











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkEditora asociada: Iriana Zuria Jordan
Introducción
El color del plumaje juega un papel importante en la comunicación, camuflaje y procesos fisiológicos en las 9 993 especies de aves modernas (Neornithes, Thomas et al. 2014). En la actualidad, se describen diversos tipos de anormalidades pigmentarias con base en el exceso, disminución, defecto o ausencia de pigmentos, principalmente melaninas (Grouw 2006, Davis 2007). Sin embargo, la terminología aplicada por más de un siglo ha sido confusa e inconsistente (Coues 1868). En consecuencia, no existe un consenso estricto referente a las definiciones presentadas con respecto a estas anormalidades (C. Benesh com. pers. <www.birdforum.net/blog.php?b=79>) debido a que estos conceptos, en un principio, no fueron elaborados para aves, sino generados para aplicarse a la patología humana y otros mamíferos (Buckley 1982). En la última década, se han desarrollado esfuerzos por resolver estas inconsistencias mediante la generación de información específica para las anormalidades que afectan la coloración del plumaje de las aves con base en descripciones detalladas sobre sus causas, métodos filogenéticos y reportes de especies afectadas (Grouw 2012, 2013, 2014).
La mayoría de los colores pigmentarios se producen por medio de la participación de tres tipos de moléculas: melanina (eumelanina y phaeomelanina), caroteno y porfirina. Las melaninas definen los colores café oscuro, gris y negro por medio del subtipo molecular eumelanina; mientras que los colores café claro a rojizo y algunos amarillos son definidos por medio del subtipo molecular phaeomelanina (McGraw et al. 2003, McGraw y Nogare 2004, Hill, McGraw 2006, Davis 2007). La melanina es un pigmento producido en la piel, escamas, plumas, pelo y ojos a través de una serie de reacciones bioquímicas complejas en la que la oxidación del aminoácido tirosina (mediante la enzima tirosinasa) da lugar a la formación de la melanina (Oetting et al. 2003, Fertl y Rosel 2009).
Por su parte, los carotenoides son colorantes químicos producidos por las plantas y obtenidos por las aves a través de la dieta, transformados vía enzimática para influir en la expresión de colores amarillo, naranja y rojo intenso, y que al combinarse con la melanina pueden producir el color verde olivo (McGraw et al. 2003, McGraw y Nogare 2004, Hill, McGraw 2006, Davis 2007).
Las porfirinas son producidas por modificación de aminoácidos y se distinguen por tener en su estructura átomos de nitrógeno. Este tipo de pigmentos produce el color rojo, café marrón y verde, los cuales se observan ante la iluminación de la luz ultravioleta. Los colores azul, violeta y verde son estructurales y se producen por la interferencia o difracción y por el ángulo de incidencia de la luz blanca en el cuerpo del ave (similar a un prisma), como resultado de la combinación de las capas de aire y la queratina dentro de la pluma, produce la coloración metálica o iridiscente en colores azul, verde o violeta (Gill 1990).
La expresión fenotípica de anormalidades pigmentarias en la coloración del plumaje de las aves resulta del exceso, ausencia o deficiencia de melaninas, carotenos y porfirinas (Griffiths et al. 2000, Davis 2007). Dichas expresiones fenotípicas pueden ser causadas por anomalías en la estructura química del metabolismo (durante la elaboración de plumas), así como por factores físicos ambientales meramente externos a través de suciedad, cambios en la exposición a radiación solar o abrasión, que suceden principalmente de forma accidental (Davis 2007). Las anormalidades pigmentarias se le han atribuido a la endogamia, hibridación, deficiencia nutrimental, falta de exposición a la luz solar y contaminación ambiental (Bensch et al. 2000, Møller y Mousseau 2001).
Si consideramos que diversas anomalías pueden producir efectos similares en un individuo (Grouw 2013, 2014), resultaría difícil lograr determinarlas en campo por indefinición en terminología, falta de unificación de criterios o falta del interés por generar información concerniente; a esto también se suma la variabilidad en los patrones que rigen el origen y los mecanismos que provocan esas anormalidades en las aves. Estos aspectos limitan la determinación acertada en campo (Davis 2007), errores que en la identificación de las anomalías pigmentarias han ocurrido por más de un siglo (Gutiérrez 1994), ya que los términos leucismo y albinismo se habían generalizado para nombrar diversos tipos de anormalidades pigmentarias en aves (Grouw 2012, 2013).
Si tratamos de facilitar la identificación de anormalidades pigmantarias con base en una clasificación esquemática, con definiciones explícitas y unificando la terminología sobre su co nocimiento en aves, así como la ubicación fenotípica de los pigmentos en los individuos, podremos determinar correctamente el tipo de anormalidad y comprender su ocurrencia a nivel poblacional. Así, con mayor información, se puede inferir sobre los patrones espaciotemporales y las causas que las provocan.
Con base en lo anterior, el objetivo principal de este trabajo fue desarrollar una metodología que permita determinar las anormalidades pigmentarias de las aves por medio del uso de claves dicotómicas, con el fin de alcanzar una mayor precisión y menor confusión en su identificación. Un siguiente objetivo fue generar e implementamos un sistema de claves dicotómicas que condensan la información especializada que unifique y facilite la identificación de estas anormalidades. Finalmente, se buscó documentar la presencia de aves con problemas de pigmentación en el plumaje, lo cual nos permite evaluar las claves dicotómicas aquí generadas. Se entiende que la identificación puede tornarse difícil si sólo se toman en cuenta imágenes fotográficas o registros visuales, por lo que las claves dicotómicas facilitan la identificación de la anormalidad durante el manejo directo del individuo (captura o colección científica). Para lograr una mejor apreciación de las anormalidades pigmentarias a la hora de identificarlas, sugerimos tomar en cuenta aspectos de tipo ambiental como la luminosidad y temperatura, biología del ave, desgaste del plumaje, la temperatura corporal y la edad del individuo, datos que permiten asegurar una identificación correcta del tipo de anormalidad.
Método
Elaboramos un esquema sinóptico de llave que representa una estructura organizada y coherente de la temática y sus múltiples relaciones, lo que es útil para detectar semejanzas o diferencias entre variables fenotípicas (Figura 1). Posteriormente, diseñamos un cuadro comparativo con una lista de las anormalidades pigmentarias, sus respectivas sinonimias y los efectos relacionados con la pigmentación (Cuadro 1). Finalmente, la información que generamos se integró en una clave dicotómica (Cuadro 2) y se relacionó con base en las características diagnósticas de algunos casos documentados en la tortolita mexicana (Columbina inca), paloma turca (Streptopelia decaocto), correcaminos norteño (Geococcyx californianus), pijuy (Crotophaga sulcirostris) y tordo de ojo rojo (Molothrus aeneus). Estos casos de anormalidades los obtuvimos a partir de observaciones ocasionales en áreas con impacto antropogénico en Tamaulipas, México, entre 2012 y 2016.
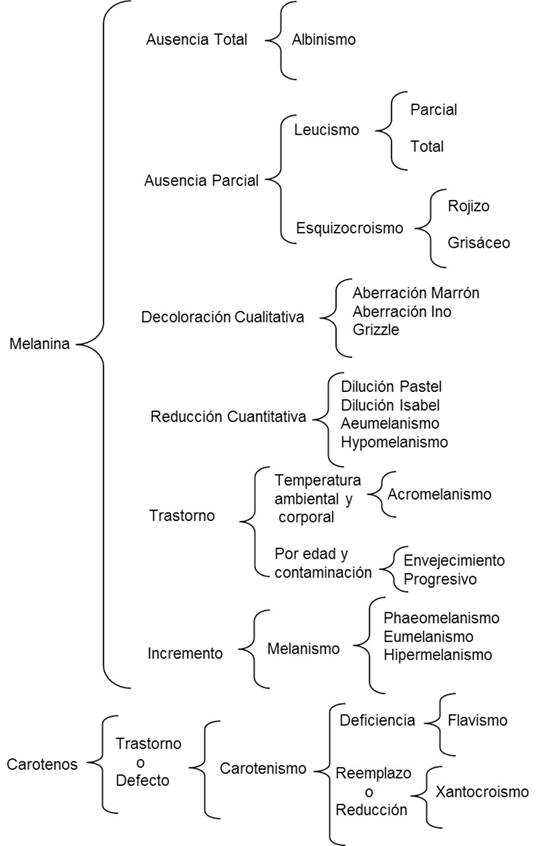
Figura 1 Esquema sinóptico de llave de anormalidades pigmentarias en aves, con énfasis en pigmentos de melanina y caroteno.
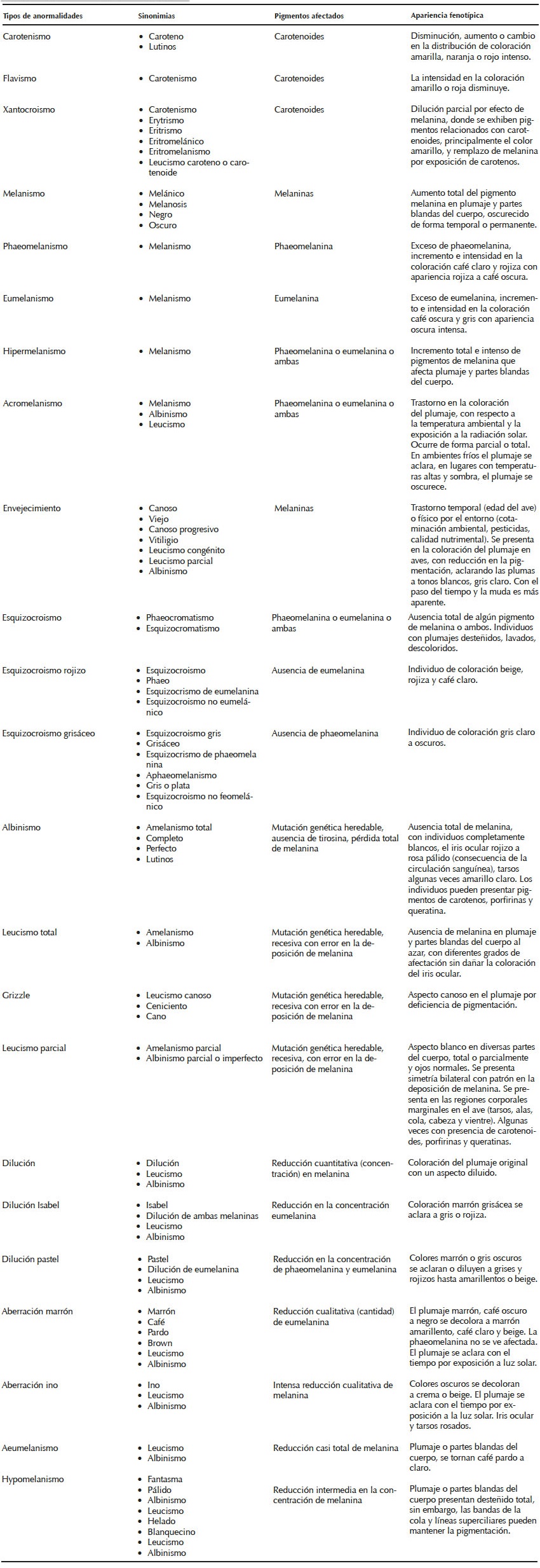
Cuadro 1 Cuadro comparativo de los tipos de anormalidades en la pigmentación corporal y del plumaje en aves presentadas en este trabajo, y las sinonimias empleadas con base en la terminología propuesta por Pettingill (1956), Harrison (1964), Buckley (1982), Grouw (1997, 2000a, 2000b, 2006, 2010, 2011, 2012, 2013, 2014), Grouw y Nolazco (2012), Grouw y Blochb (2015) y Davis (2007). Se hace referencia en el texto a los pigmentos afectados y los cambios resultantes en la apariencia fenotípica del ave.

A lo largo de la clave dicotómica se describen los pigmentos afectados y sus cambios expresados en el fenotipo de las aves, en cuanto a coloración del plumaje, pico y partes blandas del cuerpo (tarsos, piel e iris ocular), con base en la terminología propuesta por Pettingill (1956), Harrison (1964), Buckley (1982), Grouw (1997, 2000a, 2000b, 2006, 2010, 2011, 2012, 2013, 2014), Grouw y Nolazco (2012), Grouw y Blochb (2015) y Davis (2007). Lo anterior nos permitió la identificación y comparación de los distintos tipos de anormalidades pigmentarias en relación con caracteres excluyentes (Vilches et al. 2012).
Los caracteres que utilizamos para las opciones de la clave dicotómica tienden a ser constantes, de tal manera que las anormalidades pigmentarias puedan ser identificadas sin dificultad, comenzando con características generales muy contrastantes (p. ej., con o sin presencia de melanina). Para la estructuración de la clave dicotómica, asignamos a cada una de las opciones un símbolo (número y letra) que las identifique, y que permita el flujo de la información hasta lograr la determinación (Vilches et al. 2012). Para relacionar los aspectos morfológicos externos del plumaje y las partes blandas del cuerpo (presencia o ausencia total o parcial, disminución o aumento y con o sin distribución de simetría bilateral en la expresión de melaninas, así como los defectos de la expresión de carotenos). Por lo cual, es necesario observar detalladamente las características corporales de las aves, de forma cualitativa y cuantitativa, para generar la opción múltiple en cada articulación de la clave.
Estudios de caso
Tortolita mexicana (Columbina inca)
El 27 de abril de 2015, en Ciudad Mante (balneario El Campestre), Tamaulipas (22°42’42.22”N, 98°58’40.60”O; 86 msnm), registramos dos individuos adultos de Columbina inca en un huerto de mango (Mangifera indica), forrajeando a nivel de suelo. Ambos individuos presentaban un incremento notable de melanina que oscurecía su plumaje, por lo que el patrón escamoso característico de la especie apenas era distinguible (Figura 2a). La frecuencia de reportes sobre casos de anormalidades pigmentarias en Columbiformes ha sido común (Braun y Boyd 1979, Yusti-Muñoz y Velandia-Perilla 2013). Existe conocimiento sobre melanismo para la especie en otras regiones del mundo <www.internationaldovesociety.com/SeedSpecies/IncaDove.htm>; sin embargo, parece ser el primer caso reportado de melanismo en México (Rodríguez-Ruíz et al. 2015).

Figura 2 a) Dos individuos de tórtola (Columbina inca) con melanismo, incremento de coloración en el plumaje; b) Paloma turca (Streptopelia decaocto) con aberración ino en alas; c) Paloma turca (Streptopelia decaocto) con aberración ino en alas con diferente grado de decoloración; d) Correcaminos norteño (Geococcyx californianus) con aberracion ino con todo el plumaje decolorado; e) Tordo de ojo rojo (Molothrus aeneus) con leucismo parcial en plumas rectrices.
Paloma turca (Streptopelia decaocto)
El 28 de marzo de 2015, a las 14:09 h, en el Ejido El Olmo (balneario El Salto del Tigre), municipio de Güemez, Tamaulipas (24°0’52.26”N, 99°19’09.77”O; 283 msnm), registramos un individuo adulto de Streptopelia decaocto (Figura 2b) que presentaba un ala y el dorso sin melanina en el plumaje, mientras que el resto mostraba la coloración típica de la especie. Esto lo identificamos como caso de aberración ino, una mutación común en la especie (H. van Grouw com. pers.) debido a una sensibilidad alta a luz solar, que aclara el plumaje en un periodo corto. El ave fue detectada posando en un árbol de framboyán (Delonix regia) y después sobre un cable de energía eléctrica, en compañía de otro individuo de la misma especie. El 19 de junio de 2016, frente al Instituto Tecnológico de Ciudad Victoria, Tamaulipas (23°45’08.29”N, 99°09’53.92”O; 327 msnm), detectamos la presencia de un individuo adulto de S. decaocto con decoloración gradual en el cuerpo, pero principalmente en alas, lo que coincide con la aberracion ino (Figura 2c). La incidencia de casos sobre anormalidades pigmentarias para la especie no parece ser un hecho aislado ni en México ni a nivel mundial. Situaciones similares han sido asociadas a la calidad del hábitat, contaminación ambiental, dieta y baja variabilidad a nivel genético en poblaciones reducidas y semi-aisladas (Bensch et al. 2000) en áreas urbanas pequeñas, suburbanas o agrícolas (Contreras y Ruíz 2011, Ayala-Pérez et al. 2015). Una propuesta es realizar estudios biogeográficos que ayudarían a identificar las poblaciones expuestas a estrés ambiental o endogamia.
Correcaminos norteño (Geococcyx californianus)
El 20 de febrero y 1 de mayo de 2015, en el ejido El Ébano, municipio de Llera, Tamaulipas (23°32’57.09”N, 99°02’25.84”O; 471 msnm), avistamos un individuo de Geococcyx californianus con el plumaje y partes blandas del cuerpo completamente blancos e iris ocular y tarsos ligeramente rosados (Figura 2d); por lo que la clasificamos como aberración ino. Descartamos la posibilidad de un albinismo por ser una condición extrema y muy rara en la naturaleza, al ser una mutación incapaz de producir melanina. Además, rara vez se observan aves albinas gregarias adultas debido a que los polluelos mueren poco después de abandonar el nido por su mala visión que interfiere en la búsqueda de alimento y sitios de descanso, y por su visible exposición a depredadores (Grouw 2012, 2013). El ave se observó por vez primera junto a dos individuos, y en una segunda ocasión acompañado de tres individuos (Figura 2e). Este reporte representa el segundo para la especie, pues el primero fue documentado en Hurricane, Utah, EUA, en 2007, por Harlan Daubs (determinado como albino <www.utahbirds.org/featarts/2008/AlbinoBirdPhotos.htm>). Coincidentemente ambos casos se encuentran en áreas marginales de su distribución geográfica, lo que podría ser un factor que influye en la anormalidad de su pigmentación (Krecsák 2008). Sin embargo, las implicaciones ecológicas y los factores que causan los patrones de anormalidades pigmentarias en aves aún no son bien conocidas.
Pijuy (Crotophaga sulcirostris)
El 1 de mayo de 2015, en el poblado Loma Alta, municipio de Gómez Farías, Tamaulipas (22°54’19.92”N, 99°01’37.18”O; 75 msnm), avistamos un individuo de Crotophaga sulcirostris con plumaje blanco iridiscente (con presencia de queratina) y tres motas negras pequeñas en la zona dorsal, pecho y abdomen con simetría bilateral en el cuerpo del ave, mientras que el pico, ojos y patas presentaban el color oscuro típico de la especie. El individuo se encontraba en un cultivo de caña de azúcar (Saccharum officinarum) y se desplazó hacia las ramas de un huizache (Acacia farnesiana), en compañía de tres individuos con coloración de plumaje normal. Adicionalmente, el 12 de mayo de 2015, observamos al individuo con plumaje aberrante en el mismo sitio con tres individuos posados en un arbusto de guazima (Guazuma ulmifolia). En ambos casos no fue posible obtener una imagen fotográfica como evidencia. Existen registros de leucismo parcial para la especie en Guayas, Ecuador, acompañados de fotografías (Cadena-Ortiz et al. 2015).
Tordo de ojo rojo (Molothrus aeneus)
Con base en los registros de observaciones ocasionales obtenidos durante un monitoreo que desarrollamos en las épocas de inverno y primavera (noviembre a abril) de 2012 a 2016, en el Parque Bicentenario de Ciudad Victoria, Tamaulipas (23°45’03.96”N, 99°06’02.07”O; 263 msnm), registramos la presencia de un individuo macho adulto de Molothrus aeneus con dos plumas rectrices centrales sin melanina, es decir, completamente blancas (Figura 2e). El resto del plumaje presentaba la coloración típica de la especie, al igual que las partes blandas del cuerpo (piel, ojos y tarsos) y el pico. Lo identificamos como un caso de leucismo parcial; excluimos el envejecimiento, ya que en este último se magnifica el aclaramiento de las plumas, mientras que el leucismo permanece con el paso del tiempo. Esto hace evidente la relevancia de monitorear al individuo para observar si existe un incremento en la aparición de plumaje blanco (Grouw 2011). Otra de las características excluyentes es que cuando sucede el leucismo, aunque es poco común, se muestra de forma simétrica (condición de simetría bilateral); comúnmente se presenta en las áreas marginales de plumas rectrices, cabeza, alas, vientre y tarsos (Grouw 2013). Otra causa de pérdida de melanina en las plumas rectrices puede deberse a un accidente o lesión por ataque de algún depredador o que en la nueva muda no se haya presentado pigmentación (H. van Grouw com. pers.). Sin embargo, este individuo fue observado durante cuatro años consecutivos en la misma zona durante la época migratoria en parvadas mixtas con el tordo cabeza café (M. ater) y el zanate mexicano (Quiscalus mexicanus). Las parvadas estaban constituidas, aproximadamente, por 1 250 individuos, de los cuales se cuantificaron 450 M. aeneus, 600 M. ater y 33 Q. mexicanus. Lo anterior nos indicaría que, la incidencia de esta anormalidad, fue inferior a 1%. Cabe destacar que la permanencia del individuo en el grupo resulta importante, pues a pesar de su condición no fue excluido y llegó a la etapa adulta, lo que concuerda con lo que recientemente se reportó para M. ater en un área urbana del noroeste de México (Hernández et al. 2016).
Discusión
A nivel mundial se han documentado casos sobre anormalidades pigmentarias en diversos grupos taxonómicos por exceso o disminución de pigmentos, principalmente melaninas (Grown 2006, Davis 2007). Actualmente, se conocen más de 100 mutaciones en el gen de la tirosinasa, el cual ha sido asociado con albinismo en aves, mamíferos, reptiles, anfibios y peces (Fertl y Rosel 2009, Oetting et al. 2003). Además, tiende a observarse un interés creciente por el desarrollo de estudios y conocimiento de anormalidades pigmentarias en diversos grupos de vertebrados: aves (Gross 1965, Nemésio 2001, Piacentini 2001, Urcola 2011, Cadena-Ortiz et al. 2015), mamíferos (Arriaga-Flores et al. 2016), reptiles (Krecsák 2008), anfibios (Galán 2010) y peces (Veena et al. 2011). En diversos estudios sobre anormalidades pigmentarias se han reconocido alrededor de 572 especies de vertebrados a nivel mundial con alguna de estas aberraciones; las aves ocupan la mayoría de estos casos (más de 350 especies, Hayley-McCardle 2012). Con respecto a estudios de anormalidades pigmentarias de aves en México, únicamente se ha documentado la existencia de leucismo parcial o total y albinismo (Tinajero y Rodríguez-Estrella 2010, Carbó-Ramírez et al. 2011, Ayala-Pérez et al. 2015, González-Arrieta y Zuria 2015).
Vale la pena reflexionar acerca de la problemática abordada. ¿La información científica es suficiente o faltan científicos para abordar el tema? ¿La terminología empleada para consignar las anormalidades pigmentarias es apropiada? O bien, ¿la determinación de la anormalidad pigmentaria ha sido la adecuada? Un caso donde se manejaron errores de identificación ocurrió con el albinismo de Q. mexicanus y el leucismo de C. inca (Rodríguez-Ruíz et al. 2015), los cuales se debieron clasificar como aberración ino o envejecimiento por presentar plumas envejecidas o descoloridas por luz solar, con aberración marrón en otras plumas (H. van Grouw com. pers.). Por lo cual, pensamos que la identificación de alguna anormalidad pigmentaria debería ser abordada con mayor rigor y mediante el uso de claves dicotómicas.
Las claves dicotómicas propuestas y la forma de emplearlas en anormalidades pigmentarias pretenden contribuir a la unificación de criterios terminológicos, que han sido difusos por más de un siglo (Coues 1868, Davis 2007). Además, tiende a facilitar la determinación y brindar certeza, con lo que se podría aumentar el número de reportes de anormalidades pigmentarias en la literatura especializada y abonar al modelaje de patrones de incidencia, cambios en la concentración y distribución de los pigmentos, entre otros temas importantes. También, resulta conveniente registrar las consecuencias, costos ecológicos y genéticos de las poblaciones e individuos que lo padecen, mapeo de sitios y la relación que guardan con ciertas variables ambientales, a fin de dar seguimiento a los individuos con anormalidades pigmentarias en lo que corresponde a supervivencia, comportamiento (Bensch et al. 2000) y aceptación colectiva dentro de su especie. Es indispensable que los estudios realizados presenten una descripción detallada de las anormalidades pigmentarias, debido a que pueden presentarse confusiones (Grouw 2006, 2010, 2011).
Se ha demostrado que la pérdida de pigmentación conlleva a modificar los hábitos de los individuos afectados (Caro 2005, Rodríguez-Ruíz et al. 2014). Ejemplo de ello es la desventaja de supervivencia por la susceptibilidad mayor a depredadores naturales, exclusión social intraespecífica y reducción en la atracción reproductiva, lo que implicaría menor posibilidad de sobrevivir (Caro 2005, Espinal et al. 2011).
La reducción en la expectativa de vida de los individuos leucísticos sería más sensible en especies gregarias (Nogueira y Alves 2011). Sin embargo, en nuestro caso, los individuos con anormalidades pigmentarias, al parecer, no son segregados por la población de la que forman parte, sino que son aceptados; tal como ha sido observado en numerosas especies de vertebrados, quienes logran reproducirse y seguir su ciclo de vida normal (Atauchi 2015). Documentar este tipo de casos puede contribuir a evaluar la frecuencia con la que ocurren dichas anormalidades pigmentarias y el grado de variación entre poblaciones de aves (Ayala-Pérez et al. 2014, 2015). También se cuenta con información de los efectos negativos que ocurren en las relaciones intraespecíficas de individuos con anormalidad pigmentaria (Beier y Noos 1998, Fahrig 2003, Laiolo y Arroyo-Solís 2011), donde las anormalidades frecuentes en el plumaje de aves podría ser indicativo de endogamia en poblaciones pequeñas y aisladas, propiciadas por estrés ambiental (Bensch et al. 2000).
La frecuencia en la detección de anormalidades pigmentarias en aves asociadas a centros urbanos podría depender de mecanismos interactuantes a nivel poblacional, en particular el tamaño reducido del núcleo familiar (Arriaga-Flores et al. 2016). Sin embargo, estas poblaciones suelen tener una variabilidad genética reducida entre individuos de distintas poblaciones (Martínez-Sánchez et al. 2001), lo que conlleva a mayores probabilidades de manifestar anormalidades pigmentarias.
Los registros de especies ecológicamente tolerantes a la presencia humana en áreas urbanas también sugieren que los individuos con anormalidades pigmentarias podrían ser fácilmente detectados, mayor aún si desarrollan actividad diurna o si emiten llamados o vocalizaciones constantes. Del mismo modo, su presencia en estas áreas podría deberse a una menor tasa de depredación o mayor disponibilidad de recursos (Cadena-Ortiz et al. 2015). En ocasiones, las plumas blancas también pueden ser el resultado de una lesión o una mala nutrición, con la probabilidad de que en la siguiente muda se recupere el patrón de coloración normal en el cuerpo del ave. Sin embargo, existe otra anormalidad pigmentaria nombrada envejecimiento, donde el ave tiende a perder la coloración del plumaje con respecto a la edad, cuando se trata de casos tempranos se puede discernir del leucismo, pero en edades avanzadas es casi imposible diferenciarla en campo (Grouw 2012, 2013). Algo semejante podemos encontrar en colecciones científicas producto de la manipulación frecuente o el factor tiempo que provoca el desgaste y en consecuencia la pérdida del patrón de pigmentación original en el ejemplar de estudio.
Los reportes de albinismo son escasos entre aves (<1%), lo cual podría deberse a la baja tasa de supervivencia que presentan los individuos con esta condición en vida libre (Grouw 2006). Las causas pueden ser atribuidas a diversos factores: fotofobia (intolerancia a la luz, que conlleva a afecciones irritantes del globo ocular), alta perceptibilidad ante los depredadores, o dificultades en la termorregulación (helioterma) ya que, debido a la ausencia total de melanina, estas aves tienen una baja capacidad de absorción de la radiación solar, lo cual implica una seria desventaja con el resto del grupo, ya que la probabilidad de supervivencia se ve drásticamente afectada, por lo que disminuye la posibilidad de encontrar individuos adultos (Rivera et al. 2001).
Mientras la presencia de individuos melánicos es común en condiciones asociadas con el aislamiento, ya sea en las islas oceánicas o en picos de montaña (regiones alpinas res tringidas), lo que se atribuye a procesos ecológicos a corto plazo asociados a competencia interespecífica (Wilson 1961). En aves, el melanismo ocurre cuando las poblaciones se encuentran apartadas geográficamente y concentradas con altas densidades dentro de tierras continentales y, por lo tanto, la probabilidad sobre la frecuencia de individuos melánicos es mayor (Ferrer-Sánchez y Rodríguez-Estrella 2014).
El melanismo puede presentar propiedades térmicas y camuflaje en regiones insulares donde existen superficies oscuras. Las causas pueden derivar por alta nubosidad en las regiones costeras, así como protección contra la refracción de luz solar proveniente del suelo arenoso y salino (Kettlewell 1973), por lo que el melanismo bajo estas condiciones se ha clasificado de forma monomórfica en algunas poblaciones (Pérez-Mellado 1984). Por otro lado, para el caso de especies continentales el melanismo es considerado como una anormalidad pigmentaria rara y restringida (Galán et al. 2011). Adicionalmente, algunos factores como dieta deficiente o falta de exposición a la luz solar, pueden causar melanismo temporal y los individuos pueden adquirir una coloración normal en la siguiente muda, una vez eliminada la posible causa del exceso pigmentario (Grouw 2006). Sin embargo, cuando los patrones atípicos son permanentes, la producción de melanina es una consecuencia de alteraciones genéticas (Griffiths et al. 2000).
La ocurrencia de anormalidades pigmentarias en aves puede comprender hasta 4.5% de la población, principalmente en especies insulares que son puras o que presentan el efecto de cuello de botella con poblaciones reducidas; aunque dichas anormalidades pueden alcanzar 13% en condiciones alteradas por impacto antropogénico (particularmente en áreas urbanas o agropecuarias). Esto ha sucedido en poblaciones de golondrinas en los alrededores de Chernobyl, Ucrania, donde la radiación nuclear (nivel siete) ha causado probablemente los mayores niveles de mutación genética con alteración del pigmento de las aves (Nogueira y Alves 2011). Sin embargo, Acosta (2005) reporta un caso de leucismo parcial en el semillero oliváceo (Tyaris olivacea) en cautiverio, donde menciona una frecuencia alta de individuos que presentan anormalidades pigmentarias, sin asignar un valor porcentual; en cambio, E.R. Rodríguez-Ruíz (obs. pers.) estima que más del 50% de individuos en cautiverio del perico australiano (Melopsittacus undulatus) mostró incidencia de anormalidades pigmentarias principalmente por leucismo parcial, el cual es nombrado comúnmente como “perico australiano mariposa” y altamente apreciado por su valor ornamental por el humano e interés comercial en diversas regiones de México.
La incidencia de casos cercanos a áreas perturbadas sugiere un aumento aparente de especies e individuos con variación en la pigmentación, posiblemente por el estrés ambiental al que se enfrentan y que es ocasionado, en su mayoría, por diversas actividades humanas como la agricultura y ganadería, la industria y la alta urbanización (Rodríguez-Ruíz et al. 2014), lo que se manifiesta en trastornos de la expresión genética (Møller y Mousseau 2001, Nogueira y Alves 2011), estrés fisiológico (Philips 1954) y desórdenes alimenticios (Grouw 2006). Determinar aquellos factores naturales o artificiales involucrados en estos padecimientos podría contribuir a determinar las poblaciones que presentan problemas genéticos a corto plazo (González-Arrieta y Zuria 2015).
Finalmente, se espera que el uso de la clave dicotómica propuesta sea incorporado en la identificación de anormalidades pigmentarias, como una herramienta continua y apropiada en contextos modernos. Sin embargo, el manejo y resultado de esta clave dependerá de aquello que se desee identificar, por lo que sugerimos observar detalladamente los caracteres morfológicos principales correspondientes a cada opción de la clave y esquematizarlos.

