. Diversos investigadores concuerdan en que existe mayor probabilidad de producir mutaciones efectivas para hacer mejoramiento genético con dosis donde mueren el 50% de los individuos irradiados (Morela et al., 2002; Golubinova y Gecheff, 2011; Ángeles-Espino et al., 2013); esto es conocido como dosis letal media (DL50). Por otro lado, investigadores como Akgüm y Tosun (2004) y Khalil et al. (2014) , señalan que en dosis donde existe alta probabilidad de encontrar mutaciones efectivas, es cuando se reduce el crecimiento 50% (RC50). Por otra parte, el genoma sufre múltiples impactos con dosis altas que producen aberraciones o cambios negativos comúnmente (Songsri et al., 2011; Thole et al., 2011).
Los pastos navajita (Bouteloua gracilis), banderita (Bouteloua curtipendula), buffel (Pennisetum ciliare) y llorón (Eragrostis curvula) son algunas especies con potencial para ser utilizadas en programas de revegetación para zonas áridas y semiáridas (Moreno-Gómez et al., 2012; Sáenz-Flores et al., 2015). No obstante, estas especies presentan problemas que pueden mejorarse mediante herramientas biotecnológicas. Las especies nativas de alto valor forrajero como navajita y banderita han presentado problemas para establecerse en programas de rehabilitación de pastizales (Melgoza-Castillo et al., 2007). Ante esto, se ha optado por utilizar especies introducidas con mayor potencial de establecimiento como buffel y llorón. Sin embargo, comparadas con algunas nativas, estas presentan menor valor forrajero y el ganado las prefiere menos (Quero et al., 2007). En este sentido, las especies mencionadas pueden ser sometidas a programas de mejoramiento. Éste puede llevarse a cabo a través de mutagénesis mediante irradiación gamma, ya que esta técnica ha demostrado ser eficiente para inducir mutaciones, aun en genomas complejos como los de las gramíneas (Ousmane et al., 2013; Ambli y Mullainathan, 2014). No obstante, a la fecha se desconoce la dosis de irradiación gamma con mayor probabilidad de producir mutaciones efectivas en estas especies. Por lo anterior, el objetivo fue determinar la DL50 y RC50 para conocer la dosis óptima de irradiación en semilla de pasto llorón, buffel, banderita y navajita. Este estudio facilitará la inducción de nuevo material genético de interés agronómico y ecológico, que pueda utilizarse en programas de revegetación en áreas degradadas.
Materiales y Métodos
El experimento se llevó a cabo en condiciones de laboratorio. El germoplasma utilizado fue semilla irradiada con Co60 de los pastos navajita, banderita, buffel y llorón. Se plantearon seis dosis de irradiación con Co60: 100, 200, 300, 450, 600, 900 Gray (Gy) y un tratamiento control (T-0, sin irradiar). Los tiempos de exposición para las dosis evaluadas fueron determinados utilizando el sistema Gafchromic dosimetry (Li et al., 1995) y una cámara de ionización marca RADCAL modelo Acudose No. de serie 4094118, USA. La irradiación de la semilla se realizó en el Complejo MOSCAFRUT SAGARPA/IICA en Metapa de Domínguez, Chiapas, con la colaboración del Campo Experimental Rosario Izapa, INIFAP, Chiapas, México. Las dosis se aplicaron en un irradiador panorámico tipo Gamma Beam Modelo GB-127 MDS Nordion, con fuente de almacenamiento en seco y actividad de 15,000 Curies de 60Co.
Para evaluar el efecto por irradiación, se realizaron pruebas de germinación. Además, se analizó la viabilidad de las semillas del tratamiento control mediante tetrazolio (C19H15ClN4), para verificar si se presentó efecto de radiestimulación. Para las pruebas de germinación se irradió semilla comercial adquirida en Seed Curtis & Curtis, INC. Las especies utilizadas fueron navajita variedad (var.) Hachita; banderita var. Vaughn; llorón var. Ermelo; y buffel var. T-4464. De la semilla irradiada se tomaron cuatro muestras al azar por especie y se separaron glumas, lemma y palea, para utilizar solamente la cariópside desnuda. La prueba de germinación se llevó a cabo en cajas Petri de 90 mm de diámetro, provistas de algodón y papel filtro y se incluyeron cuatro repeticiones por tratamiento. La unidad experimental fue la caja Petri con 50 semillas. Cada repetición fue humectada con 25 ml de agua destilada al inicio de la prueba y posteriormente se suministraron riegos por aspersión de 2.0 ml cada dos días, durante los 20 d de la experimentación. Las cajas Petri se colocaron en incubadora Precision Scientific, modelo 6M a temperatura de 26 ± 2 °C. Se consideró semilla germinada aquella que alcanzó 0.5 cm de plúmula o de radícula. Las variables evaluadas fueron: porcentaje de germinación (%GE); índice de velocidad de germinación (IVG); longitud de radícula (LR); y longitud plúmula (LP). Para obtener LR y LP se dejaron crecer tres plántulas por caja Petri durante siete días después de germinadas. El IVG se calculó mediante la ecuación propuesta por Maguire (1962):
Donde: IVG = índice de velocidad de germinación; n¡=número de semillas germinadas por día; t = día de la germinación.
Las semillas del tratamiento control fueron evaluadas mediante la prueba de viabilidad con tetrazolio al 1%. Para esto, se utilizaron cuatro repeticiones de 25 cariópsides seleccionadas al azar, las cuales fueron colocadas en la solución durante 8 h y se visualizaron en un estereoscopio Zeiss, modelo Stemi DV4. Las semillas fueron consideradas viables cuando presentaron una tinción purpura. Al aplicar la prueba del tretrazolio, la semilla utilizada presentó un 92% de viabilidad para el pasto llorón, 68% en buffel, 92% en banderita y 88% en navajita. El porcentaje de viabilidad en llorón, buffel y banderita concuerda con su porcentaje de germinación. Sin embargo, el pasto navajita presentó 20% menos de germinación al compararla con la viabilidad obtenida. Por otro lado, la semilla de todas las especies evaluadas presentó bajo contenido de humedad. El pasto llorón presentó 6.74%, el buffel 3.59%, el banderita 4.63% y el navajita 3.92% de humedad. Además, se determinó el porcentaje de humedad en la semilla a través de secado en una estufa Precision Scientific, modelo 6M a temperatura constante de 90 °C por 48 h. La semilla seca se pesó en una balanza analítica de la marca Viper BC, Mettler Toledo y el porcentaje de humedad se calculó por diferencia de pesos.
Los datos fueron analizados mediante regresión lineal para cada variable y especie por separado. El modelo que mejor se ajustó a los datos fue determinado mediante el software CurveExpert 2.6. Los criterios de evaluación utilizados fueron el coeficiente de determinación (R2) y el criterio de información de Akaike corregido (AICc). La significancia de los modelos se obtuvo mediante el PROC REG de SAS (2006). Además, se realizó una comparación de medias con la prueba de Dunnett (1955), a un nivel de significancia de 0.05 (α=0.05). Con la ecuación de regresión estimada fue obtenida la DL50, con base en %GE y RC50, de acuerdo con IVG, LR y LP. Debido a que los valores de DL50 y RC50 presentaron un amplio rango de variación, se optó por calcular una media ponderada con los resultados de DL50 y RC50 obtenidos de las 4 variables evaluadas. La DL50 se ponderó con 55%, mientras que los valores de RG50 se ponderaron con 15% cada uno. Cabe señalar que la mayor ponderación se le dio a la germinación, ya que la muerte del individuo es el máximo daño que puede producir la irradiación.
Resultados
En pasto llorón, el comportamiento del porcentaje de germinación y el índice de velocidad de germinación se ajustó a un modelo de rendimiento-densidad conocido como Farazdaghi-Harris (P<0.0001). Esto debido a que con incremento de dosis de irradiación de 300 a 450 Gy, la germinación decayó de 98 a 36% (Figura 1). Por otro lado, el comportamiento lineal fue significativo (P<0.0001) para longitud de plúmula y longitud de radícula (Cuadro 1). En esta especie, los coeficientes de determinación (R2) fluctuaron desde 0.79 hasta 0.9. Además, las dosis de 600 y 900 Gy fueron las únicas que presentaron diferencias (P<0.05) con respecto al testigo en todas las variables evaluadas (Cuadro 2). La DL50 se encontró a 459 Gy, mientras que la RC50 se obtuvo a 396 Gy para el IVG, 960 Gy para LP y 793 Gy para LR. Por lo anterior, la media ponderada de las dosis medias se obtuvo a los 574 Gy.
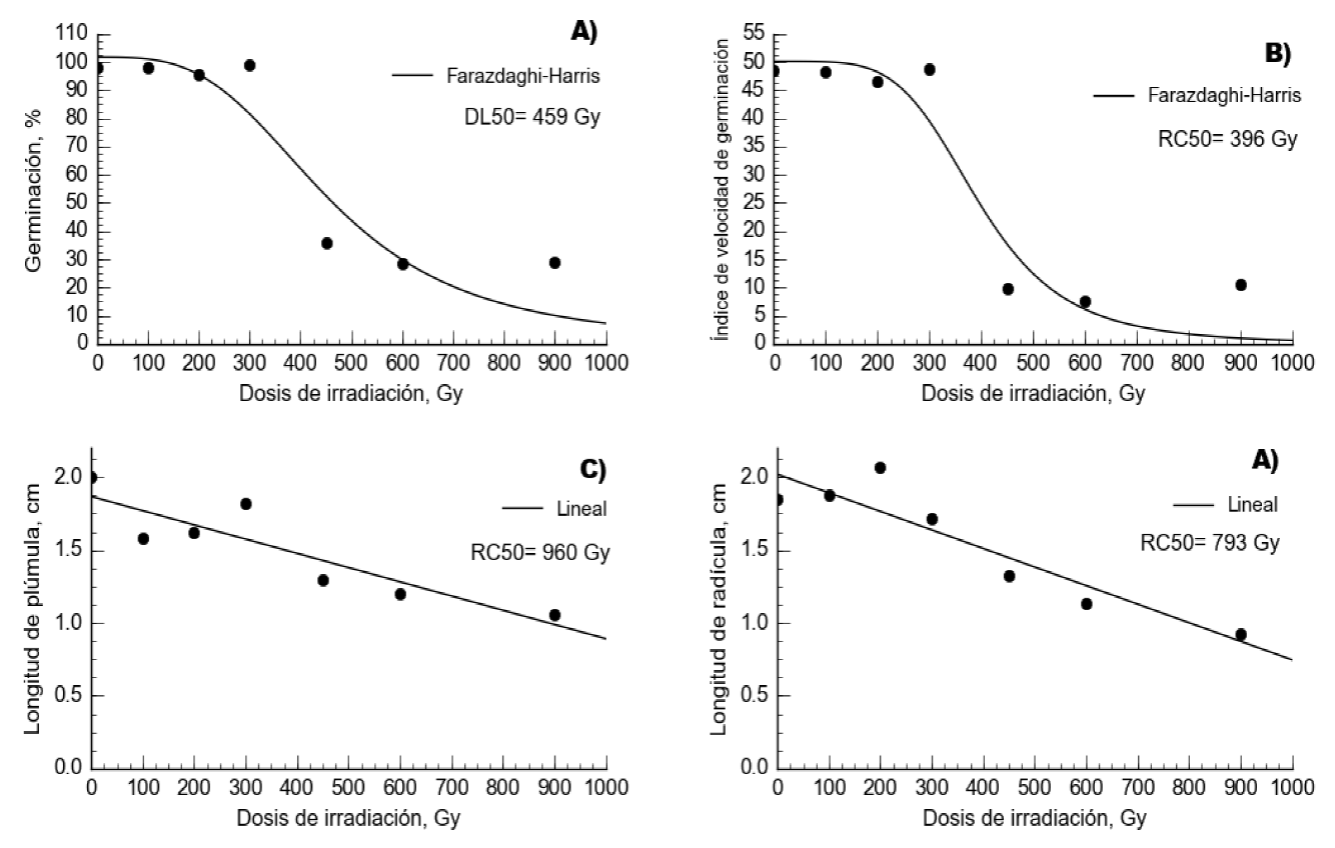
Figura 1 Irradiación con Co60 en semilla de pasto llorón (Eragrostis curvula) y su efecto en comportamiento del porcentaje de germinación (a), índice de velocidad de germinación (b), longitud de plúmula (c) y longitud de radícula (d). DL50= Dosis letal media y RC50= reducción media del crecimiento, de acuerdo a la línea de tendencia.
Cuadro 1 Modelos de regresión a los que se ajustaron el porcentaje de germinación, índice de velocidad de germinación, longitud de plúmula y longitud de radícula de cuatro especies de pastos sometidas a diferentes dosis de irradiación gamma.
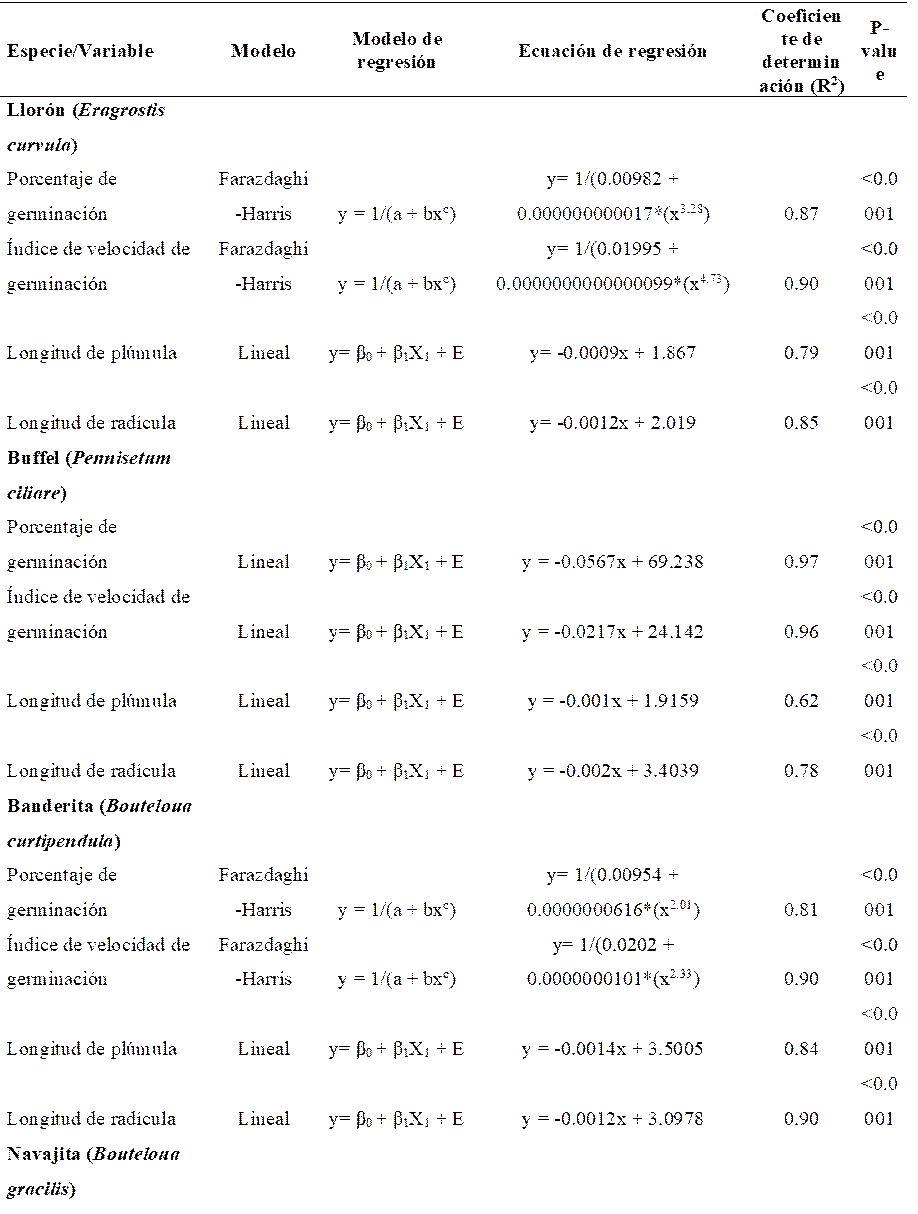
Cuadro 2 Parametros de germinación de cuatro especies de pastos sometidas a diferentes dosis de irradiación gamma con Co60.
| Especie/dosis (Gy) | Variables | ||||
|---|---|---|---|---|---|
| Porcentaje de germinación (%) | Índice de velocidad de germinación | Longitud de plúmula (cm) | Longitud de radícula (cm) | ||
| Llorón (Eragrostis curvula) | |||||
| Testigo | 98.0 | 48.6 | 2.0 | 2.1 | |
| 100 | 98.0 | 48.2 | 1.8 | 1.9 | |
| 200 | 95.5 | 46.5 | 1.6 | 1.9 | |
| 300 | 99.0 | 48.9 | 1.6 | 1.7 | |
| 450 | 36.0* | 9.9* | 1.3* | 1.3 | |
| 600 | 28.5* | 7.5* | 1.2* | 1.1* | |
| 900 | 29.0* | 10.5* | 1.1* | 0.9* | |
| Error estándar (±) | 1.89 | 0.48 | 0.14 | 0.12 | |
| Buffel (Pennisetum ciliare) | |||||
| Testigo | 67.5 | 24.8 | 1.8 | 3.1 | |
| 100 | 65.0 | 21.1 | 2.2 | 3.1 | |
| 200 | 56.0 | 18.4 | 1.3 | 2.9 | |
| 300 | 56.0 | 19.6* | 1.8 | 3.3 | |
| 450 | 40.0* | 13.5* | 1.4 | 2.7 | |
| 600 | 39.0* | 12.4* | 1.2* | 2.6 | |
| 900 | 16.5* | 4.0* | 1.1* | 1.2* | |
| Error estándar (±) | 3.16 | 1.32 | 0.12 | 0.25 | |
| Banderita (Bouteloua curtipendula) | |||||
| Testigo | 94.5 | 33.6 | 3.7 | 3.2 | |
| 100 | 94.5 | 32.6 | 3.3 | 3.1 | |
| 200 | 94.0 | 33.7 | 3.3 | 2.9 | |
| 300 | 93.0 | 31.3 | 3.1 | 2.5 | |
| 450 | 18.5* | 5.4* | 2.9 | 2.4 | |
| 600 | 16.0* | 3.9* | 2.3* | 2.4 | |
| 900 | 16.5* | 3.8* | 2.5* | 2.1* | |
| Error estándar (±) | 1.91 | 1.24 | 0.24 | 0.21 | |
| Navajita (Bouteloua gracilis) | |||||
| Testigo | 67.5 | 25.2 | 3.0 | 2.5 | |
| 100 | 90.5* | 37.4* | 3.1 | 2.3 | |
| 200 | 83.5* | 34.4* | 3.9* | 2.5 | |
| 300 | 89.5* | 35.8* | 2.9 | 2.0 | |
| 450 | 6.0* | 0.3* | 0.7* | 0.5* | |
| 600 | 1.0* | 0.1* | 0* | 0* | |
| 900 | 0* | 0* | 0* | 0* | |
| Error estándar (±) | 0.95 | 0.40 | 0.16 | 0.11 |
*Diferencias significativas (Dunnett, P<0.05) respecto al tratamiento testigo (0 Gy)
El comportamiento de todas las variables evaluadas en pasto buffel se ajustó a una tendencia lineal (P<0.0001; Figura 2). Para estas variables, los coeficientes de determinación fluctuaron desde 0.62 hasta 0.97 (Cuadro 1). En esta especie, la única dosis que presentó diferencias en todas las variables respecto a T-0 (P<0.05) fue la de 900 Gy (Cuadro 2). No obstante, la DL50 se encontró a 610 Gy y la RC50 a 556 Gy para el IVG, 917 Gy para LP y 870 Gy para LR. De acuerdo con lo anterior, la media ponderada de las dosis medias se calculó a los 686 Gy.
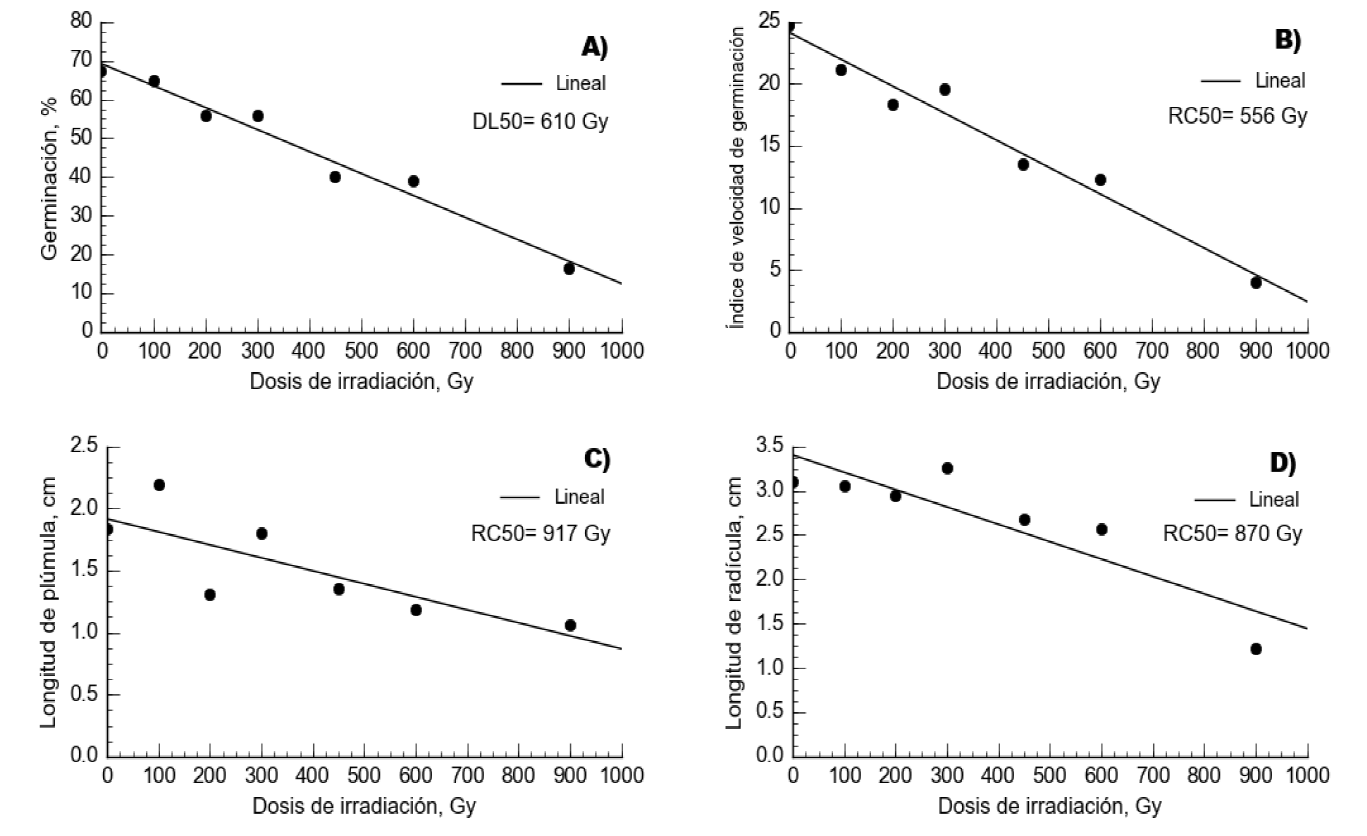
Figura 2 Irradiación con Co60 en semilla de pasto buffel (Pennisetum ciliare) y su efecto sobre el porcentaje de germinación (a), índice de velocidad de germinación (b), longitud de plúmula (c) y longitud de radícula (d). DL50= Dosis letal media y RC50= reducción media del crecimiento, de acuerdo a la línea de tendencia.
Al igual que en pasto llorón, el comportamiento del porcentaje de germinación y el índice de velocidad de germinación se ajustaron al modelo de Farazdaghi-Harris (P<0.0001), en pasto banderita (Cuadro 1). Así mismo, la germinación disminuyó de 93 a 18.5% cuando la dosis de irradiación se incrementó de 300 a 450 Gy (Figura 3). En esta especie la única dosis que mostró diferencias respecto a T-0 (P<0.05) en todas las variables fue la de 900 Gy (Cuadro 2). En pasto banderita la DL50 se presentó con 355 Gy. No obstante, para el IVG la RC50 se encontró en 491 Gy, para LP en 1271Gy y para LR en 1250 Gy. Por otro lado, el valor de la media ponderada fue de 647 Gy.

Figura 3 Irradiación con Co60 en semilla de pasto banderita (Bouteloua curtipendula) y su efecto sobre el porcentaje de germinación (a), índice de velocidad de germinación (b), longitud de plúmula (c) y longitud de radícula (d). DL50= Dosis letal media y RC50= reducción media del crecimiento, de acuerdo a la línea de tendencia.
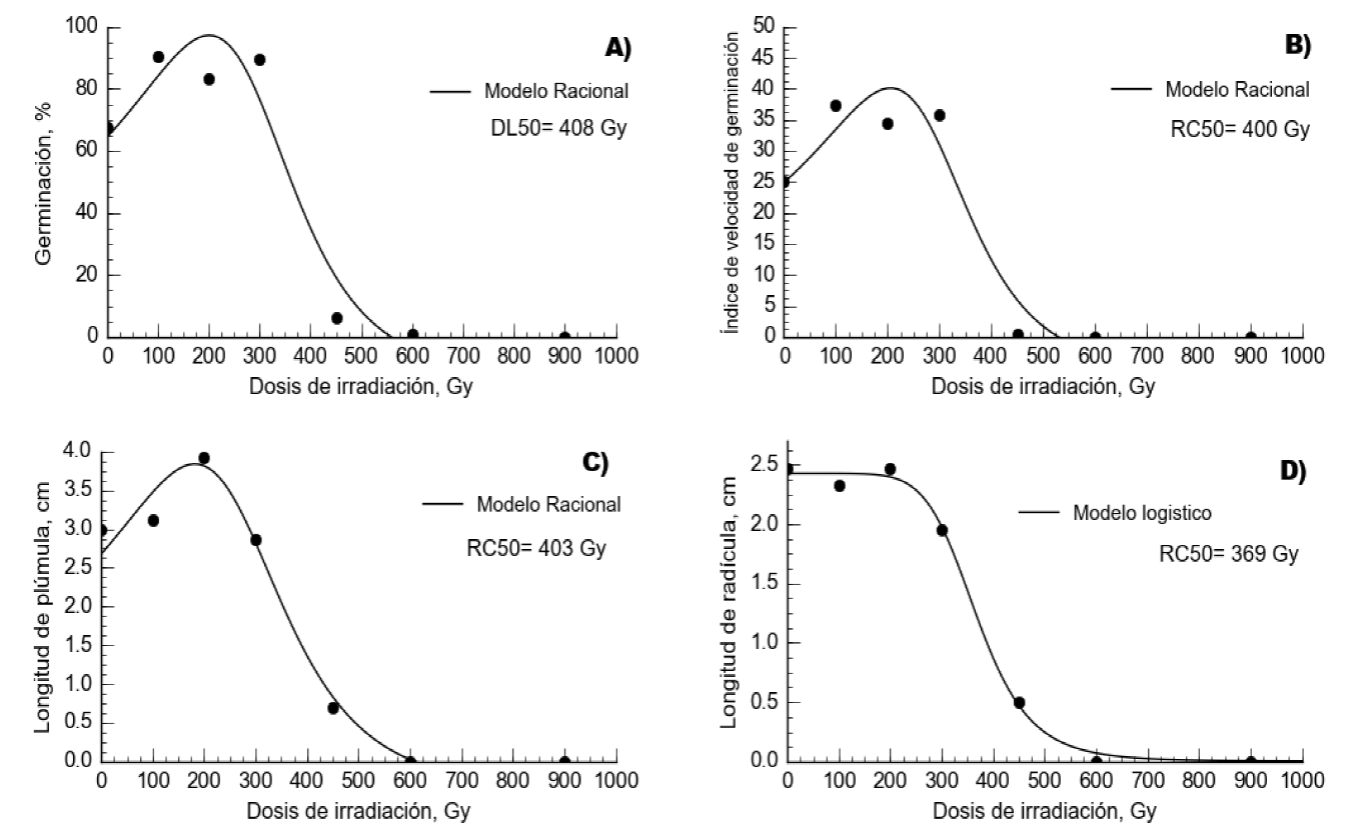
En pasto navajita el comportamiento del %GE, IVG y LP se ajustó a un modelo de regresión racional (P<0.0001), mientras que LR se ajustó a un modelo sigmoideal logístico (P<0.0001) (Cuadro 1). En esta especie, el %GE y el IVG presentaron un incremento (P<0.05) con las dosis de 100, 200 y 300 Gy, con relación al testigo (Figura 4). De igual manera, se presentó un incremento (P<0.05) en LP con las dosis de 300 Gy (Cuadro 2). Sin embargo, en todas las variables los valores obtenidos con las dosis de 450, 600 y 900 Gy fueron menor (P<0.05) a los del tratamiento testigo. De acuerdo con la ecuación de predicción, la DL50 se presentó a los 408 Gy para pasto navajita. De manera similar, la RC50 en el IVG se presentó a los 400 Gy, en LP a 403 Gy y en LR a 369 Gy. La media ponderada de estos valores fue de 400 Gy.
Discusión
La germinación del pasto llorón, buffel y banderita disminuyó conforme aumentó la dosis de irradiación. Esto concuerda con diversos autores quienes mencionan que la germinación de diferentes especies de plantas tiende a disminuir a medida que aumenta la dosis de irradiación. Anbarasan et al. (2013) encontraron una reducción 94% a 27% en la germinación de semilla de ajonjolí (Sesamum indicum) al incrementar la dosis de irradiación de 0 a 1000 Gy. De manera similar, Bharathi et al., (2013) observaron que la germinación de ashwagandh (Withania somnifera) disminuyó 75% al ser irradiada a 500 Gy. Así mismo, en otra investigación en arroz (Oryza sativa) encontraron que la germinación disminuyó un 66% con 350 Gy como dosis de irradiación (Rajarajan et al., 2016). Lo anterior, se debe a que dosis altas de irradiación inhiben las funciones vitales de las células; lo cual puede provocar la muerte del embrión en la semilla. Este fenómeno aumenta conforme se incrementa la dosis de irradiación y es la causa por la que la germinación disminuye (Olasupo et al., 2016). En contraste, el pasto navajita mostró un incremento en germinación de alrededor del 20% con dosis de 100, 200 y 300 Gy. Este resultado concuerda con el porcentaje de viabilidad obtenido para esta especie. Este fenómeno es silmilar con lo señalado por González y Nakayama (2015), quienes encontraron que bajas dosis de irradiación gamma pueden estimular hasta en un 23% la germinación de Stevia rebaudian. Lo anterior, probablemente se debe a que la exposición de la semilla a irradiación gamma, frecuentemente causa la destrucción de algunos compuestos relacionados con la germinación como el ácido ascórbico. Una de las funciones de esta fitohormona es detener la división celular para inducir el estado de latencia en las semillas (Shah et al., 2008).
La DL50 encontrada en pasto buffel fue similar a lo reportado por Ousmane et al. (2013). Estos investigadores encontraron en Pennisetum glaucum la DL50 en germinación a los 669.3 Gy mediante irradiación gamma. No obstante, en otro estudio en Pennisetum typhoidesAmbli y Mullainathan (2014) encontraron la DL50 a los 200 Gy. En otros estudios realizados en gramíneas, Harding et al. (2012) al exponer 13 variedades de arroz a irradiación gamma, encontraron que la DL50 varió de 345 y 423 Gy de acuerdo a la variedad. De manera similar, Ramchander et al. (2015) encontraron la DL50 en dos variedades de arroz en 288 y 354 Gy; mientras que, en esta misma especie, Rajarajan et al. (2016) encontraron la DL50 a 229 Gy. Por otro lado, en un estudio realizado en pasto sudan (Sorghum sudanense), Golubinova y Gecheff (2011) encontraron que la DL50 varió entre 307 y 342 Gy de acuerdo a la variedad.
En general la DL50 de las especies analizadas se presentó en altas dosis de irradiación. Lo anterior probablemente se debe a que es semilla ortodoxa con bajo contenido de humedad. En este sentido, la radiación ionizante puede provocar la ruptura de enlaces covalentes y descomposición de moléculas de agua. Esto hace que se formen radicales libres que pueden dañar los diferentes organelos de la célula. Adicionalmente, estos radicales pueden ocasionar daños en la molécula de ADN, lo que a su vez puede inducir inversiones, translocaciones y cambios en la morfología de los cromosomas. Por esta razón, cuando el material a irradiar posee bajo contenido de humedad para que haya un efecto por radiación, se deben considerar dosis altas (Alegre, 2001), lo cual concuerda con los resultados en este estudio.
El IVG del pasto llorón, buffel y banderita disminuyó conforme aumentó la dosis de irradiación. Resultados similares fueron encontrados por Aparna et al. (2013), quienes encontraron que la velocidad de germinación en cacahuate (Arachis hypogaea) disminuyó al incrementar la dosis de irradiación de 58.4 a 6.8 con dosis de 2300 Gy. Es probable que el retraso en la germinación de semillas irradiadas probablemente se deba a que dosis altas de irradiación, inhiben funciones vitales para la célula como la síntesis de proteínas y la actividad enzimática; esto a su vez retrasa la división celular y por ende la germinación (Chandrashekar, 2014; Olasupo et al., 2016). Por otro lado, en pasto navajita la velocidad de germinación aumentó con las dosis de 100, 200 y 300 Gy. Este resultado concuerda con lo reportado por Maity et al. (2005), quienes encontraron incremento en la velocidad de germinación de semillas de arroz, al someterse a dosis bajas de irradiación. De manera similar, de la Fe et al. (2000) encontraron aumento en la velocidad de germinación de semillas de soya (Glycine max) en dosis de entre 240 y 280 Gy. Con relación a esto, Chandrashekar (2014) encontró que dosis bajas de irradiación pueden estimular la actividad metabólica de las células, la división celular y la actividad enzimática. Lo anterior pudo haber sido la causa del aumento en la velocidad de germinación del pasto navajita con dosis bajas de irradiación. |
En general la longitud de plúmula y radícula de las especies analizadas disminuyó conforme aumentó la dosis de irradiación. De manera similar, Ambavane et al. (2014) encontraron este efecto en Eleusine coracana, donde encontraron la RC50 a los 500 Gy para ambas variables. Por otro lado, Talebi y Talebi (2012) encontraron este mismo efecto en arroz. Respecto a esto, la disminución en longitud de plúmula y radícula al igual que el retraso en la velocidad de germinación, puede deberse a un retraso en la actividad metabólica y la división celular por causa de la irradiación (Chandrashekar, 2014; Olasupo et al., 2016). No obstante, la LP del pasto navajita mostró incremento en la dosis de 200 Gy, lo cual podría indicar que esta dosis de irradiación estimula los procesos metabólicos y la actividad proteica en esta especie (Chandrashekar, 2014). Algo similar se presentó en un estudio de Minisi et al. (2013), quienes al someter semilla de la planta ornamental Moluccella laevi a dosis de irradiación gamma entre 0 y 175 Gy, encontraron aumento en la germinación con dosis de 25 a 75 Gy.
La DL50 y las RC50 variaron en gran medida en todas las especies evaluadas en este estudio. Ante esta situación, se optó por establecer una media ponderada.
Conclusiones
La DL50 y RC50 fue determinada a diferentes dosis de irradiación en pasto llorón, buffel, banderita y navajita. Para programas futuros de mejoramiento genético por mutagénesis con irradiación gamma, se recomienda como dosis optima de irradiación alrededor de 574 Gy para pasto llorón, 686 Gy para buffel, 647 Gy en banderita y 400 Gy en el navajita.

