











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
En Ecuador como en todo el Neotrópico la información sobre depredadores de nidos es escasa (Cockle et al. 2016, Salvador 2016). La depredación de nidos es el factor de mortalidad más importante para la mayoría de las especies de aves; aunque afecta la densidad de las poblaciones, es un agente principal de selección natural que influye en la ecología reproductiva y la historia natural de las aves (Ricklefs 1969, Angelstam 1986). La modificación y fragmentación del hábitat, debido principalmente a la expansión agropecuaria, han provocado cambios en los índices de depredación de muchas especies y de aves (Bayne y Hobson 1997, Zanette 2002), en especial en aquéllas con poblaciones sensibles y distribución restringida, como es el caso de algunos psitácidos amenazados, que anidan en cavidades, y que eventos de depredación de sus nidadas han sido poco documentados (Oren y Novaes 1986, Kyle 2007).
El perico de El Oro (Pyrrhura orcesi) es una especie endémica y amenazada bajo la categoría en peligro de extinción a nivel mundial (IUCN 2018). La especie se distribuye en una franja estrecha de bosque nublado (90 km de largo), en un rango altitudinal de 700 a 1,200 m en el suroccidente del Ecuador (Ridgely y Greenfield 2006). P. orcesi habita de manera indistinta en áreas boscosas como en pastos arbolados. La dieta del perico de El Oro se compone principalmente de frutos de árboles de Ficus spp, Cecropia reticula, Heliocarpus popayanensis e Ireartea deltoidea (Echeverría-Vaca y Garzón-Santomaro 2016).
Pyrrhura orcesi es una especie gregaria, que forma grupos de 3 a 17 individuos, con la presencia de una sola pareja reproductora (Naranjo-Saltos 2007). Los grupos muestran un comportamiento cooperativo. El cuidado parental involucra a todos los miembros del grupo, desde el juvenil de la temporada anterior, incluidos individuos de bajo rango, que realizan actividades de búsqueda de alimento, búsqueda de cavidades de anidación, protección del nido, incubación y alimentación de los pichones (Klauke et al. 2013). Esta especie anida en cavidades ubicadas en árboles, principalmente copales (Dacryodes peruvianum), palmas (Iratea deltoidea), tangaré (Carapa guianensis) y la bella María (Nectandra sp.). En estos nidos se ha observado que cada grupo de pericos tiene un máximo de dos juveniles y quizá la puesta normal sea de cuatro a seis huevos (Garzón Santomaro 2004, Garzón-Santomaro y Juiña 2007, Naranjo-Santos 2007).
Pyrrhura orcesi fue descrita en 1985 (Ridgely y Robbins 1988), pero no fue hasta 1997 que se registró el primer evento reproductivo de una pareja, en la Reserva Buenaventura (RB) (López-Lanus y Lowen 1999). Desde 2002 la población de P. orcesi ha sido estudiada en esta reserva, donde se ha realizado el monitoreo de la población y nidos naturales, radiotelemetría e implementación de nidos artificiales (Schaefer y Schimidt 2003, Garzón-Santomaro 2004, Garzón-Santomaro y Juiña 2007, Klauke et al. 2013, Klauke et al. 2014).
Pyrrhura orcesi interactúa en la RB con el tucanete lomirrojo Aulacorhynchus haematopygus. El tucanete lomirrojo tiene amplia distribución en los pisos subtropicales y templados de bosques húmedos desde el suroeste de Venezuela hasta el norte de Perú (Restall et al. 2006). En Ecuador ocurre al oeste de los Andes, entre los 500 a 2000 msnm, principalmente habita los bordes de bosques y vegetación secundaria (Ridgely y Greenfield 2006). A. haematopygus se mueve en parejas o en grupos pequeños en todos los estratos del bosque (Arias-Alzate et al. 2012). Esta especie, por lo regular, se alimenta de bayas, pequeños frutos, insectos, ranas, huevos y pichones de otras aves (Fieldsa y Krabbe 1990). Actualmente, la ecología trófica de la familia Ramphastidae todavía es poco conocida, aunque estudios recientes la posicionan entre las depredadoras más importantes de huevos y pichones de aves (Cockle et al. 2016, Menezes y Marini 2017).
La presente nota tiene el objetivo de reportar eventos depredatorios de nidos de P. orcesi por A. haematopygus en la RB (2002-2006). Exponemos también eventos de competencia de cavidades con otra especie de psitácidos y preferencias de hábitat de anidación del perico de El Oro. A partir de estas observaciones y sucesos se ha incrementado el conocimiento sobre la historia natural de P. orcesi, lo que ayuda a la implementación del sistema de nidos artificiales, estrategia para asegurar mayor éxito de anidación y conservación de esta especie.
Áreas de estudio
La RB se ubica en el suroeste del Ecuador (3°38’37.64” S, 79°44’59.72” W) en la provincia de El Oro (Figura 1). Pertenece al sistema ecológico bosque siempreverde estacional piemontano del Catamayo Alamor según el MAE (2013). La RB constituye una zona de transición de bosque nublado influenciado por los Andes del Sur, la región húmeda del Chocó y la región seca Tumbesina (Valencia et al. 1999). Presenta varios tipos de hábitats, sin embargo, prevalecen las áreas boscosas y los pastos arbolados (Figura 2). Las principales actividades antrópicas que son evidentes en las áreas circundantes son la ganadería y la minería a pequeña escala (González-Romero et al. 2019).
Figura 1 Ubicación del área de estudio en la Reserva Buenaventura, provincia de El Oro, Ecuador (mapa: Mateo Vega).

Métodos
Durante los meses de reproducción y anidación: octubre a febrero, de 2002 a 2006, y en periodos del día de mayor actividad: 5:30-12:00 h; 15:30-18:00 h realizamos búsquedas intensivas de nidos naturales de P. orcesi. Recorrimos sitios donde, previamente, los pericos habían mostrado mayor actividad de forrajeo, inspección de cavidades, cópulas u otros. Permanecimos durante dos a tres horas en los sitios de potencial anidación, hasta confirmar el regreso de los pericos al área. Analizamos e interpretamos los datos de forma descriptiva y fueron la base para la ubicación y colocación de nidos artificiales.
Resultados
El 7 de diciembre de 2002 encontramos el primer nido de P. orcesi, en un remanente boscoso a 50 m de distancia del borde de un pasto arbolado, dentro del tronco de un copal (Dacryodes peruviana), a seis metros de altura desde el suelo (Figura 3); la entrada a la cavidad presentaba un diámetro de siete centímetros, casi el mismo ancho escapular de los pericos. La vegetación presentaba abundante sotobosque y dosel de 20 m de altura, aproximadamente, con abundantes epífitas y musgos en sus ramas. Las familias de árboles más representativas fueron Rubiaceae, Arecaceae, Melastomataceae, Fabaceae, Lauraceae y Moraceae.

Figura 3 Primer nido encontrado del perico de El Oro Pyrrhura orcesi, el 7 de diciembre de 2002 en la Reserva Buenaventura, provincia de El Oro, Ecuador (foto: E. Naranjo-Saltos).
El primer día que monitoreamos el nido observamos a cuatro individuos de P. orcesi. Primero llegó un individuo y se introdujo al nido, dos horas después otros tres, que tras permanecer en el exterior 20 minutos y emitir constantes vocalizaciones de contacto, entraron a la cavidad uno por uno. Permanecieron en la cavidad 15 minutos, luego salieron sólo tres y volaron hacia el bosque. Desde que este grupo de P. orcesi encontró el nido, mantuvo un patrón regular de visitas: cuatro veces al día como máximo, y permanecía por un lapso de hasta 15 minutos dentro. La primera visita del grupo al nido se llevó a cabo alrededor de una hora después del alba, al parecer después de alimentarse; la segunda, ocurrió a mediodía; la tercera, después de tres horas, y por la tarde, aproximadamente una hora y media antes de la puesta del sol.
En 2003 continuamos con el monitoreo del nido, y el 11 de enero, a las 09:00 h, observamos una pareja de A. haematopygus que tras emitir constantes vocalizaciones se acercó al nido de P. orcesi, a la entrada de la cavidad, y cada individuo introdujo el pico y procedieron a picotear los bordes, aparentemente para ensanchar el orificio de entrada (Figura 4A). Esta actividad duró entre tres y cinco minutos sin emitir ningún sonido. Mientras tanto, dentro del nido, se encontraba un perico sin realizar ninguna vocalización. Enseguida llegó el resto del grupo de P.orcesi (tres individuos), mismos que fueron perseguidos y espantados por los A. haematopygus, por lo que permanecieron cerca, expectantes, sin intentar defender su nido. La pareja de A. haematopygus continuó toda la mañana ensanchando la entrada de la cavidad y a las 15:00 h abandonó el área del nido. Después de una hora, el grupo de P. orcesi ingresó muy tímidamente a la cavidad, con constantes vocalizaciones de contacto.
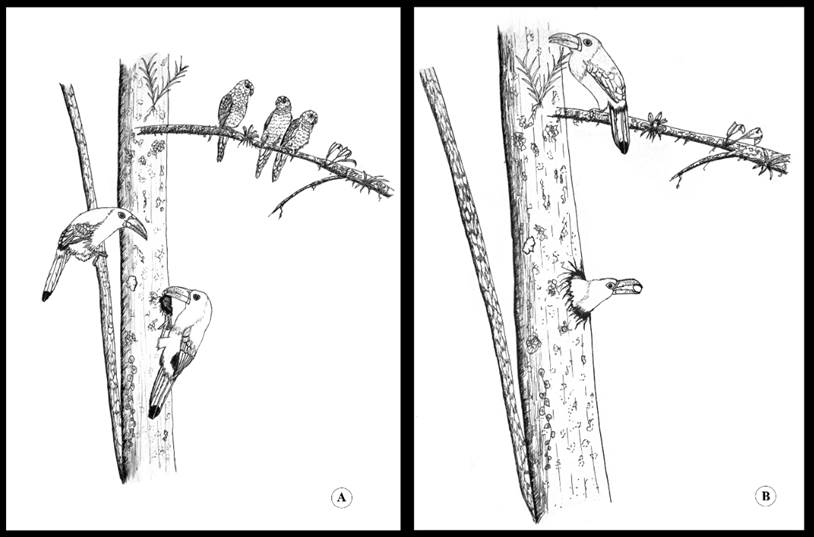
Figura 4 Aulacorhynchus haematopygus depredando un nido de Pyrrhura orcesi. (A) Ensanchamiento de la cavidad. (B) Depredación del nido (ilustración: G. Pozo-Zamora).
Al día siguiente, 12 de enero, a las 09:00 h, encontramos de nuevo a la pareja de A. haematopygus picoteando los bordes de la entrada y después ingresó al nido, alternadamente. La pareja se quedó algunos minutos dentro, emitiendo constantes vocalizaciones. Finalmente, los dos individuos volaron fuera del nido, uno llevaba consigo un huevo en el pico y se presume que el otro individuo voló con otro huevo (Figura 4B). Después de un par de minutos uno de los individuos regresó e ingresó a la cavidad, sin embargo, salió sin nada en el pico. En el lapso de la tarde ningún individuo de A. haematopygus ni P. orcesi regresaron al sitio, estos últimos posiblemente abandonaron el nido previo al saqueo. En años subsiguientes esta cavidad no fue utilizada por otros grupos de pericos.
En el periodo de estudio de 2002 a 2006 localizamos un total de 12 nidos naturales: cuatro tuvieron éxito (33%) con la salida de juveniles, cuatro más fueron depredados por A. haematopygus (33%), dos fueron desplazados por otros psitácidos (16%), y por último, dos fueron abandonados por causas desconocidas (16%). Entre 2002 y 2003 encontramos seis nidos naturales de P. orcesi, dos ubicados dentro del bosque y depredados por A. haematopygus, mientras que cuatro nidos fueron localizados cerca de los bordes de bosque y fueron exitosos. No evidenciamos el número de pichones de cada nido debido a la profundidad y a la altura en que se ubicaban (más de 7 m); sin embargo, ya avanzada la época de cría observamos que cada grupo de pericos anidantes volaba sólo con un juvenil. Entre 2005 y 2006 ubicamos seis nidos, de los cuales cuatro no fueron exitosos: dos fueron depredados por A. haematopygius (uno dentro del bosque y otro en pastos arbolados), e individuos de los otros dos nidos fueron desplazados por otras especies. Este último evento ocurrió en un pasto arbolado, un día después de que encontramos y confirmamos que eran nidos de P. orcesi. Hallamos que uno de los nidos estaba ocupado por el loro cachetirrosa (Pyrilia pulcra) y el otro por el perico caretirrojo (Psittacara erythrogenys). No presenciamos el momento de expulsión de los grupos de P. orcesi de sus nidos. En todos los nidos exitosos de este periodo observamos un juvenil en cada grupo.
El comportamiento de depredación por A. haematopygus fue similar en todos los eventos que observamos. Se avistó que el A. haematopygus persigue a los grupos de P. orcesi, hasta localizar el nido. Una vez que ubican el nido, los individuos de A. haematopygus esperan a que el grupo de P. orcesi salga de la cavidad, en ese momento, con vuelos cortos, se dirigen hacia el nido. Ensanchan el orificio de entrada, ingresan y saquean el nido (se llevan los huevos en el pico). Estos sucesos de depredación ocurren las primeras semanas de anidación de P. orcesi, en el periodo de incubación. Al pasar las semanas, las incursiones de los A. haematopygus a los nidos disminuyen, hasta desaparecer, lo cual coincide con la eclosión de los pichones. Nunca observamos que A. haematopygus depredara pichones.
A partir de los eventos de depredación que observamos en los nidos en el interior de los bosques, y el éxito de aquéllos ubicados en los pastos arbolados en la RB, en 2007 implementamos un sistema de nidos artificiales para aumentar la disponibilidad de cavidades como potenciales nidos para P. orcesi (Figura 5). Colocamos 50 nidos artificiales en los pastos arbolados cerca de los bordes boscosos en la RB. Hasta el periodo 2013-2014 los grupos de P. orcesi habían utilizado 34 nidos, ocupados por 282 individuos (Atti 2014). Ningún nido artificial ha sido depredado por tucanes, sin embargo, algunos fueron abandonados, posiblemente a causa de depredación por otros grupos de animales como mamíferos y reptiles (Klauke et al. 2014).

Discusión
Tres de los cuatro casos de depredación de nidos de P. orcesi ocurrieron en nidos localizados en el interior del bosque, mientras que los nidos exitosos estuvieron ubicados en pastos arbolados (4). Al parecer P. orcesi tiene mayores posibilidades de vigilancia en pastos arbolados, pues A. haematopygus es más visible fuera del bosque, por la incapacidad de esconderse o camuflarse eficazmente. Este comportamiento es similar al del perico dorado (Guaruba guarouba), que anida en árboles aislados de áreas abiertas, con lo cual evita la depredación de sus nidos (Oren y Novaes 1986, Silveira y Belmonte 2005, Laranjeiras 2011). Asimismo, estos registros son similares a otras familias, como el observado en México con el zacua mayor Psarocolius montezuma, en donde la ubicación de las colonias de anidación se encontraba en sitios abiertos y cerca de los bosques, lo que les da mayor oportunidad de vigilia tanto para depredadores como para aves parásitas (Nava-Solorio 1994). Sin embargo, otros factores pueden afectar el éxito reproductivo, ya que las colonias están más expuestas a perturbaciones naturales: viento, temperatura, y humanas: quema, deforestación, saqueo de nidos (Laranjeiras 2008).
Generalmente, A. haematopygus es una especie frugívora (Remsen et al. 1993), y así lo demostraron los contenidos estomacales: frutas y semillas, de dos especímenes de la colección de aves del Instituto Nacional de Biodiversidad del Ecuador (MECN 5899, MECN 2389). Sin embargo, se considera que A. haematopygus es una especie omnívora porque se alimenta de frutos, invertebrados y pequeñas presas en el dosel (Neotropical Birds 2018), así como de huevos y pichones de otras aves (Hilty y Brown 1986).
Aulacorhynchus haematopygus depreda únicamente huevos de P. orcesi y no los pichones, quizá por la incapacidad de tragarlos. Eventos similares están documentados en el bosque de Cruce Caballero, en Argentina, donde un arasari orejicastaño (Pteroglossus castanotis) saqueó un nido de carpintero lineado (Dryocopus lineatus) y solamente devoró los huevos y no al pichón, en apariencia porque los pichones eran demasiado grandes para tragarlos (Cockle et al. 2016).
Reportes en los que el género Aulacorhynchus funja como depredador son escasos. Por ejemplo, existen observaciones del tucanete esmeralda (Aulacorhynchus prasinus) en depredación de nidos de otras aves en Panamá (Wetmore 1968). Mientras que A. haematopygus sólo tiene un reporte conocido y proviene de la cordillera central de los Andes en Antioquia, Colombia, donde por medio de trampas cámara quedó registrado cuando depredaba huevos de tinamú grande (Tinamus major) (Arias-Alzate et al. 2012). En Ecuador muy poco se sabe de estos eventos, tan sólo los observados en la Reserva Buenaventura en el Proyecto de Conservación del Perico de El Oro y que originaron este estudio (Garzón-Santomaro 2004, Naranjo-Saltos 2007).
Asimismo, pocos son los reportes de tucanes depredadores de nidos de la familia Psittacidae. Por ejemplo, los tucanes goliblanco (Ramphastus tucanus) y el piquicanalado (Ramphastus vitellinus) son depredadores de huevos y pichones del perico dorado Guaruba guarouba, especie endémica de Brasil (Oren y Novaes 1986). Kyle (2007) en una investigación del guacamayo barbazul (Ara glaucogularis), en Brasil, señala que de los siete nidos activos, tres de ellos fueron depredados principalmente por el toco tucán (Ramphastos toco).
Al contrario, existe numerosa información de eventos depredadores de la familia Ramphastidae hacia otras especies de aves. Cockle et al. (2016) en Argentina, en la Selva Atlántica, Misiones, reportan que de los 33 nidos depredados (25 especies) el 50% fueron efectuados por tucanes, sobre todo de los géneros Ramphastos y Pteroglossus. En Costa Rica, en el parque Corcovado individuos de Ramphastus ambiguus fueron observados devorando un pichón en un nido de tityra carirroja (Tityra semifasciata) (Stuart 2015). Menezes y Marini (2017) en una compilación para el Neotrópico citan 14 especies de tucanes como depredadores de más de 35 especies de aves; entre los principales están Ramphastos dicolorus, R. toco y R. tucanus.
La depredación de nidos es considerada un mecanismo en la selección natural y control de poblaciones de especies de aves (Ricklefs 1969, Angelstam 1986). Los nidos registrados de P. orcesi (2002 a 2006) que fueron depredados por A. haematopygus afectaron a un tercio de la nidada de la población de pericos registrada en la RB. Estos registros, junto con la información de la plasticidad del perico en la selección de hábitat para anidar, han ampliado el conocimiento en la ecología de esta especie. Tomando en cuenta estas variables de depredación y de selección de hábitat, más la experiencia adquirida en el estudio de P. orcesi, es que optamos por la implementación de nidos artificiales, los cuales en la actualidad son la principal herramienta de investigación del P. orcesi. La información sobre depredación de nidos se ha convertido en una herramienta de gran utilidad al momento de planificar estrategias de conservación (Salvador 2016).

