











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
La agrupación de las aves en gremios constituye un paso clave que contribuye al entendimiento de la estructura del ensamble y de los procesos biológicos que interfieren en su organización (López de Casanave et al. 2008). Un gremio puede ser definido como un subgrupo dentro de la comunidad en el cual los individuos utilizan tipos de recursos similares de una manera análoga, y por tanto las interacciones tienden a maximizarse (Root 1967). Esta definición constituye uno de los aspectos ecológicos más importantes para el estudio de un ensamble (Kirkconnell et al. 1992). La distribución de las especies en gremios no es un evento azaroso e indica cómo la evaluación de los ensambles de aves, sólo en términos de riqueza de especies y abundancia, puede simplificar complejos patrones comunitarios al ocultar importantes detalles (Wiens 1989). Por otra parte, la definición de las especies en cada gremio, permite analizar de forma más lógica las interacciones ecológicas que se llevan a cabo entre un pequeño subgrupo de especies dentro del ensamble (Lovette y Fitzpatrick 2016).
La mayoría de las clasificaciones de los gremios realizadas en el mundo están basadas en las características de la dieta (Wiens 1989). Sin embargo, se han concebido otros tipos de clasificaciones basadas en las dimensiones del área de forrajeo: la conducta de forrajeo, hábitat, residencia o sitios de nidificación (Howell 1971, Karr 1976, Emlen 1977). Según Willson (1974), los tres componentes principales para la definición de un gremio son el tipo de alimento, los sitios donde lo obtiene y la forma de tomarlo; aunque todas las especies pertenecientes a un mismo gremio no tienen por qué usar los recursos del ambiente de igual modo (Landres y MacMahon 1980, Acosta et al. 1994). A su vez, la clasificación de estos gremios puede efectuarse mediante dos formas diferentes (Jacksić 1981). La más general es la clasificación “a priori”, en la cual el investigador ubica a las especies en grupos según el tipo de alimento que utilizan, teniendo en cuenta para ello su experiencia de campo. Esta modalidad es la que se aprecia en las clasificaciones ofrecidas por Acosta et al. (1992) y Kirkconnell et al. (1992) para las aves cubanas. La otra clasificación, nominada “a posteriori” es, por ejemplo, la utilizada por Acosta et al. (1994) para aves acuáticas asociadas a humedales cubanos. En dicho trabajo, los autores utilizaron valores obtenidos de las mediciones de un grupo de variables relacionadas con la alimentación y evaluadas mediante un análisis de agrupamiento, que permitió determinar los gremios presentes y las especies incluidas en cada uno de ellos.
La Ciénaga de Zapata, además de ser un sitio Ramsar y Área de Importancia para las Aves (Aguilar 2010), constituye el humedal más importante de Cuba y el Caribe (Mugica et al. 2008). Dentro de la ciénaga se encuentra Las Salinas, la cual contiene el más importante sistema de lagunas costeras dentro del parque. Un estudio reciente evidencia la importancia para las aves acuáticas, ya que se han llegado a registrar hasta 64 especies (González et al. 2016). Dada que la estructura en gremios es esencial para entender el funcionamiento del ensamble de aves, y para dar continuidad al trabajo previo de variabilidad temporal (González et al. 2016), nuestro objetivo fue evaluar la dinámica temporal de la composición y estructura en gremios del ensamble de aves acuáticas durante los años 2012 y 2013.
Métodos
Área de estudio
Realizamos este estudio en el sistema de lagunas costeras de Las Salinas, (22°06’35” N y 81°16’58” O). La zona de estudio se encuentra ubicada a 1.5 km al norte de la estación ecológica de Las Salinas y a 12 km del sur del poblado de Buena Ventura; bordea la costa occidental de Bahía de Cochinos. Este sitio comprende un sistema de lagunas costeras interconectadas que varían en profundidad según la época del año, suelo con depósitos areno-arcillosos y órgano-detríticos (Soto et al. 1994). Además, se aprecian áreas con afloramientos rocosos que se hacen más o menos evidentes según el régimen de lluvias. Los valores de profundidad de agua de las ocho varas que están dispuestas a lo largo del recorrido arrojaron un valor promedio ± EE de 13.9 ± 2.4 cm durante la migración primaveral (febrero-marzo), 26.9 ± 2.3 cm en la residencia de verano (mayo-junio) y 39.8 ± 2.4 cm en migración otoñal (octubre-noviembre) de los años 2012 y 2013. Quisiéramos aclarar que estos valores corresponden a varas que están fijadas dentro del sistema lagunar de Las Salinas, las cuales en ocasiones no quedan incluidas dentro de los microhábitats utilizados por las diferentes especies que conforman el ensamble.
Conteo de aves
Los conteos de aves los realizamos en los 8 km finales de Las Salinas, donde se cuenta con un sistema lagunar a ambos lados del camino y cuya visibilidad permite la detección de los individuos presentes. Para el conteo de aves empleamos una combinación de los métodos de transecto y parcela circular, según lo recomendado por Acosta et al. (2013) para ecosistemas acuáticos. En el caso de los transectos, realizamos los conteos desde un vehículo a una velocidad de 10 km/h con un observador de cada lado. Siempre que fue necesario se hicieron paradas para facilitar la observación. En los dos métodos utilizados tuvimos en cuenta un radio de 500 m a cada lado del camino, por la amplia visibilidad del área y la posibilidad de medir la banda con el uso de un Laser Ranger Finder con 1,000 m de alcance efectivo.
Hicimos los conteos durante tres periodos de los años 2012 y 2013: migración primaveral (febrero-marzo, 2012: n = 6 y 2013: n = 5); residencia de verano (mayo-junio de ambos años, n = 5 en cada año) y migración otoñal (octubre-noviembre de ambos años, n = 6 en cada año). En todos los casos las observaciones las efectuamos en la sesión de la mañana (07:00-10:00 h) y para ello utilizamos binoculares de 10 x 42 y un telescopio de 60x. En casos necesarios para la identificación de las especies nos apoyamos en la guía Sibley (2000). Con vistas a disminuir los efectos del horario alternamos el inicio y fin del recorrido durante los días sucesivos de los conteos.
Análisis de los datos
Las especies que conformaron el ensamble de aves acuáticas las clasificamos en gremios. Para ello seguimos la clasificación de Acosta et al. (1994) donde se describieron para Cuba los gremios: zancudas, buscadores aéreos, buceadores, sondeadores profundos, sondeadores someros y vegetarianos. Acosta et al. (1994) identificaron estos gremios a partir de un análisis “a posteriori” donde tuvieron en cuenta para cada especie estudiada su dieta, características morfológicas y hábitos de forrajeo. En el presente estudio decidimos cambiar la nomenclatura del gremio vegetarianos descrito por Acosta et al. (1994) a herbívoro. Para clasificar algunas de las especies de aves acuáticas que no están incluidas en el trabajo de Acosta et al. (1994) adicionamos las categorías filtrador y omnívoros, las cuales están incluidas en otra clasificación de referencia en Cuba hecha por Kirkconnell et al. (1992). En estas dos categorías quedaron incluidos Phoenicopterus ruber (flamenco americano) y algunos representantes de la familia Rallidae, respectivamente. Todas las especies fueron de igual modo clasificadas según la categoría de permanencia propuesta por Llanes et al. (2002) en: residentes de invierno, residentes de verano, residentes bimodales (especies con poblaciones migratorias y poblaciones que crían en Cuba) y residentes permanentes. Con la utilización de ambas categorías se evaluó la riqueza específica por categoría de permanencia para cada uno de los gremios.
A cada gremio se le calculó un valor de densidad promedio (aves/km2) (± EE) por periodo (migración primaveral, residencia de verano y migración otoñal) y año de estudio (2012 y 2013). Dado que el presente estudio sólo incluyó dos años, y que los valores de densidad anual del ensamble pueden variar por factores inherentes al área de estudio; las comparaciones temporales de los gremios las hicimos a partir de sus valores de abundancia proporcional para cada año. Calculamos el valor de abundancia proporcional para cada periodo [Total de individuos del gremio i en el año k/Total de individuos pertenecientes al ensamble j en el año k*(100)] y lo consideramos como una medida cuantitativa de su representatividad o contribución. En un primer momento, comparamos los valores de abundancia proporcional por gremios entre años para los tres periodos. Para ello, calculamos la diferencia promedio de la abundancia proporcional por gremio entre años y ese valor de diferencia real lo comparamos con una distribución nula obtenida por métodos de remuestreo de Monte Carlo con 1,000 iteraciones. En cambio, para las comparaciones entre periodos, calculamos los intervalos de confianza de Monte Carlo al 95% con 1,000 iteraciones. Todos estos análisis los realizamos con el programa accesorio al Excel Pop Tools versión 3.0.5 (Hood 2008).
Por último, calculamos la representatividad de cada especie dentro de los gremios a través de la abundancia proporcional [total de individuos de la especie i/total de individuos pertenecientes al gremio j (100)]. Este valor fue calculado para los tres periodos de estudio. Para complementar el análisis de la representatividad de las especies por gremios calculamos además los valores de frecuencia de observación de las especies por gremio. La frecuencia la determinamos con la presencia/ausencia de las especies en los conteos realizados en cada periodo, sin importar el año. Los valores de abundancia proporcional de las especies también fueron utilizados como una medida de la contribución de las mismas al gremio al que pertenecen.
Resultados
Las aves acuáticas identificadas en Las Salinas fueron agrupadas en ocho gremios (Cuadro 1). La composición de cada gremio varió entre 1 y 15 especies. El gremio de mayor riqueza específica fue el de los sondeadores someros, seguido por el gremio de las zancudas y buscadores aéreos (Cuadro 2). En tanto, el gremio filtrador estuvo representado únicamente por P. ruber. Las categorías de permanencia variaron en representatividad para cada gremio. Por ejemplo, el gremio de los sondeadores someros estuvo compuesto mayormente (67%) por especies residentes invernales, en tanto en el gremio zancudas predominaron (92%) las especies residentes bimodales. Dentro del gremio buscadores aéreos se identificaron especies pertenecientes a las cuatro categorías de permanencia establecidas (Cuadro 2).
Cuadro 1 Valores promedio de densidad (número de individuos / km2) (± EE) de los gremios de aves acuáticas presentes en Las Salinas, Ciénaga de Zapata durante la migración primaveral (MP), residencia de verano (RV) y migración otoñal (MO) de los años 2012 y 2013.
| MP | RV | MO | ||||
|---|---|---|---|---|---|---|
| Gremios | 2012 | 2013 | 2012 | 2013 | 2012 | 2013 |
| tróficos/año | n=6 | n=5 | n=5 | n=5 | n=6 | n=6 |
| Sondeadores Someros | 14.8±3.8 | 45.9±12.0 | 2.4±0.6 | 2.4±0.7 | 7.9±3.5 | 0.9±0.4 |
| Zancudas | 20.0±4.8 | 20.2±3.2 | 4.0±0.5 | 6.6±1.6 | 16.1±2.9 | 5.5±0.4 |
| Buscadores Aéreos | 30.4±5.2 | 19.9±4.9 | 0.8±0.4 | 1.5±0.5 | 16.2±6.6 | 6.4±2.4 |
| Buceadores | 34.3±7.0 | 98.5±59.3 | 4.0±2.3 | 2.4±1.7 | 498.6±309.6 | 155.4±108.1 |
| Herbívoros | 4.9±3.0 | 23.2±14.3 | 0.4±0.2 | 0.7±0.2 | 893.4±449.8 | 579.2±157.0 |
| Sondeadores Profundos | 6.6±2.3 | 8.5±2.1 | 7.8±3.6 | 6.8±2.2 | 4.6±1.9 | 5.0±2.7 |
| Omnívoros | 0.04±0.04 | 0.2±0.1 | 0.2±0.1 | 0.2±0.1 | 0.1±0.1 | 0.1±0.02 |
| Filtradores | 55.1±4.1 | 53.0±30.2 | 57.1±4.7 | 45.4±13.4 | 47.6±10.0 | 48.4±10.5 |
Cuadro 2 Lista de las especies por gremio. Se incluye la categoría de estado de permanencia (EP); y para cada periodo (MP, RV y MO) la abundancia proporcional (Ab. Prop.) y la frecuencia de observación en 100% (Fr.). RB: residente bimodal, RI: residente de invierno, RP: residente permanente; RV: residente de verano, MP: migración primaveral; RV: residencia de verano y MO: migración otoñal. La n representa el número de conteos realizados en cada periodo.
| MP n = 11 | RV n = 10 | MO n = 12 | |||||
|---|---|---|---|---|---|---|---|
| Gremio /Especie | EP | Ab. Prop. | (Fr.) | Ab. Prop. | (Fr.) | Ab. Prop. | (Fr.) |
| SONDEADORES SOMEROS | |||||||
| Pluvialis squatarola | RB | 14.6 | (91) | 0.9 | (20) | 2.6 | (33) |
| Charadrius vociferus | RB | 0.9 | (20) | 4.1 | (42) | ||
| Charadrius semipalmatus | RI | 0.6 | (36) | 0.2 | (8) | ||
| Charadrius wilsonia | RV | 0.3 | (9) | 14.0 | (50) | ||
| Himantopus mexicanus | RB | 16.2 | (64) | 45.7 | (100) | 22.1 | (25) |
| Tringa melanoleuca | RI | 7.1 | (91) | 6.3 | (60) | 2.9 | (58) |
| Tringa flavipes | RI | 9.6 | (100) | 5.9 | (20) | 4.5 | (58) |
| Tringa semipalmata | RP | 6.8 | (91) | 9.1 | (80) | 1.0 | (17) |
| Arenaria interpres | RI | 1.7 | (73) | 0.9 | (10) | ||
| Actitis macularius | RI | 0.1 | (18) | 2.6 | (42) | ||
| Calidris minutilla | RI | 17.8 | (36) | ||||
| Calidris mauri | RI | 3.1 | (18) | ||||
| Calidris himantopus | RI | 0.3 | (9) | ||||
| Calidris alpina | RI | 0.2 | (18) | ||||
| Limnodromus griseus | RI | 21.6 | (82) | 16.3 | (70) | 59.5 | (25) |
| ZANCUDAS | |||||||
| Ardea herodias | RB | 16.5 | (100) | 7.4 | (90) | 14.0 | (100) |
| Ardea alba | RB | 32.3 | (91) | 2.0 | (60) | 25.6 | (100) |
| Egretta rufescens | RB | 14.9 | (100) | 40.6 | (100) | 5.4 | (100) |
| Egretta tricolor | RB | 11.4 | (100) | 15.9 | (90) | 18.4 | (100) |
| Egretta caerulea | RB | 13.9 | (100) | 4.8 | (70) | 11.6 | (100) |
| Egretta thula | RB | 7.5 | (100) | 24.5 | (90) | 15.8 | (100) |
| Bubulcus ibis | RB | 0.4 | (8) | ||||
| Butorides virescens | RB | 1.8 | (82) | 4.4 | (100) | 4.9 | (92) |
| Nycticorax nycticorax | RB | 0.1 | (8) | ||||
| Nyctanassa violacea | RB | 0.1 | (9) | 0.2 | (10) | 0.1 | (17) |
| Mycteria americana | RP | 1.6 | (55) | 3.8 | (92) | ||
| BUSCADORES AÉREOS | |||||||
| Pelecanus erythrorhynchos | RI | 12.6 | (91) | 35.2 | (40) | 12.6 | (50) |
| Pelecanus occidentalis | RB | 3.8 | (91) | 4.2 | (50) | ||
| Fregata magnificens | RB | 0.8 | (55) | 5.5 | (30) | 0.7 | (25) |
| Leucophaeus atricilla | RB | 0.4 | (36) | 1.9 | (42) | ||
| Hydroprogne caspia | RP | 64.3 | (100) | 18.7 | (40) | 61.5 | (92) |
| Thalasseus maximus | RB | 5.5 | (100) | 4.4 | (10) | 6.6 | (50) |
| Gelochelidon nilotica | RI | 2.0 | (73) | 6.6 | (40) | 0.1 | (8) |
| Sternula antillarum | RV | 29.7 | (80) | 0.1 | (8) | ||
| Rynchops niger | RI | 7.8 | (45) | ||||
| Megaceryle alcyon | RI | 3.0 | (100) | 12.3 | (92) | ||
| BUCEADORES | |||||||
| Tachybaptus dominicus | RP | ˂0.1 | (9) | ||||
| Podilymbus podiceps | RP | 0.5 | (55) | 0.23 | (50) | ||
| Phalacrocorax auritus | RB | 5.7 | (100) | 100 | (70) | 1.7 | (75) |
| Phalacrocorax brasilianus | RB | 0.8 | (58) | ||||
| Anhinga anhinga | RB | ˂0.1 | (17) | ||||
| Fulica americana | RB | 93.7 | (73) | 97.1 | (58) | ||
| Mergus serrator | RI | 0.1 | (27) | ||||
| Aythya affinis | RI | 0.1 | (9) | 0.2 | (42) | ||
| Aythya collaris | RI | ˂0.1 | (17) | ||||
| HERBÍVOROS | |||||||
| Spatula discors | RI | 90.0 | (55) | 19.5 | (10) | 98.5 | (92) |
| Spatula clypeata | RI | 6.6 | (18) | 0.5 | (67) | ||
| Mareca americana | RI | 3.5 | (18) | 1.0 | (42) | ||
| Anas acuta | RI | ˂0.1 | (17) | ||||
| Dendrocygna arborea | RP | 80.5 | (60) | ||||
| OMNÍVOROS | |||||||
| Rallus longirostris | RP | 100 | (46) | 100 | (50) | 72.2 | (50) |
| Gallinula chloropus | RB | 27.3 | (8) | ||||
| SONDEADORES PROFUNDOS | |||||||
| Eudocimus albus | RP | 76.9 | (100) | 34.0 | (100) | 69.8 | (100) |
| Platalea ajaja | RP | 23.1 | (91) | 66.0 | (100) | 30.2 | (83) |
| FILTRADOR | |||||||
| Phoenicopterus ruber | RB | 100 | (100) | 100 | (100) | 100 | (100) |
Los gremios de mayor densidad en ambos años (no. ind./km2) fueron los buceadores, herbívoros y filtrador (Cuadro 1). Los valores de densidad para buceadores y herbívoros fueron los más altos del estudio, pero sólo durante la migración otoñal. En el caso de filtrador, los valores siempre se mantuvieron cercanos a los 50 ind./km2. De forma general, los valores de densidad más bajos por gremio se registraron durante la residencia de verano. Los gremios sondeadores profundos y omnívoros presentaron los más bajos valores de densidad (<10 ind./km2) y el patrón se mantuvo en los tres periodos.
Al comparar la representatividad de cada gremio entre años, no se encontraron evidencias de que las variaciones detectadas no fueran al azar en ninguno de los tres periodos (migración primaveral: diferencia real = -8.9, P = 0.527; residencia de verano: diferencia real = 0, P = 0.475; migración otoñal: diferencia real = 1.04, P = 0.495). Por tal motivo las comparaciones entre periodos fueron hechas sin tener en cuenta el año. Este análisis temporal registró diferencias para sondeadores someros, buscadores aéreos, buceadores, herbívoros y filtrador (Figura 1). En el caso de los dos primeros gremios las diferencias se encontraron entre el periodo de migración primaveral y los dos restantes. En cambio para filtrador y buceadores estuvo entre el periodo de residencia de verano y el resto; y para herbívoros entre el periodo de migración otoñal y los dos restantes. El análisis de la abundancia proporcional por gremio ayudó a visualizar cómo dentro del periodo de migración primaveral existe una mayor representación de todos los gremios, en tanto, en los otros periodos siempre hay un gremio con una contribución mayor de 50%. Este es el caso de herbívoros en migración otoñal y filtrador en residencia de verano.
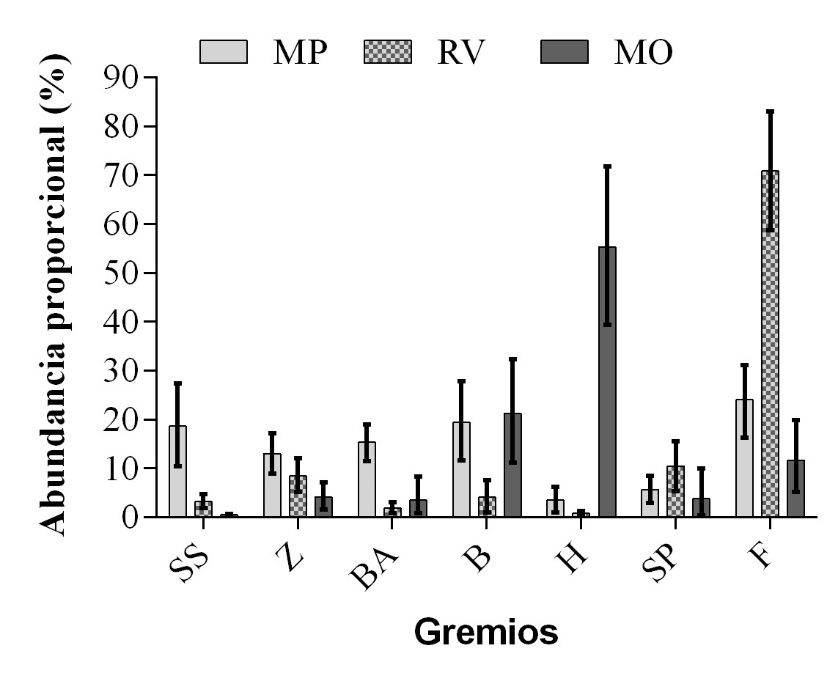
Figura 1 Valores de abundancia proporcional de cada gremio por periodos (MP: migración primaveral, RV: residencia de verano, MO: migración otoñal). Estos valores están representados por las barras. Por su parte, los bigotes representan el intervalo de confianza al 95% calculado mediante Monte Carlo. En la figura está excluido el gremio omnívoro por su baja contribución en los tres periodos.
El análisis de la abundancia proporcional de las especies por periodos mostró variaciones por gremios. Existieron gremios como sondeadores someros y zancudas donde ninguna de las especies que lo integraron mostraron valores de abundancia proporcional por encima de 60% (Cuadro 2). Sin embargo, en el resto de los gremios identificamos una especie con un valor superior al 60% en al menos dos de los periodos muestreados. Las excepciones a esto fue filtrador que como se mencionó anteriormente estuvo compuesto por una sola especie, y omnívoros el cual se detectó en dos de los periodos conformado de igual modo por una especie.
Aunque los valores de abundancia proporcional para las especies del gremio sondeadores someros estuvieron por debajo del 60%, hubo especies que se detectaron en más de la mitad de los conteos hechos por periodo. Entre estas especies se encontraron: Pluvialis squatarola (chorlo gris), Tringa melanoleuca (patamarilla mayor), Tringa flavipes (patamarilla menor), Tringa semipalmata (playero pihuiuí), Arenaria interpres (vuelvepiedras rojizo), Himantopus mexicanus (monjita americana) y Limnodromus griseus (costurero pico corto). De ellas, la única que mantuvo altos valores de frecuencia en los tres periodos estudiados fue L. griseus, el resto por lo general lo presentó durante la migración primaveral. Por su parte, las especies incluidas dentro del gremio zancudas también presentaron altos valores de frecuencia de observación. A diferencia de sondeadores someros, este comportamiento fue observado en la mayoría de las especies que integraron el gremio y se mantuvo en los tres periodos estudiados (Cuadro 2). Es de resaltar que en varias ocasiones estos valores llegaron a 100%. Dentro de los valores de abundancia proporcional para las especies de ambos gremios podemos resaltar los encontrados para Ardea alba (garza blanca) durante la migración otoñal, para H. mexicanus en residencia de verano; y L. griseus y Egretta rufescens (garza rojiza) en migración primaveral.
En los gremios buscadores aéreos, sondeadores profundos, buceadores y herbívoros las especies que se registraron con una abundancia proporcional mayor al 60% tanto en migración primaveral como otoñal fueron: Hydroprogne caspia (charrán del caspio), Eudocimus albus (ibis blanco), Fulica americana (gallareta americana) y Spatula discors (cerceta alas azules), respectivamente (Cuadro 2). En el caso de las dos últimas especies los valores llegaron a estar por encima del 90%. Durante la residencia de verano sobresalieron Pelecanus erythrorhynchos (pelícano blanco americano) y Sternula antillarum (charrán mínimo) dentro del gremio buscadores aéreos; Platalea ajaja (espátula rosada) dentro de sondeadores profundos y Dendrocygna arborea (yaguaza) dentro de herbívoros. En este periodo el gremio buceadores estuvo conformado únicamente por Phalacrocorax auritus (cormorán orejón). De forma general, todas las especies mencionadas hasta el momento también se registraron con altos valores de frecuencia (>50%). Sin embargo, hubo otras especies que también resaltaron por los altos valores de frecuencia con que se registraron (≥ 70%) y no por sus valores de abundancia proporcional, por ejemplo P. auritus en los tres periodos de estudio; P. erythorynchos, P. occidentalis (pelícano café), Thalasseus maximus (charrán real), Gelochelidon nilotica (charrán pico grueso) en migración otoñal; y Megaceryle alcyon (martín pescador norteño) tanto en migración otoñal como primaveral (Cuadro 2).
Discusión
La identificación de ocho gremios dentro del área de estudio puede considerarse como un indicador de la amplia diversidad de microhábitats y recursos que las aves acuáticas pueden encontrar dentro del sistema lagunar de Las Salinas. Similar número de gremios han sido identificados en otros humedales cubanos naturales (Acosta et al. 1992, Lamarté et al. 2018), seminaturales (González y Jiménez 2011) y antrópicos (Acosta et al. 2002), por lo que la estructura de gremios encontrada es representativa de los humedales cubanos. Es por ello, que más que la cantidad de gremios representados, un análisis más profundo incluyó para cada gremio el cálculo de su riqueza específica, densidad y abundancia proporcional.
Los sondeadores someros, las zancudas y los buscadores aéreos los identificamos como los gremios con mayor número de especies dentro del ensamble. Resultados similares fueron registrados por Lamarté et al. (2018) en la ensenada del Jato, perteneciente al Área de Recursos Manejados Cayo Sabinal. Los gremios sondeadores someros y buscadores aéreos están representados por un elevado número de especies dentro del archipiélago cubano (Llanes et al. 2002). De las 38 especies que conforman el gremio de los sondeadores someros en Cuba, se registró en Las Salinas el 39%; sin embargo, de las 12 que conforman el gremio de las Zancudas se observó el 83%. Aunque los análisis basados meramente en la riqueza específica pueden estar sesgados, esto podría considerarse una primera evidencia de que el área de Las Salinas es un hábitat de mayor importancia para el gremio de las zancudas, donde a su vez la mayoría de las especies que lo integró mostró una alta frecuencia de aparición en los tres periodos de estudio, y no tanto para el de los sondeadores someros. Como mencionamos en la descripción del sitio, el sistema lagunar de Las Salinas presenta variaciones en sus niveles de agua, los valores más bajos se presentan durante la migración primaveral. Los sondeadores someros son especies pequeñas, el 42% de las especies descritas para Cuba son menores de 22 cm con tarsos que van desde 20.1 mm (Calidris minutilla,Jiménez et al. 2015) a 26.8 mm (Calidris alpinaKoloski et al. 2016) por lo cual hacen un mayor uso de este humedal costero cuando bajan los niveles de profundidad de agua. Los elevados valores de frecuencia registrados para varias especies del gremio durante la migración primaveral refuerzan este planteamiento.
Aun cuando los gremios variaron en la cantidad de especies por categoría de permanencia, los ocho gremios estuvieron representados en los tres periodos analizados (migración primaveral, residencia de verano y migración otoñal). Los gremios donde predominaron especies residentes invernales o bimodales mostraron los valores más bajos de densidad durante el periodo de residencia de verano (en comparación con los otros periodos). Las especies migratorias comienzan su viaje de regreso a las zonas de reproducción a partir del mes de marzo, por lo que era de esperar que el valor de densidad de los gremios en el periodo de mayo-junio fuera bajo. En los restantes periodos, correspondientes a la etapa migratoria, los valores aumentan por la llegada de las aves desde sus sitios de reproducción durante la migración otoñal; o por la incorporación de aves desde regiones más al sur que utilizan a nuestra isla como sitio de parada durante la migración primaveral. Estudios recientes dentro de Las Salinas han registrado los cambios en la dinámica del ensamble durante el periodo migratorio (González et al. 2016) y esta tendencia se mantiene de igual modo en la dinámica de los gremios.
Registramos a los gremios buceadores, herbívoros y filtrador como los de mayor densidad. En el caso de los dos primeros gremios, los valores fueron particularmente altos durante el periodo de migración otoñal y estuvieron dados por la abundancia de F. americana y S. discors, las cuales representaron más del 90% del gremio. Ambas especies al igual que P. ruber son especies gregarias categorizadas como muy comunes en los humedales cubanos (Garrido y Kirkconnell 2011), y en particular dentro de Las Salinas se concentran en grandes números tanto para forrajear como para descansar. Hasta el momento no se han observado parejas de P. ruber criando dentro del área, sin embargo, el hecho de que se hayan registrado individuos juveniles anillados procedentes de la península de Yucatán, México, y del humedal de Río Máximo, evidencia que Las Salinas tiene relevancia nacional y regional como un espacio en la conservación de la especie.
Varias de las especies incluidas en el gremio sondeadores someros también se caracterizan por su conducta gregaria durante el forrajeo y descanso (Acosta et al. 1992, Acosta et al. 2002, Lamarté et al. 2018). Sin embargo, este gremio no se encontró entre los de mayor densidad. Es posible que las variaciones en los niveles de agua limiten la cantidad de microhábitats disponibles para las especies que conformaron este gremio. Durante el estudio, los sondeadores someros se distribuyeron en pequeños bandos a lo largo del sistema lagunar y se concentraron en sitios con presencia de afloramientos rocosos o en los bordes del sistema lagunar donde la profundidad de agua es menor. Por otro lado, la mayoría de las especies que se incluyen en el gremio sondeadores someros son especies migratorias por lo que las mayores densidades las registramos en los periodos de migración, fundamentalmente en la migración primaveral. Los meses incluidos en el periodo de migración primaveral (febrero, marzo) pertenecen a la estación de seca y son los meses donde los niveles de agua están más bajos. Al igual que sondeadores someros, para buscadores aéreos y zancudas el periodo de mayor importancia según los valores de densidad fue la migración primaveral. Al disminuir los niveles de agua, no sólo aumenta la accesibilidad a los hábitats, sino que determinados recursos como los peces, tienden a concentrarse y ser más accesibles.
El análisis entre años de la abundancia proporcional por gremios no reflejó variaciones significativas. Esto podríamos considerarlo como una evidencia de la estabilidad interanual en la estructura en gremios del ensamble, lo que se traduce a su vez en una constancia en la disponibilidad de hábitats de forrajeo y recursos tróficos que son utilizados por los representantes de cada gremio. Por su parte, las variaciones encontradas entre periodos para determinados gremios, refuerzan los resultados que hemos obtenido de forma aislada en los análisis de riqueza, densidad y frecuencia. El sistema lagunar de Las Salinas es particularmente un hábitat importante para sondeadores someros y buscadores aéreos durante la migración primaveral, en tanto para herbívoros los es durante la migración otoñal. No obstante, para otros gremios como zancudas el hábitat de Las Salinas parece ser importante durante todo el año. Para este último gremio, los resultados indicaron que las especies de mayor contribución cambian por periodos, sin embargo, de forma general existe un mecanismo de compensación en su estructura que impidió que registráramos variaciones significativas entre periodos. La identificación de periodos de mayor importancia para cada gremio dentro de Las Salinas podría ser de gran aplicación. El establecimiento de acciones de manejo o programas de monitoreo para alguna de las especies que conforman estos gremios podrían estar restringidas a estos periodos.
El presente estudio, aunque incluyó la dinámica de los gremios de aves acuáticas presente en Las Salinas durante dos años, aún se considera un estudio a corto plazo. Sin embargo, consideramos que los resultados presentados establecen una línea base que puede ser utilizada, no sólo para el manejo y la conservación de las aves que utilizan este importante humedal costero, sino como un punto de partida en la detección de futuros cambios ambientales. Las aves representativas de cada gremio tienen diferentes límites de tolerancia, preferencias ambientales, necesidades tróficas, y requerimientos energéticos; por tanto, su presencia/ausencia o densidad podría ser un importante indicador del hábitat (Newman 1979).

