










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHuitzil
versión On-line ISSN 1870-7459
Huitzil vol.16 no.1 Omitlán ene./jun. 2015
Artículo original
Urban birds in the Sonoran Desert: estimating population density from point counts
Aves urbanas en el Desierto Sonorense: estimando densidades poblacionales a partir de conteos de puntos
Karina Johnston López,1 Alberto Macías Duarte2* y Reyna Amanda Castillo Gámez1
1 Departamento de Investigaciones Científicas y Tecnológicas de la Universidad de Sonora. Luis Donaldo Colosio s/n, entre Sahuaripa y Reforma, Colonia Centro, Hermosillo, Sonora, 83000, México.
2 Unidad Académica Hermosillo, Universidad Estatal de Sonora. Ley Federal del Trabajo s/n, Colonia Apolo, Hermosillo, Sonora, 83100, México. *Email: alberto.macias@ues.mx
Recibido: 20 de marzo de 2014
Revisión aceptada: 21 de octubre de 2014
Editora asociada: Claudia E. Moreno Ortega
Abstract
We conducted bird surveys in Hermosillo, Sonora using distance sampling to characterize detection functions at point-transects for native and non-native urban birds in a desert environment. From March to August 2013 we sampled 240 plots in the city and its surroundings; each plot was visited three times. Our purpose was to provide information for a rapid assessment of bird density in this region by using point counts. We identified 72 species, including six non-native species. Sixteen species had sufficient detections to accurately estimate the parameters of the detection functions. To illustrate the estimation of density from bird count data using our inferred detection functions, we estimated the density of the Eurasian Collared-Dove (Streptopelia decaocto) under two different levels of urbanization: highly urbanized (90-100% of urban impact) and moderately urbanized zones (39-50% of urban impact). Density of S. decaocto in the highly-urbanized and moderately-urbanized zones was 3.97±0.52 and 2.92±0.52 individuals/ha, respectively. By using our detection functions, avian ecologists can efficiently relocate time and effort that is regularly used for the estimation of detection distances, to increase the number of sites surveyed and to collect other relevant ecological information.
Keywords: Eurasian Collared-Dove, Streptopelia decaocto, detection function, distance sampling, point count, Hermosillo, Sonora.
Resumen
Realizamos conteos de aves en Hermosillo, Sonora, usando muéstreos de distancias para caracterizar las funciones de detección en trayectos por punto para las aves urbanas, nativas y no nativas, en un ecosistema desértico. De marzo a agosto de 2013 muestreamos 240 puntos en la ciudad y sus alrededores; visitamos cada punto tres veces. Nuestro propósito fue proveer información para agilizar la evaluación de la densidad de aves de la región utilizando puntos de conteo. De las 72 especies que identificamos, seis no son nativas. En 16 especies obtuvimos suficientes registros para estimar, de forma adecuada, los parámetros de las funciones de detección. Para ilustrar la estimación de la densidad por medio de puntos de conteo utilizamos las funciones de detección inferidas y estimamos la densidad de la tórtola turca (Streptopelia decaocto) en dos distintos niveles de urbanización: alto (90-100% de impacto urbano) y moderado (39-50% de impacto urbano). La densidad de S. decaocto en las zonas altamente urbanizadas y en las moderadas fue de 3.97±0.52 y 2.92±0.52 individuos/ha, respectivamente. Si los ecólogos utilizan nuestras funciones de detección podrán administrar su tiempo y esfuerzo, por lo regular dedicado a estimar las distancias de detección, para incrementar el número de sitios muestreados y colectar otra información ecológicamente relevante.
Palabras clave: tórtola turca, Streptopelia decaocto, función de detección, muestreo de distancias, puntos de conteo, Hermosillo, Sonora.
Introduction
According to the Secretaría del Medio Ambiente y Recursos Naturales (Semarnat), from 1993 to 2002, 2.8 million hectares changed from natural ecosystems to urban and rural areas (Semarnat 2011). Urban areas are in constant expansion, causing a massive and unpredictable environmental change (McDonnell and Pickett 1990, Chance and Walsh 2004, McKinney 2008, Fuller et al. 2009). For some animals, urbanization may imply new food sources or nesting sites, but for others, urbanization may imply new predators or competitors, and depleted resources (Marzluff 2001, Blair 2001, Shochat 2004). Ecologists are interested on how the urban expansion affects native wildlife populations, particularly the avifauna (Marzluff et al. 2001a, Pennington and Blair 2012).
Birds are particularly adaptable to urban environments. In fact, some native and non-native species have developed dense and stable populations within urban areas (Blair 2001, Pennington and Blair 2012). Some native species, such as the American Crow (Corvus brachyrhynchos), the Black-billed Magpie (Pica pica), and the Greater Roadrunner (Geococcyx californianus) have become even more abundant inside cities than in their natural ecosystems (Marzluff et al. 2001b, Jerzak 2001, Schoech and Bowman 2001, DeStefano and Webster 2012). Some non-native species such as the House Sparrow (Passer domesticus), the Rock Pigeon (Columba livia), the European Starling (Sturnus vulgaris), and the recently introduced Eurasian Collared-Dove (Streptopelia decaocto) have expanded their distribution along with urbanization (Mennechez and Clergeau 2001, Blair 2001, McKinney 2006, Olden 2006, Fuller et al. 2009). Both native and non-native populations should continuously be monitored to detect population trends. Therefore, estimating abundance is a critical step towards their management and conservation. Increasing our knowledge of urban birds is also relevant to promote the awareness of citizens on the natural heritage of their surroundings.
It is important to rely on sampling methods that effectively estimate density (individuals per unit of area). Bird counts (number of birds detected in a given area) do not account for imperfect detection, this is, when individuals are overlooked during sampling. Therefore, counts can only obtain indices of relative abundance (Conroy and Carroll 2009). A popular count technique in ornithology is point counts (Ralph et al. 1995), where an observer records all birds seen and heard for a defined period of time at a point with a fixed radius. Distance sampling is a widely used method employed to estimate bird density (Bibby et al. 1998, Gregory et al. 2004, Buckland et al. 2001, Conroy and Carroll 2009) accounting for imperfect detection. In contrast to counts, distance sampling consists on counting individuals and measuring their distance from the observer, under the assumption that all birds at a zero distance are detected. Distance sampling has two forms: line-transect sampling and point-transect sampling. Line-transect sampling consists of counting organisms and measuring their distance while walking along a line. Line-transect sampling is not usually suitable for urban areas since the random allocation of sampling units is difficult given the frequent occurrence of obstacles such as building or streets. Point-transect sampling consists on measuring distances from every detected bird to a fixed observer during a defined period of time within a given radius. Therefore, point-transect sampling is more appropriate for urban areas (Buckland et al. 2001).
Detection distances are used to infer the detection function g(x), i.e., the probability to detect a bird at a given distance (x) from the observer (Buckland et al. 2001). The characterization of the detection function is the key to estimate density accounting for imperfect detection. Once a detection function is obtained from the distance data, the detection function can be used to estimate density in future point counts (i.e., without measuring distances). However, no one seems to take advantage of using detection functions from previous studies (e.g., Norvell et al. 2003, Somershoe et al. 2006, Kark et al. 2007, Newson et al. 2008, Fuller et al. 2008, Fuller et al. 2009, Loss et al. 2009, Walker and Shochat 2010). Researchers usually calculate their own detection functions when they need to estimate densities, instead of devoting their time and effort in other relevant aspects of their studies.
The objective of this study was to characterize detection functions of several native and non-native species present in the city of Hermosillo, Sonora, through an exhaustive bird sampling. By providing these detection functions, researchers interested in studying urban bird communities in similar habitats will be able to estimate population density (Thomas et al. 2010) from simple point counts without spending time and effort on recording detection distances. In addition, we exemplify the use of detection functions to estimate density from point count data. We do this in the context of a relevant aspect to be studied in urban environments: the increasing abundance of several exotic bird species which possibly compete and displace native species. In this regard, the recent invasion of the Eurasian Collared-Dove may require population monitoring since several native dove species could suffer from resource competition from this exotic species (Romagosa 2012). Therefore, we use records from our study to illustrate our suggestion of future uses of our detection functions.
Methods
Study area
Hermosillo, Sonora (29°08'N, 110°96'W; 211 m altitude) is a medium-size city (population 784,342; INEGI 2012) within the Mexican portion of the Sonoran Desert. The city has a hot desert climate (BWh in the Kóppen climate classification), with 200 mm of mean annual rainfall. Hermosillo reaches high temperatures in summer (>40 °C) and winter can bring temperatures as low as 0 °C (annual average temperature of 25 °C).
Survey method
The study was conducted during the breeding season of 2013 (March to August). We divided this breeding season in three sampling periods: March-April, May-June and July-August. We visited the same sampling points each time. We used a grid of 440 quadrants of 1 km2 defined by the UTM grid that covers the entire urban area of Hermosillo and its surroundings. Following the urban-rural gradient categorization proposed by McDonnell and Pickett (1990) and recommended by other authors (e.g., Marzluff et al. 2001a; Pennington y Blair, 2012), we gave each quadrant one of four levels of urbanization (0-29%, 30-59%, 89-60% and 90-100% of urban impact) corresponding to its percent cover of buildings and streets. The levels of urbanization were estimated using satellite images from Google Earth (2013) and from the Inventario Nacional de Viviendas (updated to 2012, INEGI 2013) at each quadrant. From the total of 440 quadrants, 60 quadrants were randomly selected to sample, 15 quadrants of each level of urbanization. We randomly positioned four plots within each square. A total of 240 plots distributed within the city's boundaries were visited each period. We sampled each plot for 8 minutes using 200 m fixed-radius point transects (Buckland et al. 2001) between 05:40 h and 09:30 h, just after sunrise and before the temperature reached more than 30 °C. Sampling was performed by a two-person team, the observer was the same during the entire sampling period. Birds disturbed during the observer's arrival were recorded at their initial location. We recorded individuals and groups of individuals (clusters hereafter) detected, as well as their species, cluster size and detection distance from the observer. We measured each detection distance (in meters) using a Nikon Prostaff Laser 440 Rangefinder®. We recorded all birds seen and heard, while birds flying through the sampling point were neither recorded nor considered for the analysis.
Data analysis
We estimated parameters of detection functions for species with >75 detections (Buckland et al. 2001), using program DISTANCE (V 6.0 release 2, Thomas et al. 2010). We adjusted three key functions (half-normal, hazard-rate and uniform), each with three expansion adjustments (cosine, simple polynomial and hermit polynomial) with up to four terms. We chose the model with lowest value of the Akaike's Information Criterion (AIC) as the most plausible (best) model. We used detection distances below the 90th percentile to reduce the impact of outliers and improve the model fit.
To illustrate the use of our detection function (built with all detection distances in our three sampling periods) using count data to estimate density, we used our records on S. decaocto from two levels of urbanization, highly-urbanized zones (90-100% of urban impact) and moderately-urban zones (30-59% of urban impact) during the third sampling period (when the species was more abundant within the breeding season). The highly-urbanized zones differed from the moderately-urbanized zones because of the heavy vehicular traffic, tall and dense buildings and poor vegetation structure. Moderately-urbanized had lower vehicular traffic, sparser and smaller buildings (mostly houses), few unpaved streets and a medium vegetation structure with several native plants. We used data that can be easily obtained from bird counts: clusters detected (n) and the number of plots surveyed (k). For the highly-urbanized level of urbanization we used the records obtained in 60 plots and for the moderately-urbanized level of urbanization we used the records obtained in 55 plots.
Among all species identified we used count data from S. decaocto given that the specie is one of the most common species in Hermosillo and it is important to monitor this species continentally. Streptopelia decaocto is a recently-introduced (1980) avian species to the American continent that has been able to expand its distribution along most of America quite rapidly. Recent studies have encountered S. decaocto from southern Canada, all over the United States and Mexico, down to Central America (Romagosa 2012). The rapid and increasing distribution of S. decaocto is occasionally attributed to the continuous urban expansion (Crooks y Soule 1999). Several studies have identified that S. decaocto inhabits areas that present some degree of urban impact (Johnston, 2001; Bonter et al. 2010).
The estimator of density  is given by the expression (Buckland et al. 2001):
is given by the expression (Buckland et al. 2001):

where ĥ(0) is the estimator of the derivative of the probability density function of detection distances evaluated at zero distance, direct output of program DISTANCE, and 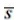 is the average number of individuals per detection. The standard error of density SE , assuming a Poisson distribution of counts, can be approximated using the delta method as follows (Buckland et al. 2001):
is the average number of individuals per detection. The standard error of density SE , assuming a Poisson distribution of counts, can be approximated using the delta method as follows (Buckland et al. 2001):

where SE(ĥ(0)) also a direct output of program DISTANCE and Var() is the sampling variance of the average number of individuals per detection. Finally, we used our estimates of density and their standard errors to test statistical differences in density of S. decaocto between highly-urbanized and moderately-urbanized zones using a Wald test (McCulloch et al. 2008).
Results
We recorded a total of 11,024 individuals from 72 species (Table 1) during the breeding season of 2013 in 240 point transects with three replicates. According to each species' account in The Birds of North America (BNA), this 72 species include 39 resident breeders, 12 summer breeders, and 21 migrants (Poole 2013). Ten species comprised 80% of the individuals detected (Figure 1). We found six non-native species inhabiting the city of Hermosillo, with Paser domesticus being the most abundant, followed by Streptopelia decaocto, Columba livia, Sturnus vulgaris, and the parrots Myiopsitta monachus and Amazona albifrons. Order Columbiformes is the most abundant taxa, represented by 5. decaocto, C. livia, Zenaida asiática, Z. macroura, and Columbina inca; Columbiformes comprises 40% of the individuals detected. Native species of high relative importance in the urban bird community included (in decreasing order) Zenaida asiática, Z. macroura, Columbina inca, Haemorhous mexicanus, Auriparus flavipens, Quiscalus mexicanus, and Melanerpes uropygialis (Figure 1). Two identified species are included in the Red List of Threatened Species (IUCN 2013), Vireo bellii as near threatened and ToxoStoma bendirei as vulnerable.

We had 16 species with sufficient detections to carry out the analysis and model a best fitting detection function (Table 2). Uniform key functions with cosine adjustments and hazard-rate key functions were superior for modeling detection functions in our bird community compared to models with half-normal and hermit and polynomial adjustments.
The best detection function for S. decaocto was the uniform key with a 4th order cosine adjustment, ĥ(0) = 11.086 ha1 (Table 2, Figure 2).
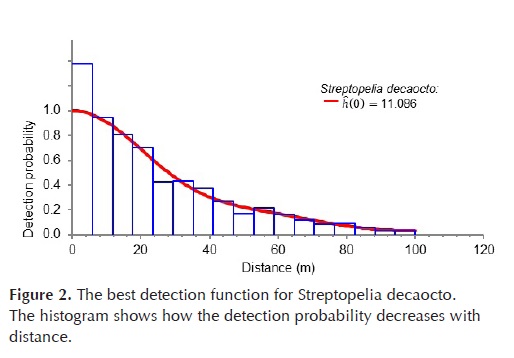
We will now illustrate the use of the inferred detection functions to transform counts into density estimates. In the highly urbanized zones we had n = 122 detections of S. decaocto in k = 60 plots, and an average cluster size per detection of = 1.111 individuals. With the estimate ĥ(0) = 11.09 ha-1 (Table 2) we estimated a density id) = 3.97 individuals/ha. With a Var() = 0.007, and SE(ĥ(0)) = 0.97 ha1 (Table 2), we obtained and SE = 0.52 individuals/ha and a 95% CI(D) = (2.96, 4.98).
In the moderately-urbanized zones we had n = 63 detections of S. decaocto in k = 55 plots, with an average cluster size per detection of = 1.44 individuals. With the estimate ĥ(0) = 11.09 ha 1 we estimated a density = 2.92 individuals/ha. With a Var() = 0.007, and having SE(ĥ(0)) = 0.97/ha, we obtained SE = 0.52 individuals/ha and a 95% CI(D) = (2.92, 3.94).
The Wald test did not show enough evidence of statistical differences in density between these two zones (z = 1.43, P = 0.15).
Discussion
The bird community of the city of Hermosillo shares some dominant components with other urban bird communities in the Sonoran Desert and the world. For instance, Paser domesticus (exotic) is also the most abundant species in urban Tucson (Emlen 1974) and Phoenix (Green and Baker 2003) in Arizona, USA; native and exotic columbids (pigeons and doves) also attain high relative abundances. Large body mass and granivorous diets, attained by these dominant species, are traits that have related to success in urban areas (Lancaster and Rees 1979, Beissinger and Osborne 1982, Ikin et al. 2 002, Kark et al. 2007, Johnston-López 2014). Furthermore, reduced bird diversity in urban environments, as revealed in this work by the existence of few species comprising the majority of the bird abundance (Figure 1), is a widespread pattern (Emlen 1974, Beissinger and Osborne 1982, Mills et al. 1989, Green and Baker2003). More predictable food sources and reduction in predation risk by top predators in urban areas compared to the surrounding arid environments are thought to change trophic relationships that ultimately lead to the dominance of fewer urban specialists (Faeth et al. 2005) with important exotic component such as Paser domesticus, Columba livia and, more recently, Streptopelia decaocto. Urbanization also reduces the insectivorous component of the native bird community in the surrounding desert (Walker and Shochat 2010), such as Campylorhynchus brunneicapillus and Polioptila melanura, which inhabit the less urbanized areas in Hermosillo (Johnston-López 2014).
The study of urban bird community dynamics requires reliable estimation of bird abundance to make reliable inferences in the ecological processes that shape structure and composition of those communities. Point counts are one of the most popular methods for estimating abundance of birds (Ralph et al. 1998, Rosenstock et al. 2002). Studies that conduct point counts can cover larger land extensions and can be conducted by a larger group of people, since only identification training is needed. Some coordinated survey efforts organize annual bird point counts, such as The North American Breeding Bird Survey, and encourage citizen participation. These bird counts collect important information about population trends and promote avian conservation, but they do not account for imperfect detection, and the information cannot be used to correctly estimate densities.
Studies that estimate densities by distance sampling usually rely on visual estimation of distances to save time when collecting the survey data. However, the use of visually-estimated distances (e.g., Norvell et al. 2003, Fuller et al. 2009, Loss et al. 2009, Walker and Shochat 2010) is questionable. Distance sampling analyses assume that all distances are estimated accurately (Buckland et al. 2001). Distances are also frequently measured in ranges (e.g., Somershoe et al. 2006, Kark et al. 2007, Fuller et al. 2008, Newson et al. 2008). Accurately measuring distances will always be preferred to visually estimating densities, even obtaining them by pacing distances. The use of detection functions inferred from previous distance data could help avoid having to rely on uncertain distances to estimate densities. By using these detection functions to estimate densities, ecologists could relocate their time and effort employed in estimating distances and readjusting their estimating capacity to obtain other important details from their studies, such as environmental information or additional counts. For the city of Hermosillo and other cities in the Sonoran Desert, we recommend that research on urban avifaunas use point counts, considering all birds seen and heard (except for those just flying by) within a radial distance of 200 m from the observer and then use our estimates of ĥ(0) to convert those counts into density. All bird surveys were conducted by a single observer (KJL) during the breeding season of 2013. Therefore, we did not have the need of accounting for differences between observers and seasons when estimating detection functions. Although our detection functions are representative of trained surveyors, further evaluations of variability due to observers is needed.
Variation of our detection functions among species are evidently due to variation in species detectability, which is quantified by the estimates of h(0). The parameter h(0) is inversely proportional to the effective area (v), that is h(0) = 2π/v. In our case, v is the inner circle within the 200 m-radius plot, such that the number of birds detected outside it (but within the 200 m-radius plot) equals the number of birds undetected within it (Buckland et al. 2001). Relatively large v then represents a better coverage of the 200 m-radius sampling plot by the surveyor. Species such as Columba livia, Tyrannus verticalis, Quiscalus mexicanus and Zenaida macroura that tend to perch high on buildings, electric poles and cables in large groups, and Cardinalis sinuatus that sings loud while perched on high branches, are all easily detectable at distances over 60 m (some are easily detected over 150 m) and attain the lowest h(0) estimates among all species (Table 2) and therefore, have relatively large effective areas. Species such as Zenaida asiática and Streptopelia decaocto also tend to perch high on electric poles and cables but individually or in small groups; Melanerpes uropygialis and Campylorhynchus brunneicapillus tend to sing loud when approached; and Chondestes grammacus are all easily detected over 30 m (some are easily detected even at 100 m); these species attain relatively low estimates of h(0) (Table 2) and therefore also have fairly large effective areas. On the other hand, species such as Haemorhous mexicanus, Passer domesticus, Columbina inca, and Auriparus flaviceps that are relatively abundant and some even sing quite loud, are not easily detectable at relatively large distances because of their small body size. These species are mostly detected under 60 m and attain intermediate h(0) estimates among the species we analyzed (Table 2) and therefore, have small-medium effective areas. Finally, species such as Polioptila melanura and Cynanthus latirostris are frequently hard to detect because of their tiny body size and their constant "hopping-flying around" behavior. These species are mostly detected under 40 m and therefore attain large h(0) estimates (Table 2) and small effective areas.
Using our detection function results from point counts readily produce relevant management information in terms of population density. In particular, density estimates for Streptopelia decaocto using the detection function suggests that the species attains similar abundances in highly-urbanized zones and moderately-urbanized zones. This species has been recorded in both urban and agricultural areas (Johnston 2001, Fuller et al. 2009) and rarely in natural lands. Fuller et al. 2009 studied the urban birds within the boundaries of the city of Sheffield (England), a 160 km2 city with temperate climate. Their population estimate for the entire city was 13,271 (95%CI = (11,312, 15,568)) individuals of S. decaocto during the summer (May-July). Our population estimate for the species in the city of Hermosillo (Mexico), a 193 km2 city with arid climate was 57,384 (95% CI = (51,484, 63,284)) individuals during the summer (March-August). We are not aware of any estimate of the species' density by levels of any urban impact classification anywhere else for comparison to our estimates. Fujisaki et al. (2010) mention that the range expansion of this species tends to follow human altered landscapes such as road and agricultural areas, but the authors do not have enough data to conclude whether S. decaocto was more abundant in urban or suburban areas. Several studies have reported this species in urban areas of several Mexican states (Pineda-Lopez and Malagamba2011, Chablé-Santos et al. 2012, Pineda-Lopez et al. 2013), but none of these studies documents the changes in the distribution of this species along an urban-rural gradient.
Acknowledgments
We thank our field assistants R.R. Valle-Salinas and F. Johnston-López for their unconditional help during the whole summer. We also thank A.G. Martinez-Vargas and an anonymous reviewers for their helpful comments during the manuscript preparation. K. Johnston-López has a National Council of Science and Technology (Conacyt) graduate scholarship (No. 273604).
Literature cited
Beissinger, S.R. and D.R. Osborne. 1982. Effects of urbanization on avian community organization. The Condor 84:75-83. [ Links ]
Bibby, C., M. Jones, and S. Marsden. 1998. Expedition Field Techniques: Bird Surveys. Expedition Advisory Centre, Royal Geographical Society. London, United Kingdom. [ Links ]
Blair, B.B. 2001. Creating a homogeneous avifauna. Pp. 459-486. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Bonter, D.N., B. Zuckerberg, and J.L. Dickinson. 2010. Invasive birds in a novel landscape: habitat associations and effects on established species. Echography, 33:494-502. [ Links ]
Buckland, ST., D.R. Anderson, K.P. Burnham, J.L. Laake, D.L. Borchers, and L. Thomas. 2001. Introduction to distance sampling: estimating abundance of biological populations. Oxford University Press. Oxford, United Kingdom. [ Links ]
Chablé-Santos, J., E. Gómez-Uc, and S. Hernández-Betancourt. 2012. Registros reproductivos de la paloma de collar (Streptopelia decaocto) en Yucatán, México. Huitzil 13:1-5. [ Links ]
Chance, J.F. and J.J. Walsh. 2004. Urban effects on native avifauna: A review. Landscape and Urban Planning 74:46-69. [ Links ]
Conroy, M.J. and J.P. Carroll. 2009. Quantitative conservation of vertebrates. Wiley-Blackwell. Chichester, United Kingdom. [ Links ]
Crooks, J.A. and M.E. Soule. 1999. Lag times in population explosions of invasive species: causes and implications. Pp. 103-125. In: O.T. Sandland, P.J. Schei, and A. Viken (eds). Invasive Species and Biodiversity Management. Dordrecht: Kluwer Academic Publishers. London, United Kingdom. [ Links ]
DeStefano, S. and C.M. Webster. 2012. Distribution and habitat of Greater Roadrunners in urban and suburban Arizona. Pp. 155-165. In: C.A. Lepczyk and PS. Warren (eds.). Urban Bird Ecology and Conservation. University of California Press. Berkeley, California, USA. [ Links ]
Green, D.M. and M.G. Baker. 2003. Urbanization impacts on habitat and bird communities in a Sonoran desert ecosystem. Landscape and urban planning 63(4):225-239 [ Links ]
Emlen, J.T. 1974. An urban bird community in Tucson, Arizona: derivation, structure, regulation. The Condor 76:184-197. [ Links ]
Faeth, S.H., P.S. Warren, E. Shochat, and W.A. Marussich. 2005. Trophic dynamics in urban communities. Bioscience 55(5):399-407. [ Links ]
Fujisaki, I., E.V Pearlstine, and F.J. Mazzotti. 2010. The rapid spread of invasive Eurasian Collared Doves Streptopelia decaocto in the continental USA follows human-altered habitats. Ibis 152:622-632. [ Links ]
Fuller, R.A., J. Trátalos, and K.J. Gaston. 2009. How many birds are there in a city of a half a million people? Diversity and Distributions 15:328-337. [ Links ]
Fuller, R.A., RH. Warren, RR. Armsworth, O. Barbosa, and K.J. Gaston. 2008. Garden bird feeding predicts the structure of urban avian assemblages. Diversity and Distributions 14:131-137. [ Links ]
Google (on line). 2013. Google Earth Version 6.2 Software. Available at <www.google.com/earth/> (accessed January 2013). [ Links ]
Gregory, R.D., D.W. Gibbons, and P.F. Donald. 2004. Bird census and survey techniques. Pp. 17-56. In: W. J. Sutherland, I. Newton, and R. E. Green (eds.). Bird Ecology and Conservation: A Handbook of Techniques.Oxford University Press. Oxford. [ Links ]
Ikin, K., E. Knight, D.B. Lindenmayer, J. Fischer, and A.D. Manning. 2012. Linking bird species traits to vegetation characteristics in a future urban development zone: implications for urban planning. Urban Ecosystems 15:961-977. [ Links ]
INEGI (Instituto Nacional de Estadística y Geografía) (on line). 2012. Perspectiva Estadística Sonora Diciembre 2012. Available at <www.inegi.org.mx/prod_serv/contenidos/espanol/bvinegi/productos/integracion/estd_perspect/son/Pers-son.pdf> (last accessed 1st March 2014). [ Links ]
INEGI (Instituto Nacional de Estadística y Geografía) (on line). 2013. Inventario Nacional de Viviendas (actualizado 2012). Available at <www3.inegi.org.mx/sistemas/mapa/inv/Default.aspx> (accessed January 2013) [ Links ]
IUCN (International Union for Conservation of Nature) (online). 2013. IUCN Red List of Threatened Species version 2013.1. Available at <www.iucnredlist.org> (last accessed 28th October 2013). [ Links ]
Jerzak, L. 2001. Synurbanization of the magpie in the Paleartic. Pp. 403-425. In: J. M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Johnston, R.F. 2001. Synanthropic birds of North America. Pp. 49-67. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Johnston-López, K. 2014. Estructura de la comunidad de aves en un ecosistema urbano en el Desierto Sonorense. M.Sc. thesis. Universidad de Sonora. Hermosillo, Sonora. [ Links ]
Kark, S., A. Iwaniuk, A. Schalimtzek, and F. Banker. 2007. Living in the city: can anyone become an 'urban exploiter'? Journal of Biogeography 34:638-651. [ Links ]
Lancaster, R.K. and William E. Rees. 1979. Bird communities and the structure of urban habitats. Canadian Journal of Zoology 57(12):2358-2368. [ Links ]
Loss, S.R., M.O. Ruiz, and J.D. Brawn. 2009. Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biological Conservation 142:2578-2585. [ Links ]
Marzluff, J.M. 2001. Worldwide urbanization and its effects on birds. Avian ecology and conservation in an urbanizing world. Pp. 19-48. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Marzluff, J.M., R. Bowman, and R. Donnelly. 2001 a. History of urban bird research. Pp. 1-17. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Marzluff, J.M., K.J. McGowan, R. Donnelly, and R.L Knight. 2001b. Causes and consequences of expanding American Crow populations. Pp. 331-363. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
McCulloch, C. E., S. R. Searle, and J. M. Neuhaus. 2008. Generalized, Linear, and Mixed Models. Wiley John & Sons. Hoboken, New Jersey, USA. [ Links ]
McDonnell, M.J. and S.T.A. Pickett. 1990. Ecosystem structure and function along urban-rural gradients: An unexploited opportunity for ecology. Ecology 71:1232-1237. [ Links ]
McKinney, M.L. 2006. Urbanization as a major cause of biotic homogenization. Biological Conservation 127:247-260. [ Links ]
McKinney, M.L. 2008. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosystems 11:161-176. [ Links ]
Mennechez, G. and P. Clergeau. 2001. Settlement of breeding European Starlings in urban areas: Importance of lawn vs. anthropogenic waste. Pp. 275-287. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Mills, OS., J.B. Dunning, Jr., and J.M. Bates. 1989. Effects of urbanization on breeding bird community structure in southwestern desert habitats. The Condor 9:416-428. [ Links ]
Newson, S.E., K.L. Evans, D.G. Noble, J.J. Greenwood, and K.J. Gaston. 2008. Use of distance sampling to improve estimates of national population sizes for common and widespread breeding birds in the UK. Journal of Applied Ecology 45:1330-1338. [ Links ]
Norvell, R.E., F.P. Howe, and J.R. Parrish. 2003. A seven-year comparison of relative-abundance and distance-sampling methods. The Auk 120:1013-1028. [ Links ]
Olden, J.D. 2006. Biotic homogenization: a new research agenda for conservation biogeography. Journal of Biogeography. 33:2027-2039. [ Links ]
Pennington, D.N. and R.B. Blair. 2012. Using gradient analysis to uncover pattern and process in urban bird communities. Pp. 9-31. In: C.A. Lepczyk and PS. Warren (eds.). Urban Bird Ecology and Conservation. University of California Press. Berkeley, California, USA. [ Links ]
Pineda-López, R. and A.R. Malagamba 2011. Nuevos registros de aves exóticas en la ciudad de Querétaro, México. Huitzil 12:22-27. [ Links ]
Pineda-López, R., A.R. Malagamba, I. Arce-Acosta, and J.A. Ojeda-Orranti. 2013. Detección de aves exóticas en parques urbanos del centro de México. Huitzil 14:56-67. [ Links ]
Poole, A. (ed.) (on line). 2013. The Birds of North America Online. Cornell Laboratory of Ornithology, Ithaca, NY. Available at <bna.birds.cornell.edu/BNA/> (last accessed on 1st March 2014). [ Links ]
Ralph, C.J., J.R. Sauer, and S. Droege (eds.). 1995. Monitoring bird populations by point counts. General Technical Report PSW-GTR-149. Pacific Southwest Research Station. Albany, California, USA. [ Links ]
Romagosa, C.M. 2012. Eurasian Collared-Dove (Streptopelia decaocto). The Birds of North America Online (A. Poole, ed.). In: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online: <bna.birds.cornell.edu/bna/species/630> (last accessed 1st March 2014). [ Links ]
Rosenstock, S.S., D.R. Anderson, K.M. Giesen, T. Leukering, and M.F. Carter. 2002. Landbird counting techniques: current practices and an alternative. The Auk 119:46-53. [ Links ]
Schoech, S.J. and R. Bowman. 2001. Variation in the timing of breeding between suburban and wild land Florida Scrub-Jay: Do physiological measures reflect different environments? Pp. 289-306. In: J.M. Marzluff, R. Bowman, and R. Donnelly (eds.). Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers. London, United Kingdom. [ Links ]
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). 2011. Estrategia Nacional del Manejo Sustentable de Tierras. México, DF. [ Links ]
Shochat, E. 2004. Credit or debit? Resource input changes population dynamics of city slicker birds. Oikos 106:622-626. [ Links ]
Somershoe, S.G., D.J. Twedt, and B. Reid. 2006. Combining breeding bird survey and distance sampling to estimate density of migrant and breeding birds. The Condor 108:691-699. [ Links ]
Thomas, L., S.T. Buckland, E.A. Rexstad, J.L. Laake, S. Strind-berg, S.L. Hedley, J.R.B. Bishop, T.A. Marques, and K. P. Burnham. 2010. Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology 47:5-14. [ Links ]
Walker, J.S. and E. Shochat, E. 2010. Scalar effects of vegetation on bird communities in an urbanizing desert ecosystem. Urban Ecosystems 13:155-167. [ Links ]

