











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El crecimiento poblacional y el desarrollo humano han alterado al ambiente natural (Crosby y Blair 2001). Se estima que más del 75% de la superficie terrestre sin hielo del planeta muestra evidencia de alteración como resultado de la presencia humana y sus actividades (Ellis y Raman-kutty 2008). De los varios tipos de perturbación a gran escala que tienen impacto en las comunidades bióticas, la urbanización es considerada la más grave, ya que provoca grandes pérdidas de biodiversidad y su homogeneización (Vitousek et al. 1997, Pauchard et al. 2006, Ondine et al. 2008).
La urbanización está relacionada con la alteración de un terreno mediante estructuras que permiten satisfacer las necesidades de vivienda del hombre, lo cual modifica el hábitat original (Berkowitz et al. 2003). En las zonas rurales de manera tradicional se integran aspectos habitacionales y agropecuarios a pequeña escala, donde los habitantes son pocos y aprovechan los recursos naturales, con los que tienen una relación estrecha (Sancho-Comíns y Reinoso-Moreno 2012), por lo que generalmente conservan fragmentos de vegetación natural y pueden mantener una heterogeneidad ambiental amplia. Para conocer el impacto que la urbanización tiene en las comunidades bióticas se han llevado a cabo estudios que comparan áreas urbanas con áreas naturales ( Crosby y Blair 2001) o analizan gradientes de urbanización, para la cual se utilizan diferentes grupos de animales, tanto vertebrados como invertebrados (Blair 1996, Blair y Launer 1997, Denys y Schmidt 1998, Limburg y Schmidt 1990, Germaine y Wakeling 2001). A diferencia de otros grupos animales, las aves pueden ser estudiadas fácilmente por observadores experimentados, lo que las convierte en un grupo útil para explorar los efectos de la urbanización, así como las respuestas a diferentes diseños urbanos (Chace y Walsh 2006).
La urbanización tiene un impacto en la diversidad de las comunidades de aves, así como en la distribución y abundancia de las especies (Robbins et al. 1989, Chace y Walsh 2006). En estudios de gradientes de urbanización se ha observado una mayor riqueza en sitios poco perturbados (Blair 1996, Blair et al. 2001), mientras que para hábitats urba nizados se reporta una menor diversidad con predominio de aves exóticas (White et al. 2005), que, en conjunto con las alteraciones ambientales propias de la urbanización, puede llevar a la extirpación de algunas especies nativas en estos hábitats (Virkkala et al. 1993, Blair 1996). Un área muy urbanizada puede conducir entonces a un cambio en la composición de especies, un incremento en la abundancia de aves exóticas o ligadas a medios urbanos, y una reducción notable en la riqueza de aves. Este patrón ha sido observado en una variedad de hábitats urbanizados: bosques, desiertos y praderas (Chace y Walsh 2006). También hay evidencia que sugiere que el desarrollo urbano a escalas moderadas tiene un efecto en el incremento de la riqueza de la avifauna en comparación con hábitats no perturbados, ya que la heterogeneidad del ambiente permite la cohabitación de especies (Blair 1996, Fraterrigo y Wiens 2005). Esto último puede aplicarse a algunas zonas rurales mexicanas, donde la heterogeneidad del ambiente generalmente incluye áreas con vegetación natural con algún grado de alteración, pequeñas propiedades con actividades agropecuarias diversas, y casas habitación separadas unas de otras (Sancho-Comíns y Reinoso-Moreno 2012), con menos de 50% de superficies impermeables (mayor a este porcentaje corresponde a una ciudad sensuMacGregor-Fors 2011) y poca infrastructura urbana (suministro de agua, electricidad, drenaje, entre otras).
En México, los estudios enfocados a conocer el efecto de la urbanización en las comunidades de aves han sido escasos. Hasta hace pocos años, se ha incursionado más en este tema y se han realizado estudios principalmente en parques urbanos, así como en áreas urbanas asociadas a áreas naturales, con el objetivo de entender cómo la urbanización afecta la diversidad, la composición y la dinámica temporal de las comunidades de aves (Ortega Álvarez y MacGregor-Fors 2009, MacGregor-Fors y Schondube 2012). En la zona semiárida del centro de México se han realizado estudios sobre aves urbanas, periurbanas y exurbanas en la ciudad de Querétaro y zonas aledañas (Pineda-López et al. 2010, Pineda-López 2011, Malagamba-Rubio et al. 2013, Pineda-López et al. 2013, Pineda-López datos no publicados), en ellos se observan los patrones mencionados de reducción de la diversidad de aves en zonas urbanas (excepto para aves acuáticas) y una importante diversidad en zonas periurbanas. En esta zona, y en general para Latinoamérica, existe una falta de conocimiento sobre la ecología de las aves de asentamientos urbanos medianos o pequeños (MacGregor-Fors et al. 2017).
Este estudio tuvo como objetivos: 1) analizar la riqueza, diversidad y composición de la avifauna en ambientes con diferente grado de alteración en un paisaje urbano y rural del centro de México, 2) analizar las posibles asociaciones de la composición de especies con atributos ambientales. Los cuatro ambientes considerados representan un gradiente de alteración antrópica que incluye una ciudad pequeña de menos de 4,000 habitantes y una zona rural en la zona semiárdida circundante a la Peña de Bernal, Querétaro. Esperamos encontrar una menor diversidad de aves en la zona urbana, a pesar de su tamaño reducido en comparación con otras urbes, y una diversidad de aves más alta en la zona rural debido a que presenta un mosaico de cultivos, huertos y manchones de vegetación seminatural (con alteraciones mínimas), lo que puede considerarse un ambiente de alta heterogeneidad.
Métodos
Área de estudio
El área de estudio comprende dos poblaciones colindantes: Bernal (Villa de San Sebastián Bernal), que presenta una estructura principalmente urbana y cuenta con una población cercana a los 4,000 habitantes (INEGI 2010), y San Antonio de la Cal, que presenta una estructura principalmente rural, de acuerdo con lo indicado por Sancho y Reinoso (2012). Ambas áreas están situadas al sur y norte, respectivamente, del monolito denominado Peña de Bernal (20°44’55.12” N, 99°56’39.87” O), en el límite entre los municipios de Ezequiel Montes y Tolimán, Querétaro (Figura 1). A excepción de la zona urbana, el área de estudio se encuentra dentro del área natural protegida (ANP) “Peña de Bernal”, que tiene categoría de paisaje protegido (Segob 2009).
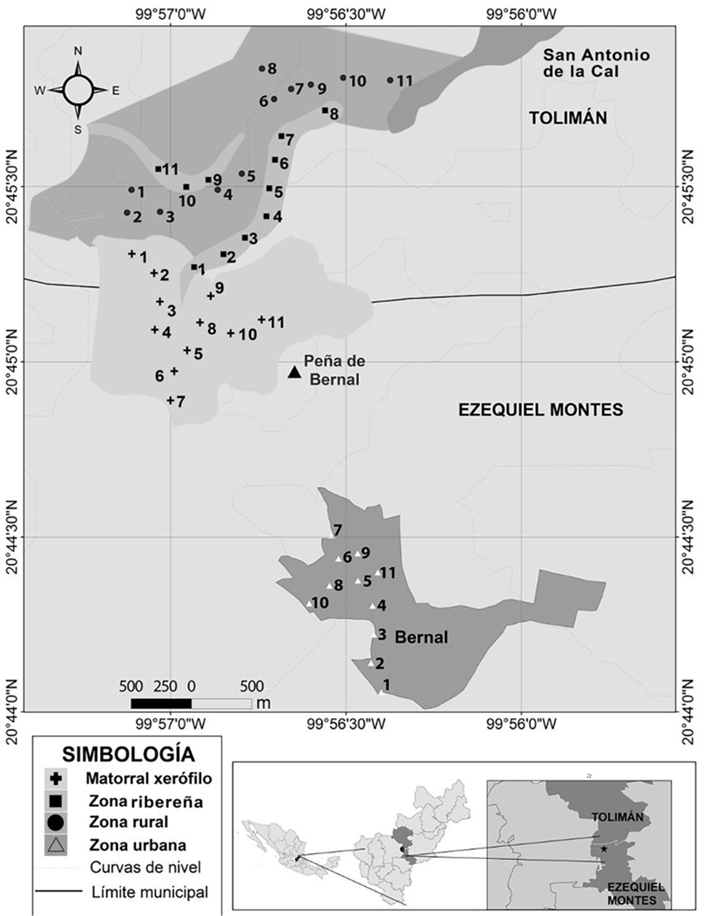
Figura 1 Se muestran los puntos de conteo para cada tipo de ambiente en Bernal y San Antonio de la Cal (arriba). Ubicación del área de estudio, estado de Querétaro y los municipios de Tolimán y Ezequiel Montes, México (abajo).
El área presenta climas secos y semisecos, y pertenece a la región del semidesierto queretano, tiene una precipi tación media anual de entre 350-650 mm. Esta zona presenta un relieve poco ondulado a plano con altitudes menores a los 2000 msnm, y se encuentra delimitada por sierras, mesetas y lomeríos del Eje Neovolcánico y de la Sierra Madre Oriental, que impide el paso de los vientos húmedos provenientes del Golfo de México (Segob 2015).
Para realizar el estudio elegimos los siguientes cuatro ambientes: 1) Matorral xerófilo, en la ladera norte del monolito. El estrato superior en este sitio no rebasa los 4 m de altura, y está dominado por Prosopis laevigata, Acacia farnesiana, A. shaffneri, Myrtillocactus geometrizans; además están presentes algunos elementos de selva baja caducifolia, como Bursera fagaroides y Celtis caudata. En su estrato medio se encuentran algunas especies arbustivas como Zaluziana au gusta, Karwinskia humboldtiana y Mimosa biuncifera (Zamudio et al. 1992). En este ambiente, al ser una propiedad privada con uso recreativo, sólo existen algunas veredas con acceso limitado y no hay actividades agropecuarias, edificaciones ni paso de vehículos automotores, 2) Ambiente ribereño en San Antonio de la Cal. Es un cauce estacional con árboles y arbustos nativos del área como Prosopis sp., Acacia sp., Ipomoea sp., Opuntia sp., e introducidos como Schinus molle y Eucalyptus sp. Este sitio colinda con algunas edificaciones distanciadas entre sí, algunas veredas, áreas de cultivo y áreas con vegetación seminatural, 3) Ambiente rural de San Antonio de la Cal. Presenta edificaciones separadas entre sí de manera irregular, las cuales representan menos del 10% del área de un paisaje ocupado mayormente por huertos, áreas agropecuarias, caminos de terracería, bardas vegetales y de piedra, remanentes de vegetación nativa, vegetación exótica (plantas ornamentales, Eucalyptus sp.), cableado escaso y presencia humana moderada y 4) Ambiente urbano en Bernal, presenta más del 80% de su superficie con edificaciones y calles pavimentadas con cableado. Este sitio también presenta algunos terrenos baldíos, escasa vegetación nativa, árboles principalmente exóticos (como Eucalyptus sp. y Ficus benjamina), algunos huertos de traspatio, así como constante afluencia de vehículos y frecuente presencia humana al ser esta ciudad un destino turístico importante.
Trabajo de campo
Utilizamos el método de conteo por puntos con radio fijo. Los datos obtenidos por este método permiten estudiar las diferentes composiciones de especies según el tipo de hábitat y los patrones de abundancia de cada especie (Ralph et al. 1996). En el área de estudio ubicamos 44 puntos, 11 en cada uno de los ambientes. Para ubicar los puntos y garantizar su accesibilidad utilizamos veredas en el matorral, caminos de terracería en la zona rural (excepto un punto), el cauce o veredas en la zona ribereña, y las calles de mayor longitud en Bernal. En estas vías colocamos los puntos en sitios que permitieran una visibilidad amplia y con una separación mínima de 150 m entre ellos. Tomamos en cuenta a las especies detectadas de manera visual o auditiva dentro de un radio de 30 m a partir del punto, durante un periodo de 10 min. Llevamos a cabo los muestreos durante las primeras tres horas después del amanecer durante los meses de diciembre de 2014 a abril de 2015. Para aumentar la detección de aves en cada punto el registro de datos lo realizamos dos personas que observamos en diferentes direcciones y conjuntamos los datos en un listado, mantuvimos comunicación constante para no contar dos veces el mismo individuo. Para considerar toda la estación invernal y aumentar la eficiencia del muestreo visitamos en cuatro ocasiones cada uno de los puntos, con tres o cuatro semanas entre cada repetición. El orden de visita varió cada vez para minimizar el efecto de la hora en la presencia de las especies. Para observar e identificar las aves utilizamos binoculares Vortex 10 x 42, cámara fotográfica y las guías de campo de Howell y Webb (1995), y Sibley (2003). Los nombres científicos fueron de acuerdo con la “American Ornithological Society” (Chesser et al. 2018, 2019) y los nombres comunes de Escalante et al. (2014). Para asignar un estado migratorio a las especies en la zona utilizamos los mapas de Howell y Webb (1995), y para el endemismo nos basamos en Berlanga et al. (2017).
En cada punto de muestreo hicimos una caracterización general de la vegetación de acuerdo con las indicaciones de Ralph et al. (1996). Esto es, dentro del radio de 30 m de cada punto estimamos visualmente el porcentaje de cobertura para los estratos herbáceo, arbustivo y arbóreo, para lo cual utilizamos la siguiente escala de valores: 1 < 5%, 2 entre 5 y 25%, 3 entre 25 y 50%, 4 entre 50 y 75%, y 5 entre 75 y 100%. Asimismo, la altura máxima la medimos en arbustos mediante una cinta métrica, y la estimamos para los árboles por comparación visual con una altura conocida. En zonas urbanas estimamos el porcentaje de cobertura de pavimento y construcciones, para lo cual utilizamos la escala de valores ya señalada para la cobertura vegetal. Adicionalmente, en el ambiente urbano medimos la distancia entre los puntos de muestreo y la periferia de la mancha urbana, y en los ambientes ribereño y matorral medimos la distancia entre el punto y la construcción más cercana. Para estas mediciones utilizamos imágenes satelitales de Google Earth de 2008.
Análisis de datos
Para agrupar los datos de las cuatro repeticiones en cada punto y no contar dos veces a un mismo individuo con el fin de evitar pseudoréplicas, consideramos sólo la abundancia máxima de cada especie en alguna de las repeticiones de cada uno de los puntos (Hurlbert 1984). Esto debido a que no hay seguridad de que los individuos vistos en cada repetición sean diferentes al no ser posible marcarlos, y no es recomendable sumarlos ni promediarlos puesto que en el primer caso se estaría posiblemente sobreestimando la abundancia por un conteo repetido de individuos, y en el segundo se subestimaría debido a que la abundancia máxima observada en una repetición es la abundancia mínima de la población, ya que no se conoce si hay individuos diferentes de las otras repeticiones (Johnson 2008). Para evaluar la eficiencia del muestreo utilizamos el estimador no paramétrico Chao 2, ya que representa el estimador menos sesgado para muestras pequeñas (Colwell y Coddington 1994). Este estimador basa su cálculo en datos de incidencia y toma en cuenta la distribución de las especies entre las muestras, y considera las especies observadas en exactamente una y dos unidades de muestreo (Moreno 2001).
Para comparar la riqueza y diversidad de especies entre los diferentes ambientes utilizamos los órdenes de diversidad de los números de Hill (números efectivos de especies), en específico el orden de diversidad cero (0D) que no considera la abundancia y corresponde a la riqueza de especies, así como los órdenes de diversidad uno (1D) y dos (2D), que respectivamente estiman los números efectivos de especies de acuerdo con su abundancia neta o dando un mayor peso a las abundancias de las especies dominantes (Hill 1973, Jost 2006). El análisis se llevó a cabo a una misma cobertura de muestra (0.97) en el programa inext, que utiliza interpolaciones y extrapolaciones de los datos (Chao et al. 2016). Se utilizaron intervalos de confianza del 84%, que son apropiados para inferencias equivalentes a p < 0.05 (Payton et al. 2004).
Para comparar visualmente la composición de especies entre los ambientes utilizamos gráficas de escalamiento multidimensional no métrico (NMDS, por sus siglas inglés), que presentan la diferencia entre objetos en un espacio bi-dimensional; para su elaboración utilizamos el índice de Jaccard, que se basa en valores de presencia-ausencia, y el índice de Bray Curtis, el cual toma en cuenta la abundancia relativa de las especies (Gauch 1982). Posteriormente, para estimar la significancia de las diferencias observadas entre el conjunto de los ambientes, aplicamos la prueba no paramétrica de análisis de similitud o anosim, la cual en una segunda prueba compara todos los hábitats entre sí utilizando la corrección de Bonferroni para disminuir el riesgo de falsas diferencias significativas (Clarke 1993). Adicionalmente, realizamos la prueba simper (porcentaje de similitud), para evaluar qué taxones son los principales responsables de la diferencia observada entre los grupos de muestras (Clarke 1993). Estos análisis los realizamos con ayuda del programa PAST 2.17 (Hammer et al. 2001).
Para comparar la diferencia en composición de especies (diversidad beta) entre los ambientes estudiados y entre los puntos utilizados (diferencia interna a los ambientes), utilizamos un análisis de partición de la diversidad beta (Veech y Crist 2009) mediante el siguiente modelo aditivo: y = α + (βpuntos) + (βambientes); donde α es la diversidad alfa (riqueza de especies) promedio en los puntos, y βpuntos y βambientes son la diversidad beta debida a la diferencia de composición de especies entre puntos y entre ambientes, respectivamente. Con la suma de estos dos valores se obtiene la diversidad beta total. En los modelos aditivos de la diversidad, como el utilizado, todos los valores obtenidos tienen la misma unidad (especies) y pueden compararse directamente en escalas espaciales o temporales (Crist et al. 2003). Este análisis lo llevamos a cabo con el programa partition 3 (Veech y Crist 2009).
Para analizar las posibles asociaciones de la composición de especies con atributos ambientales realizamos un análisis canónico de correspondencias (ACC), que es un método de análisis multivariante que permite relacionar la matriz de especies con otros datos, como variables del entorno (Ter Braak 1986). Los atributos ambientales considerados en todos los casos fueron la cobertura para los estratos herbáceo, arbustivo y arbóreo, así como la altura máxima de arbustos y árboles. Adicionalmente para el conjunto ma torral-ribereño se consideró la distancia a las construcciones, y para el urbano la cobertura de pavimento y construcciones y la distancia a la periferia de la mancha urbana. Este análisis lo realizamos con ayuda del programa past 2.17 (Hammer et al. 2001), analizando por separado los ambientes debido a las amplias diferencias en sus atributos, excepto para el ribereño y el matorral que se analizaron juntos.
Resultados
En total identificamos 64 especies en todos los ambientes estudiados (Cuadro 1), distribuidas en 24 familias y 53 géneros. Las familias Tyrannidae, Parulidae, Emberizidae e Icteridae fueron las que presentaron mayor riqueza de especies, con siete cada una. Del total de especies, 69.3% son residentes todo el año (cinco son especies exóticas), 25.8% residentes de invierno, 3.2% residentes de verano y 1.6% migratorias de paso. No detectamos ninguna especie con estatus de riesgo en la nom-059-semarnat-2010, pero sí especies en alguna categoría de endemismo: una endémica, seis semiendémicas y una cuasiendémica (Cuadro 1). La única especie endémica, Melanotis caerulescens (mulato azul), la observamos en todos los sitios salvo en el urbano, que presentó la menor riqueza de endemismos. Por otro lado, observamos una mayor riqueza y abundancia de especies exóticas en los ambientes urbano y rural, mientras que estuvieron ausentes en el matorral. Por su parte, las especies endémicas y cuasiendémicas presentaron una situación contraria, ya que se concentraron en los ambientes menos alterados y estuvieron ausentes en el área urbana. (Cuadro 2).
Cuadro 1 Listado taxonómico de las especies de aves registradas durante el invierno e inicio de la primavera del 2014-2015 en Bernal y San Antonio de la Cal, Querétaro. Se indica su clave, estatus migratorio (RP: Residente permanente, Ri: Residente de invierno, Rv: Residente de verano, Mp: Migratorio de paso), endemismo (Endem., SE: semiendémica, CE: cuasiendémica, En: endémica), y condición exótica (Exo).
| Orden | Familia | Especie | Nombre común | Clave | Estatus Migratorio | Endem. |
|---|---|---|---|---|---|---|
| Columbiformes | Columbidae | Columba livia | paloma doméstica | CL | RP | Exo |
| Streptopelia decaocto | tórtola turca | SD | RP | Exo | ||
| Columbina inca | tórtola cola larga | CN | RP | |||
| Zenaida asiatica | paloma ala blanca | ZA | RP | |||
| Apodiformes | Trochilidae | Selasphorus rufus | zumbador rufo | SR | Ri | |
| Cynanthus latirostris | colibrí pico ancho | CY | RP | SE | ||
| Amazilia violiceps | colibrí corona violeta | AV | RP | SE | ||
| Pelecaniformes | Ardeidae | Bubulcus ibis | garza ganadera | BI | RP | Exo |
| Piciformes | Picidae | Melanerpes aurifrons | carpintero cheje | MA | RP | |
| Dryobates scalaris | carpintero mexicano | PS | RP | |||
| Passeriformes | Tityridae | Pachyramphus aglaiae | mosquero-cabezón degollado | PA | RP | |
| Tyrannidae | Camptostoma imberbe | mosquero lampiño | CI | RP | ||
| Myiarchus cinerascens | papamoscas cenizo | MY | Rv | |||
| Contopus sordidulus | pibí occidental | CS | Rv | |||
| Empidonax sp. | mosquero | EM | - | |||
| Sayornis nigricans | papamoscas negro | SN | RP | |||
| Sayornis saya | papamoscas llanero | SS | RP | |||
| Pyrocephalus rubinus | mosquero cardenal | PY | RP | |||
| Vireonidae | Vireo cassinii | vireo de cassin | VC | Ri | SE | |
| Corvidae | Corvus corax | cuervo común | CC | RP | ||
| Hirundinidae | Hirundo rustica | golondrina tijereta | HR | RP | ||
| Remizidae | Auriparus flaviceps | baloncillo | AF | RP | ||
| Aegithalidae | Psaltriparus minimus | sastrecillo | PU | RP | ||
| Troglodytidae | Catherpes mexicanus | chivirín barranqueño | CM | RP | ||
| Troglodytes aedon | chivirín saltapared | TA | Ri | |||
| Thryomanes bewickii | chivirín cola oscura | TB | RP | |||
| Campylorhynchus brunneicapillus | matraca del desierto | CB | RP | |||
| Polioptilidae | Polioptila caerulea | perlita azul gris | PO | RP | ||
| Regulidae | Regulus calendula | reyezuelo de rojo | RC | Ri | ||
| Turdidae | Catharus guttatus | zorzal cola rufa | CG | Ri | ||
| Mimidae | Melanotis caerulescens | mulato azul | MC | RP | En | |
| Toxostoma curvirostre | cuitlacoche pico curvo | TC | RP | |||
| Mimus polyglottos | centzontle norteño | MP | RP | |||
| Ptiliogonatidae | Phainopepla nitens | capulinero negro | PN | RP | ||
| Passeridae | Passer domesticus | gorrión casero | PD | RP | Exo | |
| Fringillidae | Haemorhous mexicanus | pinzón mexicano | HM | RP | ||
| Spinus psaltria | jilguero dominico | SP | RP | |||
| Passerellidae | Chondestes grammacus | gorrión arlequín | CR | Ri | ||
| Spizella passerina | gorrión ceja blanca | SI | RP | |||
| Spizella atrogularis | gorrión barba negra | SA | RP | |||
| Melospiza lincolnii | gorrión de Lincoln | ML | Ri | |||
| Melozone fusca | toquí pardo | MF | RP | |||
| Pipilo maculatus | toquí pinto | PI | RP | |||
| Icteridae | Icterus wagleri | bolsero de Wagler | IW | RP | ||
| Icterus cucullatus | bolsero encapuchado | IC | Mp | SE | ||
| Icterus bullockii | bolsero calandria | IB | RP | SE | ||
| Icterus galbula | bolsero de Baltimore | IG | Ri | |||
| Molothrus aeneus | tordo ojo rojo | ME | RP | |||
| Molothrus ater | tordo cabeza café | MT | RP | |||
| Quiscalus mexicanus | zanate mexicano | QM | RP | Exo | ||
| Parulidae | Parkesia sp. | chipe | PR | - | ||
| Leiothlypis celata | chipe corona anaranjada | OC | Ri | |||
| Leiothlypis ruficapilla | chipe de coronilla | OR | Ri | |||
| Setophaga coronata | chipe coronado | SC | Ri | |||
| Setophaga townsendi | chipe negro amarillo | SE | Ri | |||
| Basileuterus rufifrons | chipe gorra rufa | BR | RP | CE | ||
| Cardellina pusilla | chipe corona negra | CP | Ri | |||
| Cardinalidae | Piranga flava | tángara encinera | PF | RP | ||
| Piranga rubra | tángara roja | PR | Ri | |||
| Piranga ludoviciana | tángara capucha roja | PL | Ri | |||
| Pheucticus melanocephalus | picogordo tigrillo | PM | RP | SE | ||
| Passerina cyanea | colorín azul | PC | Ri | |||
| Passerina versicolor | colorín morado | PV | RP | |||
| Thraupidae | Sporophila torqueola | semillero de collar | ST | RP |
Cuadro 2 Riqueza y abundancia (entre paréntesis) de especies de aves para los ambientes estudiados durante el invierno e inicio de la primavera de 2014-2015 en Bernal y San Antonio de la Cal, Querétaro, incluidos estado migratorio, endemismo y condición de exótica.
| Ambiente | Residentes | Migratorias | Endémicas- Cuasiendémicas | Semi- endémicas | Exóticas | Total |
|---|---|---|---|---|---|---|
| Matorral | 29 (158) | 7 (29) | 2 (9) | 4 (17) | 0 (0) | 187 |
| Ribereño | 30 (166) | 8 (40) | 2 (9) | 4 (21) | 1 (8) | 214 |
| Rural | 31 (310) | 11 (84) | 1 (1) | 5 (31) | 4 (92) | 486 |
| Urbano | 24 (255) | 7 (24) | 0 | 3 (10) | 4 (350) | 629 |
La completitud del inventario general fue de 89%, y por ambiente fue mayor al 86%, excepto para el matorral que fue de 76.3%. Los ambientes de matorral, ribereño y rural no tuvieron diferencia estadística en la riqueza de especies, sólo el área urbana presentó una menor riqueza (Figura 2) ligada a una mayor abundancia debida principalmente a especies exóticas. Las especies residentes nativas siguieron este mismo patrón en la riqueza, pero su abundancia fue mayor en el ambiente rural, seguido del ambiente urbano;
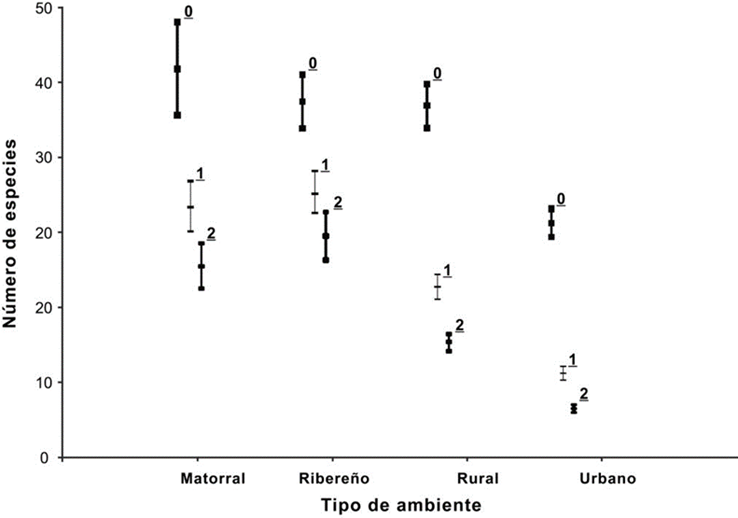
Figura 2 Diversidad de aves en Bernal y San Antonio de la Cal, Querétaro, en cuatro ambientes estudiados durante el invierno e inicios de primavera 2014-2015. Los números sobre las líneas indican los órdenes de diversidad cero (0D), uno (1D), y dos (2D), de acuerdo con Jost (2006). Se indican con barras horizontales el promedio y los intervalos de confianza del 84%. Todos los valores se tomaron a una misma cobertura de muestra (0.97) de acuerdo con Chao et al. (2016).
mientras que las especies migratorias (incluyendo semiendémicas) presentaron una mayor riqueza y abundancia en el ambiente rural (Cuadro 2). En cuanto a los órdenes de diversidad uno y dos, no encontramos una diferencia entre los ambientes matorral y ribereño, mientras que los ambientes rural y urbano tuvieron una menor diversidad, especialmente el urbano (Figura 2).
Las gráficas NMDS muestran un cambio gradual en la composición de especies en los ambientes. De izquierda a derecha los ambientes se sitúan en orden de su alteración ambiental: matorral, ribereño, rural y urbano (Figura 3). Así, los puntos de conteo que se ubicaron a la izquierda de la gráfica NMDS son del matorral y estaban alejados de cualquier indicio de urbanización, los puntos del ambiente ribereño se encuentran más a la derecha y presentaban una alteración relativamente mayor (p. ej. árboles exóticos, cercanía a construcciones o cultivos), al igual que en el ambiente rural donde el punto que se ubicó más a la derecha de la gráfica (11) presentó una mayor alteración debido a su cercanía a la zona urbana de San Antonio de la Cal. Al considerar sólo la incidencia de las especies mediante el índice de Jaccard, los tres primeros ambientes mencionados presentaron mayor similitud entre sí, mientras que el urbano se encuentra separado, lo cual indica una mayor diferencia en la composición de especies en este ambiente (Figura 3A). Al considerar la abundancia de las especies, mediante el índice de Bray-Curtis, el ambiente rural presentó una mayor separación con respecto al matorral y al ribereño, y un traslape mayor con el ambiente urbano (Figura 3B). La prueba de ANOSIM indicó que sí existe una diferencia estadística en la composición de especies entre los diferentes ambientes (R = 0.49; P = 0.0001). En la comparación entre pares de ambientes, encontramos que no existe diferencia significativa entre los ambientes matorral y ribereño, pero existe diferencia altamente significativa entre todos los demás pares posibles: matorral-rural, matorral-urbano, ribereño-rural, ribereño-urbano y rural-urbano (P = 0.0006).
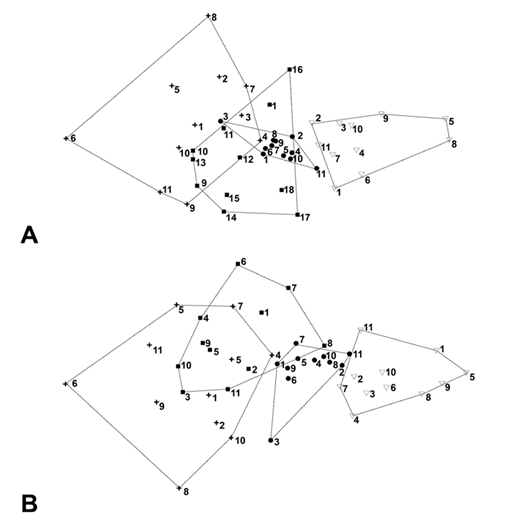
Figura 3 Escalamiento multidi-mensional no métrico (NMDS, por sus siglas inglés) de los puntos de conteo de aves realizados durante el invierno e inicio de primavera 2014-2015, en Bernal y San Antonio de la Cal, Querétaro. Se indican los puntos situados en el ambiente de matorral (cruces), ribereño (cuadrados), rural (círculos) y urbano (triángulos), con un número consecutivo de muestreo. A) Con base en el índice de Jaccard. B) Con base en el índice de Bray-Curtis.
En el análisis simper, para los pares de ambientes que sí presentaron diferencias significativas, encontramos en todos los casos que diez especies representaron más del 60% de la diferencia entre las áreas, en su mayor parte especies que tuvieron una mayor abundancia en la zona urbana o la rural. En general, Passer domesticus (gorrión casero) fue el principal factor de cambio, tanto para la zona urbana como para la rural. En ambos ambientes también fueron importantes Spizella passerina (gorrión ceja blanca), Haemorhous mexicanus (pinzón mexicano) y Setophaga coronata (chipe coronado), las dos especies últimas especialmente en la zona rural. Otras especies que fueron importantes para marcar la diferencia de la zona urbana fueron Quiscalus mexicanus (zanate mexicano), Columba livia (paloma doméstica) y Molothrus ater (tordo cabeza café); mientras que para el ambiente rural Zenaida asiatica (paloma ala blanca) fue también importante (Cuadro 3).
Cuadro 3 Principales especies de aves que representan el factor de cambio entre ambientes durante el invierno e inicio de la primavera de 2014-2015 en Bernal y San Antonio de la Cal, Querétaro. Se indica entre paréntesis el ambiente en el cual la especie tuvo mayor influencia (MA: matorral, RU: rural, RI: ribereño, UR: urbano). Las especies de aves se muestran como CB: Campylorhynchus brunneicapillus, CL: Columba livia, CN: Columbina inca, HM: Haemorhous mexicanus, ME: Molothrus aeneus, MF: Melozone fusca, MT: Molothrus ater, PD: Passer domesticus, PN: Phainopepla nitens, PY: Pyrocephalus rubinus, QM: Quiscalus mexicanus, SC: Setophaga coronata, SI: Spizella passerina, TC: Toxostoma curvirostre, ZA: Zenaida asiatica.
| MA-RU | % Acum. | MA-UR | % Acum. | RI-RU | % Acum. | RI-UR | % Acum. | RU-UR | % Acum. |
|---|---|---|---|---|---|---|---|---|---|
| PD (RU) | 14.13 | PD (UR) | 31.08 | PD (RU) | 14.24 | PD (UR) | 29.68 | PD (UR) | 19.67 |
| SC (RU) | 24.65 | QM (UR) | 39.53 | SC (RU) | 24.82 | QM (UR) | 38.01 | MT (UR) | 27.64 |
| HM (RU) | 33.21 | MT (UR) | 46.96 | HM (RU) | 33.66 | MT (UR) | 45.57 | SC (RU) | 35.1 |
| CN (RU) | 41.69 | SI (UR) | 52.02 | CN (RU) | 40.93 | SI (UR) | 50.58 | QM (UR) | 42.23 |
| ZA (RU) | 46.83 | HM (UR) | 56.13 | ZA (RU) | 45.62 | CL (UR) | 54.72 | CN (RU) | 48.92 |
| SI (RU) | 51.22 | CL (UR) | 60.22 | SI (RU) | 49.99 | HM(UR) | 58.81 | SI (UR) | 55.13 |
| PY (RU) | 55.37 | CB (MA) | 63.25 | PY (RU) | 53.92 | CN (UR) | 62.29 | HM (RU) | 61.21 |
| CB (MA) | 58.88 | CN (UR) | 66.24 | MF (RI) | 57.3 | MF (RI) | 64.77 | CL (UR) | 65.96 |
| TC (RU) | 61.97 | PN (MA) | 68.9 | CB (RU) | 60.09 | PY (UR) | 67.07 | ZA (RU) | 70.14 |
| PN (MA) | 65.03 | TC (MA) | 71.46 | ME (RU) | 62.74 | SC (RI) | 69.36 | TC (RU) | 72.69 |
El análisis de partición de la diversidad beta fue estadísticamente significativo (P < 0.005). La diversidad beta en los puntos fue ligeramente mayor que en los ambientes, de acuerdo con el siguiente modelo: 64 (γ) = 11.2 (α) + 26.6 (βpuntos) + 26.3 (βambientes).
La principal variable asociada a los cambios en composición de especies en los ambientes matorral y ribereño fue la distancia a las construcciones. En segundo lugar, aunque en sentido opuesto, fue la altura máxima arbórea (Figura 4A). Para el ambiente rural la variable visiblemente asociada al eje principal fue el porcentaje de cobertura vegetal, mientras que para el eje secundario fue el porcentaje de construcción; de manera adicional, se asociaron la altura máxima arbórea y la de arbustos (Figura 4B). En el ambiente urbano la variable más asociada al eje principal fue la altura máxima arbustiva, mientras que para el eje secundario fueron la altura máxima arbórea y la distancia a la periferia, y entre ambos ejes participó el porcentaje de construcción (Figura 4C).

Figura 4 Análisis Canónico de Corres pondencias en los ambientes matorral y ripario (A), rural (B) y urbano (C). Las principales variables asociadas a la composición de especies se muestran con líneas, donde AM: altura máxima arbórea, AMA: altura máxima arbustiva, CC: porcentaje de cobertura de pavimento y construcciones, CN: distancia a las construcciones, CV: porcentaje de cobertura vegetal, DP: distancia a la periferia. Las especies de aves se muestran con abreviaturas, donde AF: Auriparus flaviceps, AV: Amazilia violiceps, BI: Bubulcus ibis, BR: Basileu terus rufifrons, CB: Campylorhynchus brunneicapillus, CC: Corvus corax, CG: Catharus guttatus, CI: Camptostoma im berbe, CL: Columba livia, CM: Cather pes mexicanus, CN: Columbina inca, CP: Cardellina pusilla, CR: Chondestes grammacus, CS: Contopus sordidulus, CY: Cynanthus latirostris, EM: Empi donax sp., HR: Hirundo rustica, HM: Haemorhous mexicanus, IB: Icterus bullockii, IC: Icter cucullatus, IG: Ic terus galbula, IW: Icterus wagleri, MA: Melanerpes aurifrons, MC: Melanotis caerulescens, ME: Molothrus aeneus, MF: Melozone fusca, ML: Melospiza lincolnii, MP: Mimus polyglottos, MT: Molothrus ater, MY: Myiarchus cine rascens, OC: Oreothlypis celata, OR: Oreothlypis ruficapilla, PA: Pachyram phus aglaiae, PC: Passerina cyanea, PD: Passer domesticus, PF: Piranga fla va, PI: Pipilo maculatus, PK: Parkesia sp., PL: Piranga ludoviciana, PM: Pheucti cus melanocephalus, PN: Phainope pla nitens, PO: Polioptila caerulea, PR: Piranga rubra, PS: Dryobates scalaris, PU: Psaltriparus minimus, PV: Passerina versicolor, PY: Pyrocephalus rubinus, QM: Quiscalus mexicanus, RC: Regulus calendula, SA: Spizella atrogularis, SC: Setophaga coronata, SD: Streptopelia decaocto, SE: Setophaga townsendi, SI: Spizella passerina SN: Sayornis ni gricans, SP: Spinus psaltria, SR: Selas phorus rufus, SS: Sayornis saya, ST: Sporophila torqueola, TA: Troglodytes aedon, TB: Thryomanes bewickii, TC: Toxostoma curvirostre, VC: Vireo cassi nii, ZA: Zenaida asiatica.
Discusión
La completitud general del inventario, que fue > 80%, indica un muestreo adecuado, y que es probable la presencia de más especies que no fueron detectadas, especialmente en el matorral, ambiente con menor completitud. La menor riqueza observada en el ambiente urbano indica que la mayor urbanización de Bernal, a pesar de ser un asentamiento pequeño, ha reducido la riqueza de especies de aves, como generalmente ocurre en estos ambientes (Chace y Walsh 2006), y como ha sido observado en asentamientos urbanizados mexicanos de tamaño reducido (MacGregor-Fors y Schondube 2012). Esta reducción de especies en el ambiente urbano fue acompañada por una marcada mayor abundancia de las especies exóticas Streptopelia decaocto (tórtola turca), Columba livia, Passer domesticus y Quiscalus mexicanus, como ha sido registrado para este tipo de especies en muchas otras ciudades (Robbins et al. 1989, White et al. 2005, Chace y Walsh 2006).
Contrario a lo esperado, no encontramos diferencia en la riqueza de especies entre los ambientes menos alterados (matorral y ribereño) con el rural. En este aspecto, la predicción era encontrar una mayor riqueza en el ambiente rural, debido a que en él o en la interfase rural-urbana se ha reportado la riqueza más alta en estudios de gradientes urbanos (Blair 1996, Blair et al. 2001), debido posiblemente a que una urbanización moderada da lugar a una mayor hete rogeneidad ambiental y, por ende, a un incremento en la disponibilidad de recursos adicionales como sitios de anidación, fuentes permanentes de agua, y alimento de fácil acceso y con un alto aporte de energía, entre otros, que promueve una mayor cohabitación de especies (Crooks et al. 2004, Bock et al. 2008). De hecho, en otras zonas del estado de Querétaro se ha reportado que ambientes rurales mantienen un mayor número de especies que ambientes naturales (Hiley et al. 2016). Sin embargo, a pesar de no haber encontrado una mayor riqueza de especies en la zona rural, en este ambiente encontramos la mayor abundancia de especies residentes nativas y la mayor riqueza y abundancia de especies migratorias. Así, nuestros resultados resaltan que las zonas rurales con remanentes de vegetación natural en una matriz agropecuaria heterogénea son de gran importancia para la conservación de la avifauna, un patrón señalado por otros estudios en México (Ramírez-Albores 2010, González-Valdivia et al. 2012). Es importante considerar en este punto que los ambientes rurales, aún ubicados en diferentes zonas bioclimáticas, presentan entre sí menores cambios en su composición de especies, lo que con lleva una reducción de la diversidad global y favorece la homogeneización biótica, por lo que la conservación de las aves no puede depender exclusivamente de estas zonas y es necesaria la conservación de la vegetación seminatural, que presenta una composición de especies diferente (Hiley et al. 2016, Vázquez-Reyes et al. 2017, Karp et al. 2018).
La diversidad (órdenes 1 y 2) fue menor en los sitios rural y urbano respecto de los otros sitios, debido a la mayor abundancia (dominancia) de especies como Passer domesticus, Setophaga coronata, Spizella passerina, Molothrus ater y Haemorhous mexicanus en la zona rural; y Columba livia, S. passerina, M. ater, P. domesticus, Quiscalus mexicanus y Columbina inca (tórtola cola larga) en el ambiente urbano. Este último fue el ambiente con menores valores en ambos órdenes de diversidad, lo cual está asociado también a su menor riqueza de especies.
El cambio gradual detectado en la composición de especies y su abundancia por ambiente, coincidente con el gradiente de alteración antrópica matorral-ribereño-rural-urbano, sugiere que en general las especies tienen diferentes grados de tolerancia a la alteración del medio, como se ha observado en otros estudios (Fraterrigo y Wiens 2005). En este sentido resalta el que las especies exóticas mostraron una alta preferencia por los ambientes alterados rural y urbano, las especies endémicas por los ambientes poco alterados matorral y ribereño, y otras especies residentes y migratorias tuvieron mayores abundancias en el medio rural. El cambio gradual encontrado en la composición de especies también sugiere que las comunidades responden a factores ambientales muy locales, como ha sido observado para la riqueza y la abundancia de aves en diferentes tipos de urbanización en la Ciudad de México (Ortega-Álvarez y MacGregor-Fors 2009).
Las especies que marcaron las principales diferencias entre los ambientes fueron especies exóticas ligadas a ambientes urbanizados, como ha sido reportado para P. domesticus, C. livia y Q. mexicanus; mientras que M. ater, que en los muestreos urbanos lo registramos sólo en las primeras horas de la mañana, es una especie nativa que muchas veces pernocta en las ciudades pero se alimenta en los campos (Howell y Webb 1995, Wehtje 2003, White et al. 2005). Por otra parte, cuatro especies no exóticas resaltaron para diferenciar la avifauna del ambiente rural: Haemorhous mexicanus, Zenaida asiatica, Columbina inca y Setophaga coronata, las tres primeras son residentes y la última es migratoria, lo cual explica en parte la mayor abundancia de especies residentes y migratorias encontrada en la zona rural.
La diferencia entre ensambles también estuvo determinada por las especies exclusivas a cada ambiente. Por ejemplo, la zona urbana presentó seis especies exclusivas: cinco residentes y la exótica Streptopelia decaocto, que es una especie que se observó por primera vez en la ciudad de Querétaro en el 2009 (Pineda-López y Malagamba 2011) y se encuentra en proceso de expansión en dicha ciudad (López-Puebla 2019) y posiblemente en la región. Las especies residentes exclusivas de la ciudad incluyeron a Sporophila torqueola (semillero de collar), Piranga rubra (tángara roja) y P. ludoviciana (tángara capucha roja), que en la zona semiárida del estado de Querétaro se observan frecuentemente dentro de zonas arboladas urbanas o ligadas a cuerpos de agua (obs. per.). La zona rural también presentó seis especies exclusivas, dos de ellas relacionadas con cultivos (Molothrus aeneus -tordo ojo rojo- y Bubulcus ibis -garza ganadera-), una a zonas arboladas (Piranga flava -tángara encinera-) y tres de matorrales y zonas abiertas (Troglodytes aedon -chivirín saltapared-, Selasphorus rufus -zumbador rufo- y Chondestes grammacus -gorrión arlequín-). Los ambientes de matorral y ribereño presentaron siete especies exclusivas, propias de matorral y zonas arboladas de acuerdo con Howell y Webb (1995): Camptostoma imberbe -mosquero lampiño-, Contopus sordidulus -pibí occidental-, Pipilo maculatus -toquí pinto-, Parkesia sp., Leiothlypis ruficapilla -chipe de coronilla-, Pachyramphus aglaiae -mosquero cabezón degollado- y Passerina versi color -colorín morado-.
La diversidad beta alta que encontramos es reflejo de una gran heterogeneidad en la composición de especies, tanto entre ambientes como dentro de ellos, ya que sus componentes (βpuntos y βambientes) presentaron valores muy semejantes y altos en comparación con la diversidad alfa. Y, también, confirma la importancia de los cuatro ambientes para la conservación de la diversidad de aves, debido a que cada uno aporta una composición de especies con diferencias importantes (Pereyra y Moreno 2013). Por ende, la idea de integrar zonas naturales con ambientes alterados en la conservación de las aves cobra relevancia, lo cual ha sido señalado en otros estudios de zonas agropecuarias de México y del resto de Mesoamérica (Harvey et al. 2008, Ramírez-Albores 2010, González-Valdivia et al. 2012, Hiley et al. 2016). Sin embargo, el ambiente urbano presentó menor riqueza de especies, especialmente en el caso de especies residentes y endémicas, y una mayor abundancia de especies exóticas que pueden afectar negativamente a especies nativas (Álvarez-Romero et al. 2008). Asimismo, entre otros aspectos desfavorables, en las zonas urbanas las aves pueden enfrentar una mayor depredación por fauna doméstica, principalmente gatos domésticos (Baker et al. 2005, Garrard et al. 2018), que en el caso de las zonas rurales mexicanas generalmente también son abundantes (obs. per.). No obstante, en nuestra zona de estudio, Bernal presentó mayor abundancia de especies nativas residentes res pecto de los ambientes de matorral y ribereño. Por lo tanto, es importante proponer y realizar acciones de manejo que reduzcan esos aspectos negativos para que Bernal y otras ciudades puedan funcionar como refugios de avifauna, ya que actualmente se contempla a nivel global la necesidad de integrar a las ciudades en la conservación de la biodiversidad (Aronson et al. 2017, Garrard et al. 2018).
El alto valor de la diversidad beta a nivel de puntos (βpuntos), sugiere una importante heterogeneidad ambiental en el interior de los diferentes ambientes estudiados. La heterogeneidad ambiental en Bernal se observó en algunos puntos ubicados en sitios con lotes baldíos, huertos, patios amplios, parques, o diferente arbolado en las calles. El análisis ACC mostró la importancia de 1) la altura máxima arbustiva, que estuvo presente principalmente en lotes baldíos y patios, lo cual apoya las observaciones de otros autores respecto a que parches pequeños de vegetación dentro de las ciudades son capaces de mantener una composición avifaunística diferente (Carbó-Ramírez y Zuria 2011), 2) la altura máxima arbórea, que correspondió a especies exóticas como el Eucaliptus, porque, aunque se encontraron en bajos números, pueden tener un valor ecológico para las aves (Martin et al. 2009), y de 3) la distancia a la periferia de la mancha urbana. Esto último concuerda con lo encontrado en la gráfica NMDS donde los puntos más parecidos al ambiente rural están hacia la periferia, y muestra que aun en un área urbana pequeña hay variaciones de las comunidades de aves de la periferia hacia el centro, como se ha observado en ciudades grandes (Chace y Walsh 2006, Batáry et al. 2018).
En el ambiente rural, la heterogeneidad ambiental se observó en la presencia irregular de huertos, casas, caminos, pequeños depósitos de agua, parches de vegetación nativa, zonas de cultivo y, en sólo un punto, por una calle de concreto. Al igual que en el ambiente urbano, en el análisis ACC, la principal característica que influyó en la composición de la avifauna fue la vegetación, si bien en este caso la variable más importante fue la cobertura vegetal, que se debió a manchones de vegetación seminatural y a huertos, ya que por la época que se realizó el estudio (invierno y época de secas) las zonas de cultivos estaban sin cobertura. En consecuencia, es importante mantener los manchones de vegetación en las zonas rurales como refugio de especies nativas, especialmente si son áreas grandes (Rueda-Hernández et al. 2015); aunque manchones pequeños son importantes para la biodiversidad en general (Lindenmayer 2019, Wintle et al. 2019). En el ambiente rural también fue importante el porcentaje de construcción, asociado a diferencias de tamaño en las casas y a la calle de concreto que presentó el punto rural más cercano a la zona urbanizada. Esto sugiere un efecto paulatino de la urbanización en la composición de especies, como ha sido observado en gradientes con baja densidad de construcciones (Fraterrigo y Wiens 2005).
En los ambientes ribereño y matorral, que no presentaron diferencias entre sí en su composición avifaunística, la heterogeneidad ambiental de los puntos se observó en su cercanía a casas y cultivos, y a la presencia de arbolado no nativo. En este caso, resaltó la importancia de la distancia a construcciones para la composición de la comunidad de aves, como se ha observado en otros estudios (Fraterrigo y Wiens 2005, Grajales 2009, Jacoboski et al. 2017). También fue relevante la altura máxima del arbolado, aunque en sentido casi opuesto a la distancia a las construcciones.
Esto se explica porque los eucaliptos son los árboles más altos del sitio y se encontraban cercanos a áreas habitadas. Nuestros resultados apoyan la propuesta de que los ambientes ribereños, aún en ambientes alterados, son importantes para la conservación de la avifauna (Domínguez-López y Ortega-Álvarez 2014).
En el presente estudio se muestra que, durante el invierno, las zonas urbanas pequeñas sí pueden promover una diferencia en la riqueza y composición de las aves, y que zonas rurales como la estudiada pueden mantener una riqueza alta de especies, incluidas especies residentes y endémicas, y una mayor abundancia de especies residentes. La alta diversidad beta documentada implica la necesidad de realizar acciones de conservación en el conjunto de ambientes, especialmente el mantenimiento de parches de vegetación en zonas urbanas y rurales, así como la conservación de los ambientes con menos impacto antrópico. Nuestros resultados apoyan la idea de realizar acciones de manejo para integrar a ambientes alterados en la conservación de la bio-diversidad. Finalmente, consideramos que Bernal y la zona rural de San Antonio de la Cal son representativas de pequeñas zonas urbanas y rurales del centro de México, por lo que estas sugerencias podrían ser tomadas para sitios con condiciones semejantes.

