










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHuitzil
versão On-line ISSN 1870-7459
Huitzil vol.13 no.1 Omitlán Jan./Jun. 2012
Científica
Influencia de la asimetría de las rectrices exteriores en la eficiencia de vuelo del Rabihorcado (Fregata magnificens)
Influence of the outer rectrices asymmetry in flight efficiency of the Magnificent Frigatebird (Fregata magnificens)
Mónica González-Jaramillo1,2,3 y Horacio de la Cueva1,3*
1 Departamento de Ecología Marina, Centro de Investigación Científica y Educación Superior de Ensenada. Carretera Ensenada-Tijuana No. 3918, Zona Playitas, Apartado Postal 360, Ensenada, Baja California, 22860, México. Correo electrónico: *cuevas@cicese.mx.
2 Departamento de Ecología y Sistemática Terrestres, El Colegio de la Frontera Sur. Av. Rancho Polígono 2a, Parque Industrial Lerma, Campeche, Campeche, 24500, México.
3 PO Box 434844, San Diego, CA, 92143-4844, EUA.
Recibido: 2 de agosto de 2011.
Revisión aceptada: 10 de enero de 2012.
Editor asociado: Eduardo Palacios Castro.
Resumen
La asimetría de las rectrices exteriores en Fregata magnificens (rabihorcado) influye en su eficiencia de vuelo, mientras que la cola, profundamente bifurcada de esta especie sexualmente dimórfica, juega un papel crucial en su desempeño aerodinámico. Para evaluar la presión de la selección natural en la asimetría de la cola, modelamos el desempeño de rabihorcados machos y hembras simétricos y asimétricos adultos durante el vuelo de planeo en la colonia reproductiva de Isla Isabel, México, mediante la proporción levante:arrastre y las polares y superpolares de planeo. Encontramos diferencias significativas entre rabihorcados machos y hembras, simétricos y asimétricos cuando analizamos su desempeño en el vuelo de planeo y sus costos de compensación asociados. El modelo teórico indicó que la asimetría de la cola tendrá un efecto más grande en los machos que en las hembras durante el vuelo, y que los individuos asimétricos volarán menos eficientemente que los simétricos corrigiendo su asimetría en función de la velocidad al incorporar más costos de arrastre. Investigación futura para estimar las tasas de supervivencia de individuos simétricos y asimétricos, costos de construcción de nido y de éxito reproductivo podría revelar las ventajas de tener una cola simétrica en esta especie y elucidar si la asimetría tiene un costo en la adecuación.
Palabras clave: asimetría de la cola, ave marina, desempeño de vuelo, selección natural.
Abstract
The asymmetry of the outer rectrices in Fregata magnificens (Magnificent Frigatebird) has an influence on flight efficiency, while the deeply forked tail of this sexually dimorphic species plays a crucial role in its aerodynamic performance. In order to assess natural selection pressure on tail asymmetry, we modelled gliding flight performance of symmetric and asymmetric adult frigatebirds at the breeding colony of Isla Isabel, Mexico, through lift:drag ratio and glide polars and superpolars. We found significant differences between symmetric and asymmetric male and female frigatebirds when we analyzed gliding flight performance and flight compensation costs. Our theoretical model pointed that outer rectrices asymmetry has a greater effect on males than on females during gliding flight, and that flight efficiency of symmetric individuals will be higher than asymmetric ones, which will correct asymmetry according to flight velocity by adding drag costs. Further research in survival rates of tail symmetric and asymmetric males, nest construction costs, and breeding success could reveal the advantages of having a symmetric tail in this species and elucidate if asymmetry has a fitness cost.
Keywords: tail asymmetry, seabird, flight performance, natural selection.
Introducción
La función de las rectrices exteriores, por su prominencia, variabilidad intraespecifica y supuesto reflejo de desarrollo somático han sido extensivamente estudiadas, atribuyéndola en ocasiones a la selección natural para incrementar la eficiencia aerodinámica de los individuos (Norberg 1994, Evans 1998, 1999) y otras veces, a la selección sexual como un indicador de calidad (Møller 1991, 1992).
La cola de las aves tiene al menos tres funciones aerodinámicas: (1) mantener la estabilidad aerodinámica del ave, (2) control longitudinal o direccional en maniobras rápidas y (3) producción de fuerza de levante (Thomas 1995). La fuerza de levante actúa perpendicularmente y de manera ascendente al curso de vuelo del ave, su principal función es contrarrestar la fuerza de gravedad que determina el peso del ave (Pennycuick 1989). La fuerza de levante es adicional a la de las alas e incrementa el área de soporte sin incrementar la envergadura (distancia entre las puntas de las alas extendidas) durante el vuelo lento, despegue y aterrizaje, así como en la aceleración y durante los virajes (Norberg 1990).
Las aves con colas profundamente bifurcadas (colas con las rectrices exteriores más largas que las centrales) tienen la desventaja de que las rectrices exteriores son particularmente susceptibles al crecimiento asimétrico y a sufrir daños (Evans y Thomas 1992, Balmford et al. 1993, Thomas 1993a, 1995). La asimetría de las rectrices exteriores se puede deber a un patrón de muda asincrónico, desgaste, desgarre, ruptura o puede ser el resultado de la asimetría fluctuante (desviaciones aleatorias de la simetría bilateral que aparece cuando un individuo no desarrolla por igual ambos lados del cuerpo (Møller y Höglund 1991, Bjorksten et al. 2000). Dichas causas "naturales" de asimetría resultan en una diferencia en la longitud de las rectrices exteriores de las aves (González-Jaramillo y de la Cueva 2010). Independientemente del origen o de la causa de asimetría, las aves con colas asimétricas deben realizar las mismas actividades de vuelo durante la temporada reproductiva (forrajeo, recolección de material para la construcción de nidos, etc.) que las aves con colas simétricas, lo que implica un mayor costo aerodinámico.
La magnitud del efecto de la asimetría de las rectrices exteriores en el desempeño aerodinámico reflejado en la maniobralidad, agilidad, aceleración y velocidad en vuelo horizontal depende de qué tanto se abre (extiende) la cola durante el vuelo (Balmford et al. 1993, Thomas 1993a, 1993b, Evans 1998). La maniobralidad mide el radio mínimo de vuelta ejecutado en vuelo sin perder velocidad y la agilidad mide la facilidad o rapidez con que se puede alterar el curso de vuelo (Norberg y Rayner 1987). En vuelo rápido, la cola se cierra generando poca o ninguna fuerza de levante o arrastre, mientras que en vuelo lento la cola se abre ampliamente para incrementar la fuerza de levante generada en las alas (Thomas 1993a). Así, el arrastre (fuerza que actúa paralelamente al flujo de aire incidente; Pennycuick 1989) a cualquier velocidad es proporcional al área de la superficie expuesta de la cola (Thomas 1993a). Puesto que los cambios en las fuerzas aerodinámicas en una parte del animal afectan la aerodinámica de las otras partes, el costo de la asimetría puede ser distribuido, pero no eliminado (Thomas 1993a, Norberg 1994). Este costo aerodinámico adicional debe ser compensado por las aves con rectrices asimétricas para así tener el mismo desempeño en las actividades de vuelo que los individuos simétricos. Además, en aves con dimorfismo sexual en talla, el desempeño aerodinámico aumenta conforme decrece la masa, por lo que machos y hembras suelen tener diferentes capacidades de vuelo (Andersson y Norberg 1981).
El rabihorcado (Fregata magnificens) es un ave marina con notorio dimorfismo sexual en coloración y talla (las hembras son 15% más grandes que los machos; Osorno 1996), y se caracteriza por tener una cola profundamente bifurcada y una envergadura de hasta 2.29 metros (Pennycuick 1983). Los rabihorcados machos y hembras tienen diferentes ciclos reproductivos: los machos se reproducen cada año, mientras que las hembras reproductivamente exitosas lo hacen cada dos años (Diamond 1972, 1973). Esto se debe a que cada sexo tiene diferentes inversiones durante el cuidado parental; el macho deserta el nido después de los primeros 3 meses de crianza y deja a la hembra encargada de la alimentación del pollo hasta por 14 meses más (Osorno 1999, Osorno y Székely 2004). Debido a lo anterior, se ha especulado que en los rabihorcados las capacidades de vuelo son diferentes entre sexos (Pennycuick 1983).
Aunque estas aves pueden volar por periodos de tiempo prolongados remontándose en las termales que se forman sobre el mar (vuelo de remonte), viajando grandes distancias a muy bajo costo energético (Pennycuick 1983, Weimerskirch et al. 2003, 2006), ambos sexos tienen un comportamiento cleptoparasítico (robo de alimento; Brockmann y Barnard 1979) y suelen robar ramas a otros individuos durante la temporada de construcción de nidos (MGJ, obs. pers.), por lo que también pueden exhibir un vuelo activo muy acrobático.
Aunque en algunas especies de aves con cola profundamente bifurcada, como Hirundo rustica, la asimetría de las rectrices exteriores de los machos suele ser usada por las hembras como un indicador de calidad durante la elección de pareja (Møller 1990, 1992, Møller et al. 1998), en los rabihorcados la asimetría de las rectrices exteriores no parece influir ni en la selección de pareja ni en el desempeño del cuidado parental a corto plazo (González-Jaramillo y de la Cueva 2010), por lo que la función de la simetría de las rectrices exteriores en esta especie podría ser principalmente aerodinámica a través de la selección natural y no un indicador de calidad debida a la selección sexual.
Nuestro objetivo fue estimar la influencia de la asimetría de las rectrices exteriores en la eficiencia de vuelo de los rabihorcados machos y hembras mediante el cálculo del costo aerodinámico y su compensación durante el vuelo de planeo y remonte (sin movimiento de alas), usando observaciones en campo y modelos teóricos aerodinámicos para determinar si su función es principalmente aerodinámica debida a selección natural.
Considerando que en esta especie las hembras son más grandes que los machos (Diamond 1972), que existe una relación alométrica entre sexos (v. g. Schmidt-Nielsen 1984, Calder 1996) y que el desempeño aerodinámico está asociado al tamaño (Andersson y Norberg 1981), bajo la hipótesis de que la asimetría de las rectrices exteriores tiene un efecto cuantificable en el desempeño aerodinámico de las aves con colas profundamente bifurcadas, esperamos un efecto en la capacidad de vuelo diferencial entre sexos con las siguientes predicciones: (1) si la asimetría de la cola causa arrastre aerodinámico diferenciado entre ambos lados de la cola, disminuyendo la proporción de la fuerza de levante, lo que representa un costo para el ave (Norberg 1990, Evans y Thomas 1992, Balmford et al. 1993, Thomas 1993b, 1995), predecimos que los machos simétricos volarán más eficientemente que las hembras simétricas. Es decir, la proporción levante:arrastre será mayor en machos simétricos que en hembras simétricas, lo que deberá reflejar en una mejor velocidad de planeo. (2) Si el costo de la asimetría puede ser distribuido en alas y cola, pero no eliminado (Thomas 1993a, Norberg 1994), esperamos que al incorporar en el modelo de vuelo más costos de arrastre a diferentes velocidades en machos y hembras asimétricos, el efecto de la asimetría de la cola disminuya en mayor proporción en machos asimétricos que en hembras asimétricas.
Métodos
Realizamos este estudio de noviembre de 2000 a abril de 2001, durante la temporada reproductiva de los rabihorcados que anidan en el Parque Nacional Isla Isabel ubicado en el Pacifico Central Mexicano (21º52’N, 105º54’O). Isla Isabel es también un "Área de Importancia para la Conservación de Aves en México" (AICA; Drummond et al. 2000) que alberga una colonia reproductiva de entre 15,000-16,000 rabihorcados (Madsen 2005).
Aleatoriamente capturamos de noche y con las manos rabihorcados adultos, machos (n=34) y hembras (n=30) perchados cuya asimetría de la cola desconocíamos, los medimos (±1 mm), pesamos (±25 g) y los liberamos sin causarles daño. Los rabihorcados machos mudan las plumas de la cola de manera continua y el patrón identificado no incluye la muda simultánea de las rectrices exteriores, por lo que la teoría de la asimetría fluctuante que implica un crecimiento simultáneo no se puede aplicar a esta especie (Madsen 2005). Hasta donde sabemos, a la fecha no existe ningún estudio sobre el patrón de muda de las hembras de rabihorcado, por lo que, para efecto de esta investigación, supusimos que ocurre un patrón continuo como en los machos. Todas las medidas corporales y de velocidad de vuelo fueron tomadas por la misma persona para minimizar el error sistemático de medición.
Alas (cm)
Debido a la longitud y fragilidad de las alas de los rabihorcados, no pudimos seguir el método de medición alar sugerido por Pennycuick (1989). En su lugar, medimos la longitud (cm) de diferentes partes anatómicas de ambas alas extendidas (±1 mm): (1) humero, (2) longitud del hombro a la punta de la pluma axilar más larga, (3) ulna, incluyendo el primer dígito; (4) longitud del codo a la punta de la décima pluma remera secundaria, (5) longitud del primer dígito a la punta de la décima pluma remera primaria, (6) longitud del primer dígito a la punta de la primera pluma remera, (7) longitud de la punta de la primera pluma remera a la punta de la décima pluma primaria remera y (8) distancia entre hombros (Figura. 1).
Cola (cm)
Medimos (± 1mm): (1) la longitud de rectrices centrales exteriores desde su nacimiento (quilla) hasta la punta (bifurcación; Figura 2A), (2) la longitud de las rectrices exteriores desde la quilla hasta la punta (Figura 2A), (3) la extensión de la cola, distancia desde la quilla de la rectris exterior derecha a la quilla de la rectris exterior izquierda, mientras la cola estaba completamente extendida (Figura 2B). Controlamos el grado de extensión de la cola presionando ligeramente el rabihorcado contra el pecho de la persona que lo sostenía. En esta posición, la cola de los rabihorcados se extiende completamente.
Cuerpo (cm) y masa (kg)
Medimos (±1 mm) la longitud del cuerpo, desde la punta del pico hasta la punta de la rectris exterior más larga, colocando al ave sobre su dorso y estirando su cuello. Registramos la masa corporal de cada individuo con un dinamómetro (±25 g) suspendiéndolo dentro de una bolsa de tela.
Longitudes y asimetrías de las rectrices exteriores (cm)
Para ambos sexos calculamos: (1) la longitud relativa de la estructura (humero, ulna, mano, rectrices y ala; Figura 1), que es la longitud de la estructura (media de las medidas de la estructura izquierda y derecha), dividida entre la longitud del cuerpo (Balmford et al. 1994); (2) la asimetría absoluta que es la diferencia numérica entre la estructura derecha e izquierda y la asimetría relativa de rectrices exteriores y alas que es la asimetría absoluta dividida entre el tamaño promedio de la estructura (Palmer y Strobeck 1986, Møller y Höglund 1991).
Clasificación de aves simétricas y asimétricas
Puesto que algunas aves no pueden detectar asimetrías menores al 1.25% (Swadle 1999), a que en los rabihorcados la asimetría de la cola per se no parece ser un indicador de calidad (González-Jaramillo y de la Cueva 2010) y a que las asimetrías tienen costos aerodinámicos importantes cuando la asimetría de las rectrices exteriores es de al menos el 6% (Evans et al. 1994), clasificamos a los rabihorcados como asimétricos cuando tenían asimetría mayor al 6% (asimetría absoluta mayor a 2.7 cm).
Longitud de las alas (LA; m)
Suma de las longitudes de las medidas 1, 3 y 5 de cada ala (Figura 1).
Envergadura (b; m)
Distancia entre las puntas de las alas sumando las longitudes de las medidas 1, 3 y 5 (Figura 1) de ambas alas al ancho del cuerpo (medida 8; Figura 1).
Área total alar (S; m2)
Dividimos las alas en cuatro figuras geométricas (Figura 1), cuyas áreas calculamos de las medidas de longitud tomadas en el campo y sumamos el área total proyectada de ambas alas completamente extendidas (Figura 1, áreas B-D) incluyendo el área del cuerpo entre ellas (Figura 1, área A).
Carga alar (Q; Nm2)
Diferencia de la presión promedio entre la superficie inferior y superior del ala cuando el ave esta planeando, medida como peso en Newtons (N) por unidad de área del ala (m2) (Pennycuick 1989). Q =mgS-1, dónde: m =masa (kg), g =aceleración debida a la gravedad (9.81 ms-1) y S =área alar (m2).
Proporción dimensional del ala (Λ)
Es una medida de la forma del ala. Las aves que planean reducen sus proporciones dimensionales en vuelo rápido flexionando el codo y la muñeca (Pennycuick 1989). Mide la proporción entre la envergadura y la profundidad media del ala (Λ=b2S-1). Dónde b=envergadura (m) y S=área alar (m2)
Área de la cola (AC; m2)
Calculamos el área de la cola dividendo la cola en figuras geométricas, calculamos sus áreas a partir de las medidas 1-3 de la Figura 2 y sumamos las áreas calculadas: a+b (Figura 2B).
Velocidades de vuelo y viento (km h-1)
Medimos oportunísticamente las velocidades de vuelo de planeo de 326 rabihorcados cuya asimetría de la cola se desconocía (183 machos, 143 hembras) usando un radar Doppler MPH, K15X990114 (Kansas, EUA). Medimos las velocidades desde la misma ubicación entre dos colinas donde los rabihorcados siempre planean hacia el norte con viento cruzado hacia el noroeste. De esta manera, el error por viento (causado cuando hay diferentes direcciones de viento) se disminuye y al estar quieto el observador, se estandariza el error de paralaje. Así, estandarizamos las medidas y minimizamos el error de medición (de la Cueva y Blake 1997). Medimos la velocidad y dirección del viento usando una estación meteorológica portátil, Water Base 10 WS2103 (EUA), ubicada en la cima del árbol más alto entre las colinas.
Modelo de vuelo
Incluimos en el modelo de vuelo de Pennycuick (1989) las velocidades de vuelo de planeo, las velocidades y dirección del viento, así como los datos de desempeño aerodinámico: masa corporal, envergadura (b), área alar (S), carga alar (Q), proporción dimensional del ala (Λ)y área de la cola (AC). Calculamos la proporción levante:arrastre (L/D) para rabihorcados machos y hembras con colas simétricas y asimétricas para comparar sus costos de desempeño aerodinámico, y la superpolar de planeo, que supone que el ave selecciona su envergadura óptima para obtener un menor arrastre en cada velocidad.
Para modelar la superpolar de planeo y encontrar la mejor velocidad de planeo (Vbg) correspondiente al intervalo máximo de velocidad, primero calculamos las polares individuales de planeo desde que las alas y la cola están completamente abiertas (100%; ß=1.0) hasta que las alas y la cola están parcialmente extendidas (60%; ß=0.4). ß da la menor velocidad de hundimiento (VZ, poder total requerido para volar entre el peso del ave; Pennycuick 1989). Supusimos una distribución igual del peso del ave en ambas alas. Modelamos el costo de planeo de rabihorcados hembras y machos, simétricos y asimétricos para predecir los costos adicionales instantáneos de planeo que las aves asimétricas tienen que superar para alcanzar los mismos resultados que las aves simétricas. Estos costos se demuestran en una menor eficiencia de vuelo de planeo.
Análisis estadísticos
Para comparar la morfometría y los datos de desempeño aerodinámico de los rabihorcados machos y hembras, usamos la prueba no paramétrica U Mann-Whitney de dos colas para muestras con heterocedasticidad, o sus aproximaciones normales (Z) cuando el tamaño de la muestra lo permitió. Para todos los análisis consideramos α=0.05 (Zar 1999). Reportamos medias ± desviación estándar.
Resultados
Proporción de rabihorcados asimétricos
De los 64 rabihorcados medidos, el 23% (n=15) fueron asimétricos; de estos, el 29% (n=10) fueron machos y el 17% (n=5) hembras.
Diferencias aerodinámicas entre machos y hembras
La longitud relativa de alas, cola y la asimetría relativa de la cola en los machos fue significativamente mayor que en las hembras, pero no la asimetría relativa de las alas (Cuadro 1). Las hembras tuvieron un área alar (S), envergadura (b), carga alar (Q) y área de la cola significativamente mayor que los machos, pero la proporción dimensional del ala (Λ), mayor en hembras que en machos, no fue significativa (Cuadro 2).
Obtuvimos las velocidades de vuelo de planeo con vientos del noreste de entre 0 y 7 ms-1. Las velocidades mínimas y máximas de planeo para los machos fueron de 3.9 y 15.8 ms-1 respectivamente, mientras que para las hembras fueron de 4.4 y 11.9 ms-1 respectivamente (Cuadro 3).
Costos de desempeño aerodinámico, proporción levante:arrastre (L/D)
La proporción levante:arrastre (L/D) fue mayor en aves simétricas (Figura 3A y B) que en asimétricas (Figuras 3C y D) de ambos sexos. Las proporciones L/D pronosticadas por el modelo fueron 2.2 veces más grandes en machos simétricos que en asimétricos (Figura 3C). Un macho asimétrico producirá una menor proporción L/D cuando su velocidad se incrementa porque su arrastre aumentará rápidamente con una diferencia máxima a una velocidad horizontal de Vx=10 ms-1 . En el peor caso para las hembras a Vx=12 ms-1 (Figura 3D), la proporción L/D de las asimétricas fue 2.1 veces mayor que la de las simétricas.

Corrección de asimetría y eficiencia de vuelo: polares y superpolares de planeo
De acuerdo a las polares de planeo, para lograr altas proporciones de L/D y de penetración (avance horizontal vs. pérdida de altura) conforme aumenta la velocidad, los rabihorcados reducirán la extensión de su envergadura (b), área alar (S) y área de cola (AC). El modelo teórico indica que los machos serán más eficientes que las hembras en corregir su asimetría durante el planeo al modificar la morfología de las alas y la cola (Figuras 4A y B). Teóricamente, los machos tendrán mejor penetración que las hembras cuando planean, aún si vuelan a bajas velocidades (menos de 10 ms-1). Sin embargo, las hembras podrían descender más rápido que los machos (Figura 4A) y su eficiencia de vuelo de planeo menor les permitiría perder altura a una tasa más alta al incrementar su velocidad de vuelo o disminuir sus áreas de alas y cola.

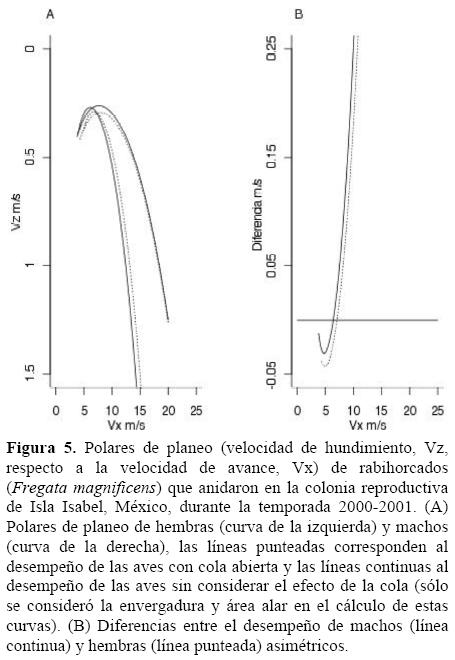
Como se esperaba, las superpolares de planeo indican que los rabihorcados simétricos serán más eficientes que los asimétricos en el desempeño del planeo (Figuras 5A y B) y que la asimetría de la cola tiene un efecto más importante en los machos que en las hembras. Debido a las diferencias morfológicas entre sexos, el modelo teórico indica que las velocidades óptimas de planeo (Vbg) de machos y hembras serán diferentes (5 ms-1 y 10 ms-1, respectivamente; Figura 5A). Un rabihorcado planeando con asimetría extrema perderá verticalmente entre 2.5 ms-1 (hembra) y 7.5 ms-1 (macho) a una velocidad de entre 15 y 20 ms-1 (Figura 5B). Es decir, un macho asimétrico podrá perder hasta 67% más altura que una hembra asimétrica si vuela a una velocidad de entre 15 y 20 ms-1. Mientras que las hembras perderían menos de 2 ms-1 a una velocidad de vuelo de 20 ms-1 con la cola abierta, los machos casi 4 ms-1 (Figura 5A). Al eliminar el efecto de la cola abierta en el vuelo de los machos, entonces la velocidad de hundimiento aumentó a casi 5 ms-1.
Discusión
Nuestros resultados sugieren que la asimetría de las rectrices exteriores de los rabihorcados influye en su eficiencia de vuelo y por lo tanto, la selección natural parece ser la responsable del desarrollo de una cola simétrica. De acuerdo a nuestras predicciones, el modelo teórico indica que la asimetría de la cola tiene un efecto más importante en los machos que en las hembras durante el vuelo, siendo los individuos asimétricos menos eficientes durante el vuelo que los simétricos, aunque los machos asimétricos pueden corregir su asimetría más eficientemente que las hembras asimétricas en función de la velocidad al incorporar más costos de arrastre.
Diferencias aerodinámicas entre machos y hembras
De acuerdo al modelo, los machos pueden alcanzar un amplio intervalo de velocidades de vuelo, planear más eficientemente y pueden compensar con mayor eficiencia la asimetría de sus rectrices exteriores que las hembras. Esto se puede explicar en parte a que la longitud relativa de las alas es mayor en machos que en las hembras, reduciendo así el arrastre. La carga alar (Q) menor de los machos y su proporción dimensional del ala (Λ) les permite una tasa más grande de ascenso que en las hembras (Pennycuick 1983) y reduce su velocidad de hundimiento y el costo de vuelo durante el vuelo de planeo lento. Sin embargo, tanto la asimetría relativa como la longitud relativa de la cola fueron menores en hembras que en machos y por lo tanto también los costos de compensación.
Debido a que los machos se reproducen cada año y las hembras cada dos años (Diamond 1972, 1973), y a que principalmente los machos llevan el material para la construcción del nido y compiten por él contra conespecíficos, mientras que las hembras deben alimentar a los pollos después de la deserción del macho (Osorno 1996, 1999), se ha sugerido que el papel que desempeña cada sexo durante la crianza junto con el dimorfismo sexual influyen en las diferentes capacidades y tipos de vuelo en cada sexo: un vuelo ágil y de gran maniobralidad en machos, y vuelos de campo traviesa (cross-country) en hembras (Pennicuick 1983). La eficiencia aerodinámica es proporcionada no sólo por la simetría de su cola, sino también por la carga alar (Q) y dimensión proporcional del ala (Λ). La carga alar (Q) de las hembras, mayor que en los machos, les permite planear lentamente y junto con la dimensión proporcional del ala (Λ) reduce su arrastre (Pennycuick 1983). Así, probablemente los machos enfrentan un alto costo de compensación por una cola asimétrica durante la construcción del nido y la alimentación del pollo durante el cuidado biparental, pero después de la deserción del macho las hembras requieren que su plumaje de vuelo este en condiciones óptimas para evitar costos extra cuando se quedan a cargo de la crianza. Durante el cuidado biparental, las hembras invierten en la crianza de manera proporcional a los machos, pero cuando este deserta ellas duplican su tasa de alimentación para compensar la ausencia de su pareja (Osorno y Székely 2004). Lo anterior tiene implicaciones en el desempeño aerodinámico que podrían explicar la proporción de individuos simétricos menor en machos que en hembras; aunque esta hipótesis debe ser probada.
El modelo de vuelo indica que la asimetría fluctuante de la cola tendrá un efecto mayor en el desempeño aerodinámico de los machos que en las hembras debido a que las diferencias en la asimetría y longitud relativa de la cola son mayores en los machos. La asimetría en la cola causa arrastre aerodinámico disminuyendo la fuerza de levante, lo que representa un costo para el ave (Norberg 1990, Evans y Thomas 1992, Balmford et al. 1993, Thomas 1993b, 1995). El costo aerodinámico de la asimetría absoluta y relativa se incrementa con la longitud de la cola, pero mientras que el costo de la asimetría absoluta aumenta de manera lineal, el costo de la asimetría relativa lo hace exponencialmente (Thomas 1993a).
Por otro lado, la longitud relativa de las alas de los machos resultó mayor que en las hembras, lo que ayuda a corregir la asimetría de su cola y, en consecuencia, a reducir el costo debido al arrastre. Se ha visto que dentro de las familias de aves de cola larga, el dimorfismo sexual de la cola está asociado con el dimorfismo sexual de las alas para reducir el costo aerodinámico (Balmford et al. 1994).
Los machos tuvieron un intervalo de velocidades de vuelo más amplio y, de acuerdo con el modelo, tendrán un desempeño de vuelo más eficiente al modificar su envergadura y extensión de la cola para compensar la asimetría, lo que resulta en un menor hundimiento a todas las velocidades, es decir, los machos serán más eficientes que las hembras para compensar la asimetría de sus rectrices exteriores. Además, debido a que la agilidad de vuelo de un ave aumenta conforme decrece la masa (Andersson y Norberg 1981), los machos serán también más ágiles que las hembras debido a que tienen tamaño y masa menores.
Influencia de la asimetría de la cola en el desempeño aerodinámico
De acuerdo con el modelo de vuelo, las diferencias morfológicas de tamaño, peso, carga alar (Q), proporción y extensión de la cola, y el planeo y radio de vuelta entre sexos, se podría suponer que en los machos el tipo de forrajeo es principalmente cleptoparasítico, puesto que requieren de maniobralidad y agilidad, en contraste con un forrajeo de pesca en las hembras, donde es importante el planeo y la velocidad de picada o pérdida de altura. Sin embargo, en Isla Isabel, los rabihorcados son en su mayoría oportunistas (Calixto-Albarrán y Osorno 2000) y una gran proporción de los ataques cleptoparasíticos son realizados por hembras y juveniles con un éxito del 67% una vez que provocan la regurgitación (Osorno et al. 1992). De acuerdo con el modelo, las hembras podrían ser mejores planeadoras que los machos y los machos podrían tener mayor agilidad, pero ambos sexos tendrían la misma capacidad de maniobralidad. La agilidad, maniobralidad y eficiencia de los machos para modificar su envergadura y extensión de la cola, los hacen aptos tanto para forrajear en el mar como para cleptoparasitar a presas de menor tamaño (Andersson y Norberg 1981).
Cómo ocurre en los machos, una carga alar (Q) baja y una dimensión proporcional del ala (Λ) grande, permite una tasa de ascenso alta y volar en círculos pequeños (Pennycuick 1983). Asimismo, la combinación de una proporción dimensional del ala (Λ) alta, carga alar (Q) baja y masa corporal baja, resultan en un costo de vuelo bajo y en la posibilidad de planear lentamente con una mínima velocidad de hundimiento (Norberg 1990). Sin embargo, en machos asimétricos el costo de la asimetría relativa de la cola junto con el costo para corregir esta asimetría podrían limitar de manera importante su eficiencia de forrajeo, puesto que existe una relación comprobada entre el comportamiento de forrajeo y el modo de vuelo asociado (Norberg 1990, Thomas 1995).
Durante la temporada de construcción del nido, los machos usan su agilidad y maniobralidad para llevar ramas al nido, ya que son perseguidos por conespecíficos que intentan robar estas ramas. Por lo tanto, cada rama entregada a la hembra en el nido muestra la eficiencia del macho sobre sus atacantes. En promedio, los machos más ágiles podrían estar construyendo nidos más rápidamente; aunque esta hipótesis debe comprobarse en el futuro. Los resultados predecibles del modelo se pueden comprobar en campo si las velocidades de planeo media y la mediana de aves simétricas y asimétricas marcadas se comparan y si se cuantifican los costos de la construcción de nidos en los rabihorcados.
En conclusión, la asimetría de las rectrices exteriores de machos y hembras de rabihorcado influye en su eficiencia de vuelo sugiriendo la acción predominante de la selección natural en el desarrollo de éste carácter. Investigación futura para estimar las tasas de supervivencia de individuos simétricos y asimétricos, costos de construcción de nido y de éxito reproductivo podría revelar las ventajas de tener una cola simétrica en esta especie y elucidar si la asimetría tiene un costo en la adecuación.
Agradecimientos
Gracias a JL Osorno por su asesoría y constante apoyo en todos los sentidos durante el desarrollo de esta investigación; V Madsen nos proporcionó comentarios invaluables a las primeras versiones de éste trabajo. Gracias a JC Pérez por su ayuda en campo. La Comisión Nacional para Áreas Naturales Protegidas (CONANP) nos permitió realizar el trabajo de campo en la Isla Isabel y junto con la Armada de México nos apoyaron con la logística. Agradecemos también la ayuda incondicional de los pescadores de San Blas y Boca de Camichín.
Literatura citada
Andersson, M. y R.A. Norberg. 1981. Evolution of reversed sexual size dimorphism and role partitioning among predatory birds, with a size scaling of flight performance. Biological Journal of the Linnean Society 15:105-130. [ Links ]
Balmford, A., I.L. Jones, y A.L.R. Thomas. 1993. On avian asymmetry: selection for symmetrical tails and wings in birds. Proceedings of the Royal Society of London Series B 252:245-251. [ Links ]
Balmford, A., I.L. Jones y A.L.R. Thomas. 1994. How to compensate for costly sexually selected tails: the origin of sexually dimorphic wings in long-tailed birds. Evolution 48:1062-1070. [ Links ]
Bjorksten, T.A., K. Fowler y A. Pomiankowski. 2000. What does sexual trait FA tell us about stress? Trends in Ecology and Evolution 4:163-166. [ Links ]
Brockmann, H.J. y C.J. Barnard. 1979. Kleptoparasitism in birds. Animal Behaviour 27: 487-514. [ Links ]
Calder, W.A., III. 1996. Size, function, and life history. Dover. New York, EUA. [ Links ]
Calixto-Albarrán, I. y J.L. Osorno. 2000. The diet of the magnificent frigatebird during chick rearing. Condor 102:569-576. [ Links ]
Cueva, H. de la y R.W. Blake. 1997. Flight speed, foraging efficiency, and daily energy budget of the Barn Swallow, Hirundo rustica. Canadian Journal of Zoology 75:1176-1183. [ Links ]
Diamond, A.W. 1972. Sexual dimorphism in breeding cycles and unequal sex ratio in Magnificent frigate-birds. Ibis 114: 395-398. [ Links ]
Diamond, A.W. 1973. Notes on the breeding biology and behavior of the Magnificent Frigatebird. Condor 75: 200-209. [ Links ]
Drummond, H., C. Macías, A. Valiente-Baunet y J.L.Osorno. 2000. Isla Isabel. Pp. 41. In: M. del Coro-Arizmendi y L. Márquez-Valdemar (eds.).Áreas de importancia para la conservación de las aves en México. CIPAMEX, CONABIO, FMCN, CCA. México, DF. [ Links ]
Evans, M.R. 1998. Selection on swallow tail streamers. Nature 394:233-234. [ Links ]
Evans, M.R. 1999. Length of tail streamers in barn swallows. Nature 397:115. [ Links ]
Evans, M.R. y A.L.R. Thomas. 1992. The aerodynamic and mechanical effects of elongated tails in the scarlet-tufted malachite sunbird: measuring the cost of a handicap. Animal Behaviour 43:337-347. [ Links ]
Evans, M.R., T.L.F. Martins y M. Haley. 1994. Tha asymmetrical cost of tail elongation in red-billed streamertails. Proceedings of the Royal Society of London Series B 256: 97-103. [ Links ]
González-Jaramillo, M. y H. de la Cueva. 2010. Natural tail streamer asymmetry in male Magnificent Frigatebirds Fregata magnificens: influence on mate selection and male parental care performance. Marine Ornithology 38: 85-90. [ Links ]
Madsen, V. 2005. Female mate choice in the Magnificent Frigatebird (Fregata magnificens). Tesis doctoral, Universidad Nacional Autónoma de México. México, DF. [ Links ]
Møller, A.P. 1990. Male tail lenght and female mate choice in the monogamous swallow Hirundo rustica. Animal Behaviour 39:458-465. [ Links ]
Møller A.P. 1991. Sexual ornament size and the cost of fluctuating asymmetry. Proceedings of the Royal Society of London Series B 243:59-62. [ Links ]
Møller, A.P. 1992. Female swallow preference for symmetrical male sexual ornaments. Nature 357: 238-240. [ Links ]
Møller, A.P. y J. Höglund. 1991. Patterns of fluctuating asymmetry in avian feather ornaments: implications for models of sexual selection. Proceedings of the Royal Society of London Series B 245:1-5. [ Links ]
Møller, A.P., A. Barbosa, J.J. Cuervo, F. de Lope, S. Merino y N. Saino. 1998. Sexual selection and tail streamers in the Barn Swallow. Proceedings of the Royal Society of London Series B 265:409-414. [ Links ]
Norberg, R.A. 1994. Swallow tail streamer is a mechanical device for self-deflection of tail leading edge, enhancing aerodynamic efficiency and flight manoeuvrability. Proceedings of the Royal Society of London Series B 257:227-233. [ Links ]
Norberg, U.M. 1990. Vertebrate flight. Springer-Verlag. Berlin, Alemania. [ Links ]
Norberg, U.M. y J.M.V. Rayner. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactionsof the Royal Society of London Series B 316:335-427. [ Links ]
Osorno, J.L. 1996. Evolution of breeding behavior in the Magnificent Frigatebird: copulatory pattern and parental investment. Tesis doctoral, University of Florida. Gainesville, Florida, EUA. [ Links ]
Osorno, J.L. 1999. Offspring desertion in the Magnificent Frigatebird: are males facing a trade off between current and future reproduction? Journal of Avian Biology 30:335-341. [ Links ]
Osorno, J.L. y T. Székely. 2004. Sexual conflict and parental care in Magnificent Frigatebirds: full compensation by deserted females. Animal Behaviour 68:337-342. [ Links ]
Osorno, J.L., R. Torres y C. Macías-García. 1992. Kleptoparasitic behavior of the Magnificent Frigatebird: sex bias and success. Condor 94:692-698. [ Links ]
Palmer, A.R. y C. Strobeck, C. 1986. Fluctuating asymmetry: measurement, analysis, pattern. Annual Review of Ecology and Systematics 17:391-421. [ Links ]
Pennycuick, C.J. 1983. Thermal soaring compared in three dissimilar tropical bird species, Fregata magnificens, Pelecanus occidentalis, and Coragyps atratus. Journal of Experimental Biology 102:307-325. [ Links ]
Pennycuick, C.J. 1989. Bird flight performance: a practical calculation manual. Oxford University Press. New York, EUA. [ Links ]
Schmidt-Nielsen, K. 1984. Scaling. Why is initial size so important? Cambridge University Press. Cambridge, Reino Unido. [ Links ]
Swaddle, J.P. 1999. Limits to lenght asymmetry detection in starlings: implications for biological signalling. Proceedings of the Royal Society of London Series B 266: 1299-1303. [ Links ]
Thomas, A.L.R. 1993a. The aerodynamic costs of asymmetry in the wings and tail of birds: asymmetric birds can’t fly round tight corners. Proceedings of the Royal Society of London Series B 254:181-189. [ Links ]
Thomas, A.L.R. 1993b. On the aerodynamics of birds’ tails. Philosophical Transactions of the Royal Society of London Series B 340:361-380. [ Links ]
Thomas, A.L.R. 1995. On the tails of birds. Lund University. Lund, Suecia. [ Links ]
Weimerskirch, H., O. Chastel, C. Barbraud y O. Tostain. 2003. Frigatebirds ride high on thermals. Nature 421:333-334. [ Links ]
Weimerskirch, H., M. Le Corre, F. Marsac, C. Barcraud, O. Tostain y O. Chastel. 2006. Postbreeding movements of frigatebirds tracked with satellite telemetry. Condor 108:220–225. [ Links ]
Zar, J.H. 1999. Biostatistical analysis, 3a ed. Prentice Hall. New Jersey, EUA. [ Links ]

