











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Helminths (parasitic worms) do not form a monophyletic group; the assemblage consists of members of the phyla Platyhelminthes, Nematoda, Acanthocephala and Annelida (subclass Euhirudinea). Helminths parasitize all classes of vertebrates and invertebrates (Pérez-Ponce de León et al., 2011). They are important as parasites of wild and domestic animals, and they also provide ecological, biogeographical, and evolutionary information on host-parasite relationships (Horwitz & Wilcox, 2005; Mejía-Madrid, 2013).
Approximately, 402 species of amphibians have been described from Mexico (Frots, 2018; Parra-Olea et al., 2014), of which only 66 have been studied from a helminthological perspective; this number represents only about 18% of all Mexican species (Cabrera-Guzmán et al., 2021). Most of those works have focused mainly on taxonomic descriptions, lists of species, or population level studies (Cabrera-Guzmán, 2002; Cabrera-Guzmán et al., 2007, 2010; Galicia-Guerrero et al., 2000; Goldberg & Bursey, 2002; León-Règagnon et al., 2005; Velázquez-Urrieta & León-Règagnon, 2018), and only a few reports of the ecological structure of helminth communities (Adán-Torres et al., 2018; Espínola-Novelo et al., 2017; Goldberg et al., 2001; Mata-López et al., 2013; Paredes-Calderón et al., 2004; Ruiz-Torres et al., 2017; Zelmer et al., 2004). The only helminthological studies of amphibians from Hidalgo state are the helminth fauna of Ambystoma velasci (Dugès, 1888), Lithobates spectabilisHillis and Frost, 1985 and Rhinella horribilis (Wiegmann, 1833) (Falcón-Ordaz et al., 2014, 2015; Pulido-Flores et al., 2009; Rodríguez-Amador et al., 2013). However, most of the works in Hidalgo have focused on a single locality and none have carried out the analysis of the structure of helminths communities.
Lithobates spectabilis (showy leopard frog) is endemic to the central region of Mexico. Adults inhabit rivers, streams, lagoons, and ponds in ecosystems of mesophilic forests, coniferous forests, sub-montane scrub, and xerophilous habitats with an altitudinal interval of 1,200 m and 3,000 m. They are relatively small, with an average snout to cloaca length (SVL) of 69 mm. Common food habits include organisms belonging to the orders Coleoptera, Hymenoptera, Hemiptera, Diptera and Odonata (Canseco-Márquez & Gutiérrez-Mayén, 2010; Hillis & Frost, 1985; Ramírez-Bautista et al., 2014).
The aim of this study was to analyze the structure of the helminth community at the infracommunity level (community of helminth infrapopulations in a single species of host) and at the component community level (all infrapopulations of helminths associated with some subset of a host species) (Bush et al., 1997) in order to better understand the patterns and processes of the helminth communities of L. spectabilis from 2 localities, separated mainly by altitude, from the Biosphere Reserve Barranca de Metztitlán, Hidalgo, Mexico. Both localities have supported populations of L. spectabilis for more than 15-20 years and provide an ideal setting for a comparative analysis of the population structure of helminths (Pulido-Flores et al., 2009).
Materials and methods
The Biosphere Reserve Barranca de Metztitlán is in the east-center part of the state of Hidalgo (20°14’15” N, 98°22’59.99” W). The topography is rugged, consisting of a river (Metztitlán) with steep mountains rising on each side; elevations in the reserve are between 1,000 and 2,000 m asl (Conanp, 2003; INEGI, 2013). The 2 study localities were: Las Pilas (20°28’17.27” N, 98°40’51.27” W), at an altitude of 1,300 m (a large pool formed by a low-water dam on Metztitlán River) with shore vegetation cover consisting mainly of pecan trees, Carya illinoinensis (Wangenh.) K. Koch, 1869, and secondary vegetation; and Metznoxtla pond (man-made) (20°37’46.43” N, 98°51’27.60” W) at an altitude of 1,600 m (mountain top above Las Pilas), with vegetation predominantly of submontane scrub, with coverage between 80 and 90% and a height up to 2 m (Conanp, 2003). The straight-line distance between the 2 localities is about 25 km and the altitude difference is 300 m.
Frogs (40 from Las Pilas and 37 from Metznoxtla pond) were collected during September, October, and November (2017), and January, February, April, June and August (2018). The animals were captured using a hand net during the first hours of night at the 2 localities. Frogs were collected from the low bank of the river, near water but not in it at Las Pilas, and at Metznoxtla they were found sitting in shallow water near the shore of the pond. Animals were taken to the laboratory where they were sacrificed with an overdose of ether. Frogs were dissected and the body cavity and organs were examined for helminths. All helminths were collected and fixed for morphological study using current techniques for each group (Pritchard & Kruse, 1982). Platyhelminthes and acanthocephalans were stained with Mayer’s Carmalum, Delafield’s hematoxylin and Gomori’s trichromic and were mounted on slides in balsam of Canada. Nematodes were sacrificed and fixed in Glacial Acetic Acid, mounted in temporary mounts in glycerin for morphological study, and then transferred to 70% ethanol for storage. Specimens were examined using a compound optical microscope (Leica CME) and drawings were made with the aid of a drawing tube. Specimens were identified using appropriate scientific literature (Anderson, 2000; Anderson et al., 2009; Bray, 2008; Chabaud, 1978; Gibbons, 2010; Gibson et al., 2002; Petrochenko, 1971; Yamaguti, 1961, 1963, 1971).
Data analysis
For this study, the sizes of frogs from each locality were compared in a one-way ANOVA using the statistical program Past V. 4.06b (Hammer et al., 2001).
For characterization of the infection, prevalence, abundance, mean intensity, and range of intensity were quantified according to Bush et al. (1997) and Bautista-Hernandez et al. (2015). To evaluate potential correlations between abundance of helminths species and the snout-vent length of the frogs from each sex and locality, a Pearson correlation test was performed, and dispersion diagrams were plotted in RStudio v. 2022.07.2 (RStudio, 2020). The method of Nested Analysis of Variance (ANOVA) was used with the sex factor of the frogs. Analyses were performed using STATISTICA 10 (StatSoft, 2011).
For the analysis of the data at the infracommunity level, only species richness (using the order of diversity q = 0) (Hill, 1973), and the average abundance in samples for each locality and species diversity were calculated using the order of diversity q = 1 (Hill, 1973; Jost, 2006). To compare the diversity at the infracommunity level with that reported for other anurans, the Brillouin Index (HB) also was used because it is considered ideal for fully-censused communities, and it measures the homogeneity of the community in a manner that is sensitive to the presence of rare species and is moderately sensitive to the size of the sample (Magurran, 2004); furthermore, it provides a comparison index for previous studies which did not use Hill numbers. For the component community analysis, the sample coverage (C) was calculated for each locality.
Hill numbers measure the proportion represented by the individuals of each species in the sample with respect to the total number of individuals (Chao & Jost, 2012). The species accumulation curve was calculated using the method of rarefaction due to sample coverage (Chao & Jost, 2012) in the statistical program R (R Core Team, 2021) with tools from the iNEXT package (Hsieh et al., 2016). The total number of individuals from each of the locations was extrapolated to double and the analysis was carried out with 100 randomizations (Chao et al., 2014).
For the analysis of the diversity the effective number of species, Hill numbers ( q D = w), or the true diversity, were used (q = 0 = species richness; q = 1 = the exponential of Shannon’s entropy index, and q = 2 = the inverse of Simpson’s concentration index) (Chao et al., 2014). Hill numbers are a mathematically unified family of diversity indices which are indicators (standardized values) that quantify the number of species that are considered in the sample being analyzed, depending on the common and rare species (Chao et al., 2014; Hill, 1973; Jost, 2006). These standardized metrics incorporate relative abundance and species richness based on the value of the exponent “q”, The value of q = 0 is insensitive to the differences in the relative abundances of species (S), which is equal to the species richness ( 0 D). The value of q = 1 includes all species with a weight exactly proportional to their abundance in the community; this is Shannon´s diversity index. This can be interpreted as the effective number of common species in the assemblage ( 1 D). The value of q = 2 considers the dominant species in the assemblage ( 2 D) (Hill, 1973; Jost, 2006; Moreno et al., 2011). These measures express the effective number of species that a virtual community would have in which all the species were equally common, conserving the average relative abundance of the studied community (Jost, 2006; Moreno et al., 2011). As the q values increase, more emphasis is placed on the dominant species while conserving the relative abundance of each species. The 3 Hill numbers were calculated according to Hill (1973). The calculation of sample coverage and the Hill numbers were performed in the statistical program R (R Core Team, 2021) with the iNEXT package (Hsieh et al., 2016). To determine patterns in the distribution of the abundance of the species of helminths and to identify the dominant species in the 2 localities, rank-abundance curves were calculated using the Excel program (Avolio et al., 2019).
The difference in the species composition was evaluated at the community level using a similarity analysis (ANOSIM). To represent the general pattern of dissimilarity in the composition of species between the 2 localities, a Non-Metric Multidimensional Scaling was performed (NMDS). Both methods were based on the dissimilarity index of Bray-Curtis (Bray & Curtis, 1957; Jongman et al., 1995) and were performed using the statistical R program (R Core Team, 2021) with tools of the Vegan package (Oksanen et al., 2018).
Beta diversity (β-diversity) is represented as the difference in species composition between different communities, which is determined by species turnover (species replacement) and nestedness (richness difference) (Baselga, 2010; Carvalho et al., 2012). The total dissimilarity of helminths between the 2 localities was divided into the dissimilarity due to species replacement (hypothesized replacement) and dissimilarity due to differences in richness (nestedness) by means of the beta diversity partition method (POD approach) (Podani & Schmera, 2011). This method is based on the Jaccard Index (βjac) and is represented by decomposition of additive terms that allow the consideration of the relative importance of species replacement components (β-3) and richness difference components (βrich) in the configuration of the structure with incidence data based (Podani & Schmera, 2011) as follows: βjac = β-3 + βrich, where: βjac = total dissimilarity, β-3 = dissimilarity due to species replacement components, and βrich = dissimilarity due to richness difference components. The analysis of the data was performed in the statistical program R (R Core Team, 2021) with the BAT package (Biodiversity Assessment Tools; Cardoso et al., 2015) using abundance data (Cardoso et al., 2015; Carvalho et al., 2013).
To determine if there was a significant difference in the values of prevalence and abundance of the shared species (helminths occurring in both localities), the non-parametric Chi-square test (χ2) was used (McDonald, 2009; Plackett, 1983). This test determines if 2 variables are related or not; the analysis was performed using the STATISTICA 10 program (StatSoft, 2011).
Results
The total sample of 77 frogs, L. spectabilis, was comprised of 40 individuals from Las Pilas (28 females-SVL = 6.2-13.5 cm, mean = 8.9 cm and 10 males- SVL = 4.8-8.7 cm, mean = 7.3 cm) (data on sex and size of 2 frogs were not recorded), and 37 individuals from Metznoxtla (16 females- SVL = 2.9-9.4 cm, mean = 5.92 cm and 21 males- SVL = 4-8.1 cm, mean = 6.07 cm). A total of 1,346 helminths were recovered, corresponding to 12 species, 6 of which were identified to species level and 6 could be identified only to genus level (Table 1); 7 were adults and 5 were larvae. Of the frogs collected, 92.5% (n = 37) from Las Pilas and 97.29% (n = 36) of the frogs from Metznoxtla were infected with at least 1 species of helminth.
Table 1 Prevalence, abundance, mean intensity, and range of intensity of helminths in Lithobates spectabilis from the 2 localities of the Reserva de la Biosfera Barranca de Metztitlán, Hidalgo, Mexico. * = Undescribed species, † = juvenal/larvae, ■ = adults, only females, without symbol = adult.
| Number of helminths | Prevalence (%) | Abundance | Mean intensity | Range of intensity | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species/ Maturity | Las Pilas (LP) | Metznoxtla (M) | LP | M | LP | M | LP | M | LP | M |
| Platyhelminthes | ||||||||||
| Glypthelmins quieta | 183 | 408 | 25 | 51.35 | 4.57 | 11.03 | 18.30 | 31.11 | 1-73 | 1-165 |
| Megalodiscus americanus | - | 257 | - | 75.68 | - | 6.95 | - | 9.18 | - | 1-48 |
| Haematoloechus caballeroi | 10 | 2 | 17.5 | 2.70 | 0.25 | 0.05 | 1.43 | 2 | 1-3 | 2 |
| Halipegus sp.* | - | 97 | - | 40.54 | - | 2.62 | - | 6.47 | - | 1-30 |
| Clinostomum sp.† | - | 1 | - | 2.70 | - | 0.03 | - | 1 | - | 1 |
| Nematoda | ||||||||||
| Foleyellides cf. flexicauda | 79 | 8 | 65 | 10.81 | 1.97 | 0.22 | 3.00 | 2 | 1-8 | 1-5 |
| Rhabdias cf. joaquinensis | 15 | - | 17.5 | - | 0.37 | - | 2.14 | - | 1-4 | - |
| Physaloptera sp.† | 70 | - | 17.5 | - | 1.75 | - | 10 | - | 1-38 | - |
| Falcaustra sp.† | 7 | - | 2.5 | - | 0.17 | - | 7 | - | 7 | - |
| Aplectana sp.▪ | 84 | - | 15 | - | 2 | - | 14 | - | 2-62 | - |
| Acanthocephala | ||||||||||
| Lueheia inscripta † | 120 | 3 | 52.5 | 5.41 | 3 | 0.08 | 5.71 | 0.13 | 1-12 | 1-2 |
| Oncicola sp.† | 2 | - | 2.5 | - | 0.05 | - | 2 | - | 2 | - |
The abundance of species of helminths, with respect to the snout-vent length of male frogs, was not correlated (Las Pilas- t = 1.670, df = 7, p > 0.05; Metznoxtla- t = 1.827, df = 21, p > 0.05). However, the abundance of helminths in female frogs was positively correlated with snout-vent length in Metznoxtla (t = 2.884, df = 13, p < 0.05), but not in Las Pilas (t = -1.073, df = 24, p > 0.05) (Fig. 1, Table 2).
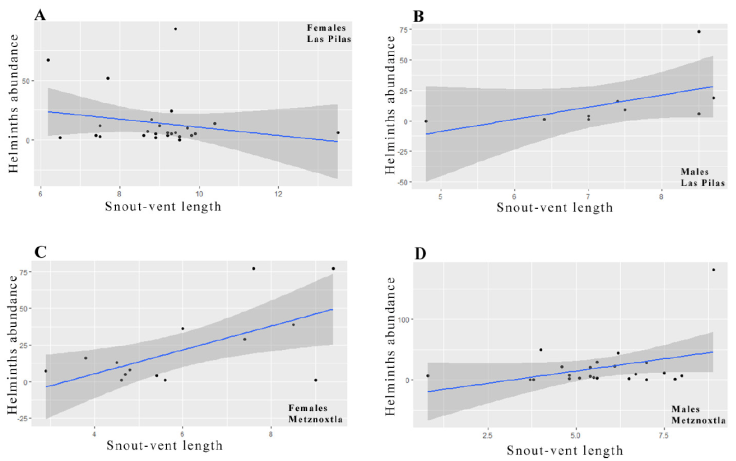
Figure 1 Scatter plots of the abundance of helminths in relation to the snout-vent length of frogs, Lithobates spectabilis. Each graph shows the locality and data of sex and Snout-vent length (SVL).
Table 2 Values of Pearson’s correlation coefficients to evaluate the relationship between abundance of helminths and Snout-Vent Length of Lithobates spectabilis. * = Significant at p < 0.05.
| Locality | Sex | Pearson’s correlation | t | df. | p-value |
|---|---|---|---|---|---|
| Las Pilas | Females | -0.214 | -1.073 | 24 | 0.293 |
| Males | 0.533 | 1.670 | 7 | 0.138 | |
| Metznoxtla | Females | 0.624 | 2.884 | 13 | 0.012* |
| Males | 0.370 | 1.827 | 21 | 0.081 |
In the ANOVA to test for nestedness, the sex of the frogs nested within the locality for snout-vent length variable; length was associated with the locality (SS = 96.502, df = 1, p < 0.05) but not to gender (SS = 10.831, df = 1, p > 0.05). However, frogs from Metznoxtla were significantly smaller than those from Las Pilas, regardless of sex (SS = 49.833, df = 1, p < 0.05). In the test for nestedness in relation to differences in the abundance of helminths in each sex of frogs from each locality, the difference was not significant between localities (SS = 1.001, df = 1, p > 0.05), nor between sexes (SS = 0.459, df = 1, p > 0.05). Although the average abundance was higher in Metznoxtla than Las Pilas, the variation was higher, so the difference was not significant (Table 3).
Table 3 Results of the nested ANOVA for snout-vent length (SVL) (a) and abundance of helminths (b) data.
| Source | Sum of squares | df | Mean square | F-value | p-value |
|---|---|---|---|---|---|
| (a) SVL | |||||
| Locality | 96.502 | 1 | 96.502 | 35.029 | 0.000 |
| Sex | 10.831 | 1 | 10.831 | 3.931 | 0.051 |
| Residuals | 190.091 | 69 | 2.755 | ||
| (b) Abundance | |||||
| Locality | 1.001 | 1 | 1.001 | 0.649 | 0.423 |
| Sex | 0.460 | 1 | 0.460 | 0.298 | 0.587 |
| Residuals | 106.366 | 69 | 1.542 | ||
The most prevalent species of helminth in frogs from Las Pilas was Foleyellides cf. flexicauda (Schacher and Crans, 1973) Esslinger, 1986, present in 26 of 40 frogs, followed by Lueheia inscripta (Westrumb, 1821) Travassos, 1919, present in 21 of 40 frogs, and Glypthelmins quieta (Stafford, 1900) Stafford, 1905, present in 10 of 40 frogs. The species with the lowest prevalence were Falcaustra sp. and Oncicola sp., each present in 1 of 40 frogs. Glypthelmins quieta was the most abundant species, followed by L. inscripta; the mean intensity followed a similar pattern as the abundance (Table 1).
In Metznoxtla, the most prevalent species was Megalodiscus americanus Chandler, 1923, present in 28 of 37 frogs, followed by G. quieta in 19 of 37 frogs and Halipegus sp., present in 15 of 37 frogs. Species with the lowest prevalence were L. inscripta, present in 2 of 37 frogs, followed by Haematoloechus caballeroi (Skrjabin and Antipin, 1962) León-Règagnón and Tabon, 2018, and Clinostomum sp., each presents in 1 of 37 frogs. Glypthelmins quieta was the most abundant species, followed by L. inscripta. The mean intensity followed a similar pattern as the prevalence (Table 1).
Infracommunity diversity, measured by the first Hill number (q = 0 = species richness), in frogs from Las Pilas was 2.27 species of helminths per frog, whereas, in Metznoxtla there were 1.91 species of helminths per frog (Table 4). The second Hill number (q = 1 = the exponential of Shannon’s diversity index = effective species diversity), indicated that Las Pilas had 1.8 effective species per frog and Metznoxtla had 1.6 effective species. Nevertheless, Metznoxtla had a greater mean abundance (20.97) than Las Pilas (14.56).
Table 4 Ecological parameters of helminth infracommunity reported from the genus Lithobates in Mexico. HB = Brillouin diversity index.
| References | Present study | García-Altamirano (1993) | Paredes-Calderón et al. (2004) | Yáñez-Arenas (2007) | Adán-Torres et al. (2018) | |
|---|---|---|---|---|---|---|
| Host | Lithobates spectabilis Hillis et Frost | L. dunni (Zweifel) | L. vaillanti (Brocchi) | L. brownorum (Sanders) | L. montezumae (Baird) | |
| Locality | Las Pilas | Metznoxtla | Lago de Pátzcuaro | Los Tuxtlas | Celestún; Lagunas de Yalahau and Río Lagartos | San Pedro Tlaltizapán |
| State | Hidalgo | Michoacán | Veracruz | Yucatán | Estado de México | |
| Richness | 2.27 (± 1.22)* | 1.91 (± 0.86)* | 3.3 (± 1.5) | 3.49 (± 0.22) | 1.19 (± 0.91) | 2.4 (± 1.1) |
| Diversity (HB) | 0.37 (± 0.32) | 0.31 (± 0.30) | 0.54 (± 0.43) | 1 (± 0.55) | 0.16 (± 0.26) | 0.42 (± 0.36) |
Index used to calculate the average diversity; * Hill numbers (q = 0).
The third Hill number q = 2 does not apply at the infracommunity level, so the Brillouin index (HB) was used (Moreno et al., 2011). The diversity estimated using the Brillouin index in Las Pilas was 0.37 and in Metznoxtla 0.31 (Table 4).
At the level of the component community, in Las Pilas the sample coverage was 100%, and in Metznoxtla the coverage was 99% (Fig. 2); i.e., these numbers indicate that there is little likelihood (0% and 1%, respectively) that more species would have been discovered with an increase in sampling effort. The calculated species diversity (q = 0) at Las Pilas was 9 species of helminth, and 7 species at Metznoxtla; however, extrapolating the accumulation curves the confidence intervals for species richness for the 2 localities overlap, so the differences between the 2 curves are not statistically significant at the component community level (Fig. 2). With the order of diversity = 1 (q = 1), the number of effective species for Las Pilas was 5.73, almost double that of Metznoxtla (2.87 effective species). With the order of diversity q = 2, the same pattern was found; Las Pilas had 4.48 effective species and Metznoxtla had about half as many (2.48 effective species) (Table 5). Glypthelmins quieta was the dominant species in both localities. In Las Pilas, the species with the second highest dominance value was L. inscripta, followed by Aplectana sp.; L. inscripta was present in both localities but Aplectana sp. was present only in Las Pilas. In Metznoxtla, the second most dominant species was M. americanus, followed by Halipegus sp. (Fig. 3); these 2 species were found only in Metznoxtla.

Figure 2 Species accumulation curves of the species of helminth from Lithobates spectabilis from the 2 localities in the Biosphere Reserve Barranca de Metztitlán. Shaded area indicates 95% confidence intervals; continuous lines are interpolations of the model, and the dashed lines are extrapolated values that would be achieved if additional samples were made.
Table 5 Helminth component community diversity of anurans from Yucatán and Hidalgo, Mexico, calculated using the formulas for Hill numbers.
| Host | Rhinella marina | Incilius valliceps | Lithobates spectabilis | |
|---|---|---|---|---|
| Locality | Yalahau, Yucatán, Mexico | Las Pilas | Metznoxtla | |
| Environment | Terrestrial | Aquatic-terrestrial | Aquatic-terrestrial | Aquatic |
| Sample coverage (%) | 100 | 100 | 100 | 99 |
| q = 0 ( 0 D) | 7 | 4 | 9 | 7 |
| q = 1 ( 1 D) | 4 | 2 | 5.73 | 2.87 |
| q = 2 ( 2 D) | 3 | 2 | 4.48 | 2.48 |
| Reference | Espínola-Novelo et al. (2017) | This study | ||

Figure 3 Rank-abundance curves of the species of helminth illustrating the difference in the composition of the component communities of Lithobates spectabilis from Las Pilas and Metznoxtla. Letters represent each species in the curves (a) Glypthelmins quieta; (b) Lueheia inscripta; (c) Aplectana sp.; (d) Foleyellides cf. flexicauda; (e) Physaloptera sp.; (f) Rhabdias cf. joaquinensis; (g) Haematoloechus caballeroi; (h) Falcaustra sp.; (i) Oncicola sp.; (j) Megalodiscus americanus; (k) Halipegus sp.; and (l) Clinostomum sp.
The difference in the species composition between the 2 localities was significant (Anosim; R = 0.54, p < 0.05) and the grouping given by the NMDS corroborates the results of the Anosim; overlapping but partially separated groups of each locality were formed with their respective samples with a high level of reliability (stress 0.098) (Fig. 4). The total beta diversity (βjac) between both localities was 66%, and of this percentage, 50% (β-3) corresponded to the replacement components and 16% (βich) corresponded to richness difference components. The result of the χ2 test of the abundance values was significant (χ2 = 257.36, gl 3, p < 0.05); however, the prevalence values were not significantly different (χ2 = 48.00, gl 49, p > 0.5).

Discussion
Inclusion of helminths in biodiversity studies increases the knowledge of interspecies interactions. Amphibians are opportunistic predators and are prey to a great diversity of organisms; therefore, they function as intermediate, paratenic and final hosts of a great diversity of parasites that have different transmission strategies (Aho, 1990; Campião et al., 2012; Hamann et al., 2010; Imasuen et al., 2012; King et al., 2008). Since most of their lives they are found near the breeding area, amphibians have been considered to be biological markers of local ecological conditions (Aho, 1990). For this reason, they are an excellent model for the study of biological interactions between parasites and hosts (Aho, 1990; Campião et al., 2012). Environmental conditions in which the host develops are of great importance in the transmission dynamics of parasitic species, so changes, or differences, in environmental variable directly affect amphibians. Factors such as the temperature, humidity, salinity, pH and oxygen, as well as the quality of water, local landscape characteristics and the geographical distance between habitat patches are involved in and determine the structure of helminth communities in anurans (Adán-Torres et al., 2018; Bower et al., 2018; de Sena et al., 2018; King et al., 2008; Koprivnikar & Poulin, 2009; Paredes-Calderón et al., 2004; Poulin, 2003).
The Biosphere Reserve Barranca de Metztitlán has extreme altitude gradients that occur between localities only short distances apart and climatic variations throughout the region that corresponds to the protected area. The 2 localities are relatively close together, but in different altitudes, with corresponding differences in microclimate (Conanp, 2003). The distance between Metznoxtla and Las Pilas is about 25 km in a straight line, but the distance between Metznoxtla and Metztitlán River is only about 2.5 km; Las Pilas is in the same river, but about 22 km upstream from that closest point to the river. The difference in altitude between Metznoxtla, on the mountain top, and the river is about 300 m. Consequently, the microclimates are different in each locality.
Las Pilas is in the valley floor, on the banks of Metztitlán River, which functions as a regulator of the local climate and provides the ecological conditions (humidity, shade, etc.) that allows frogs to spend more time out of the water. These conditions allow helminths with heteroxenous life cycles to complete their life cycles. The pond at Metznoxtla is shallow, less than 1 m depth, and in an open area on the mountaintop in a xeric scrub habitat that is warmer, and the trade winds arrive with little humidity and contribute to its semi-dry habitat (Conanp, 2003). There is no shade around the pond, and these conditions cause frogs to spend most of the time in the shallow waters of the pond (Pulido-Flores et al., 2009). This habitat favors digeneans, dominant in this locality, which use snails and tadpoles as intermediate hosts.
Frogs collected in Metznoxtla were captured within the pond, they probably spend most of their time in the water rather than the shore, where there is no shelter, and rarely were seen in shore/terrestrial environments. Because of this, they probably feed mainly on insects found in or near the water, which are more likely intermediate hosts of the digeneans. This behavior could explain the helminth composition and values in the ecological parameters reported for the digeneans, since some of these species use aquatic insect larvae to reach their definitive host and others use aquatic snails as intermediate hosts (Rankin, 1944; Razo-Mendivil & Pérez-Ponce de León, 2008; Stigge & Bolek, 2016). The results of the present study suggest that the physiographic characteristics of each locality are directly involved in producing the differences in the composition of the helminth infracommunity of Lithobates spectabilis.
In Las Pilas, nematodes were the group with the highest species richness (Table 1). Of these, F. cf. flexicauda was the most prevalent species, and the digenean G. quieta was the most abundant. In Metznoxtla, M. americanus, G. quieta and Halipegus sp. were the helminths with the highest values of ecological parameters (Table 1). This suggests that frogs from both localities are consuming insects (intermediate hosts of digeneans and nematodes). However, at Metznoxtla, snails, intermediate hosts of these digeneans, although the life cycle of Halipegus Looss, 1899 also includes dragonflies (Goater et al., 1990). Snails are more obvious at Metznoxtla, and were observed at the shoreline and in shallow waters. As well, G. quieta can be transmitted by cannibalism of infected tadpoles and juvenal frogs and ingesting the shed skin containing encysted metacercaria after metamorphosis (Leigh, 1946). Because of the size and shallowness of the pond, frogs were in proximity sitting in water and on the muddy shore, bringing them in in the vicinity of infective stages of digeneans.
At Las Pilas, frogs were captured mainly on the riverbank, a more terrestrial environment than at Metznoxtla. The higher prevalence of F. cf. flexicauda at Las Pilas (65% vs. 10.81% at Metznoxtla), an obligate parasite of amphibians, could be related to the life cycle of this parasite, which uses mosquitoes of the family Culicidae (Anderson, 2000). No report of mosquitos has been made in the reserve, but mosquitoes were present in the more sheltered riverbank while collecting frogs. Abundance of mosquitoes, and the prevalence of F. cf. flexicauda, suggests that they are feeding constantly on the frogs.
Cystacanths of Lueheia inscripta were found in the mesentery of the frogs; prevalence of 52.5% at Las Pilas and 5.41% at Metznoxtla. Frogs, a paratenic host of this helminth, must eat an infected arthropod intermediate host, thought to be cockroaches (Periplaneta americana Linnaeus, 1758) (Acholonu, 1976) and the parasites re-encyst in the frog, which, in turn, would be eaten by a bird; however, it is not known if this insect is present in these localities. Lueheia inscripta, as adult, is a parasite of the intestines of birds of the families Ardeidae, Icteridae and Turdidae (Salgado-Maldonado & Caspeta-Mandujano, 2010; Smales, 2013; Werby, 1938); all of which have members present in the reserve (Ortiz-Pulido et al., 2010).
Helminths that had the lowest values of the ecological parameters were juveniles of the nematode, Falcaustra sp. and the acanthocephalan, Oncicola sp. Kathlaniid nematodes develop to the third stage outside the host and then invade various species of invertebrates (aquatic insects, snails) and fishes (Bartlett & Anderson, 1985; Hasegawa & Nishikawa, 2009; Moravec et al., 1995), which, along with frogs, serve as paratenic hosts of third-stage larvae that do not develop further in those hosts (Anderson, 2000); large reptiles (lizards, turtles, and snakes) that eat frogs are typical definitive hosts. However, although invertebrates are common in frog’s diets, the low parameter values indicate that the helminths are most likely sporadic in this locality. Acanthocephala of the genus Oncicola Travassos, 1916, involve amphibians and lizards as paratenic hosts (Becker et al., 2019; Nickol et al., 2006), and feline mammals as definitive hosts (Fuller & Nickol, 2011; Santos et al., 2016); although, the specific life cycles have not been thoroughly studied (Nickol et al., 2006; Palmer et al., 2020; Romero-Mayén et al., 2016; Schmidt, 1972). Lithobates spectabilis appears to be a paratenic host of the nematode and this acanthocephalan, although future studies would have to be done to confirm this hypothesis.
The abundance and prevalence of G. quieta and M. americanus in Metznoxtla may be related to the life cycle. Both species use gastropods as the first intermediate host; G. quieta uses Physella gyrina (Say, 1821) and M. americanus uses 1 or several species of Helisoma Swainson, 1840. In both life cycles, the cercaria leave the first intermediate host and swims actively until finding the second intermediate host, tadpoles. They penetrate the epidermis of the tadpoles and form metacercariae in cysts. Adult frogs become infected by ingesting the skin shed during metamorphosis or by feeding on juvenile frogs infected with the metacercaria (Pulido-Flores et al., 2009; Rankin, 1944; Razo-Mendivil & Pérez-Ponce de León, 2008; Schell, 1970). For Halipegus sp., found in the buccal cavity of the frogs, molluscs are the first intermediate hosts, copepods, and ostracods are second intermediate hosts, and odonate larvae, as paratenic hosts, are involved in the life cycle (Krull, 1935; Stigge & Bolek, 2016; Zelmer & Esch, 1999). Frogs, the definitive host, are infected with this helminth by feeding on intermediate or paratenic hosts carrying the metacercaria.
The helminths with lower values of ecological parameters in Metznoxtla were H. caballeroi, parasites of the lungs, Clinostomum sp., found in the cloaca, and L. inscripta, encysted in the mesentery. In the case of H. caballeroi, the parameter values may be influenced by the local abiotic conditions (Metznoxtla is much arid than Las Pilas) and the absence of some undetermined species of potential intermediate host. Species of Clinostomum Leidy, 1856 are not common for amphibians; they normally use fish as intermediate hosts and ictiophagus and piscivorous birds as definitive hosts (Dias et al., 2003). This species was found in the cloaca of a single frog. The acanthocephalan, L. inscripta, thought to use insects as intermediate host and known to mature in birds (Turdidae), was not as common in Metznoxtla (but common in Las Pilas), possibly because of the difference in habits of the frogs (setting in water vs. sitting on the shore) or because either intermediate hosts or infected birds are not common in that locality or because the presence of more vegetation in Las Pilas makes it a better habitat for birds.
At the infracommunity level, Las Pilas had a greater species richness and diversity than Metznoxtla (Table 4). The species richness and diversity in this study is consistent with previous reports for species of Lithobates Fitzinger (Table 4) in Mexico. Yáñez-Arenas & Guillén-Hernández (2010) reported the species richness and diversity of the helminth infracommunity of L. brownorum (Sanders, 1973) in Yucatán, and observed that the parameters were low (Table 4). This condition was attributed to the constant presence of tropical cyclones, which directly or indirectly affect the populations of the definitive and intermediate hosts. The regular perturbation of the habitat by the cyclones was thought to have a considerable influence that would lower the values of richness, diversity, and abundance (Yáñez-Arenas & Guillén-Hernández, 2010). Adán-Torres et al. (2018) suggested that, for the small number of infracommunities studied so far in Nearctic and Neotropical biogeographic regions, it appears that local ecological conditions of the studied localities are more important factors in the structure of helminth community structure of frogs than the biogeographic region is. Furthermore, they considered the limited information available so far was insufficient for them to draw definite conclusions.
Aho (1990) argued that helminth parasite communities of anurans generally are depauperate than other types of definitive hosts, and that they consist mainly of species of nematodes. However, in previous studies of frogs with semi-aquatic habits, the helminthological fauna has been found to be composed mainly of digeneans. This trend has been reported several species closely associated with water: L. berlandieri (Baird, 1859) (León-Règagnon et al., 2005); L. forreri (Boulenger, 1883) (Goldberg et al., 2002); L. cf. forreri (Cabrera-Guzmán et al., 2007); L. psilonota (Webb, 2001) (Romero-Mayén et al., 2016); and L. vaillanti (Brocchi, 1877) (Paredes-Calderón et al., 2004). The dominance of digeneans is thought to reflect the close association of these species with aquatic environments. In contrast, the helminth composition reported for species that are more terrestrial (not spending as much time in water), have more species of nematodes than digeneans: such as L. brownorum (Velázquez-Urrieta & León-Règagnon, 2018; Yáñez-Arenas & Guillén-Hernández, 2010); L. tarahumarae (Boulenger, 1917) (Bursey & Goldberg, 2001); L. cf. forreri (Velázquez-Urrieta & León-Règagnon, 2018); and L. magnaocularis (Frost and Bagnara, 1974) (Goldberg & Bursey, 2002). This difference is reflected in the results of the present study; the frogs at Las Pilas are more terrestrial (more species of Nematoda) and the frogs at Metznoxtla are more aquatic (more species of Digenea) (Table 4).
Differences in the composition of species of helminths in each locality (β-diversity) is influenced by such factors as feeding habits, characteristics of the habitat, the ecological conditions in which the host develops, and the presence of potential intermediate hosts (Adán-Torres et al., 2018; Bower et al., 2018; Cabrera-Guzmán et al., 2007; Campião et al., 2012, 2014; García-Altamirano et al., 1993; Paredes-Calderón et al., 2004; Poulin, 1997; Yáñez-Arenas & Guillén-Hernández, 2010). Furthermore, these conditions may influence the survival of free-living stages of some species of helminths (Adán-Torres et al., 2018; de Sena et al., 2018; Luque et al., 2005; Muzzall, 1991; Muzzall et al., 2001). The presence or absence of particular species of helminth in each locality can be related to the type of life cycle of the parasite present (Poulin, 1997) and the presence of potential intermediate hosts (Paredes-Calderón et al., 2004). The behavior of frogs in their respective localities is also thought to affect β-diversity (Bolek & Coggins, 2003; Espínola-Novelo et al., 2017; Hamann et al., 2006a; Kehr, et al., 2006; McAlpine, 1997; Muzzall & Mychek-Londer, 2014; Romero-Mayén et al., 2016; Santos & Amato, 2010; Yáñez-Arenas & Guillén-Hernández, 2010). Local differences in the strength of these factors, and interactions between the factors present in given localities, generates dissimilarity in the structure of the communities of helminths at local levels, which, combined with local environmental factors, generate β-diversity. Thus, local conditions produce higher species richness, abundance, and prevalence of some species in one locality and, in others with different local conditions, lower values of ecological parameters occur. Comparing β-diversity is an important area of basic and applied ecological research; however, few studies have addressed β-diversity in studies of communities of parasitic organisms, especially in anurans. Studies of β-diversity are carried out more commonly with parasites of mammals and fish (Krasnov et al., 2019; Qin et al., 2019; Spickett et al., 2017; Spickett et al., 2019). The present study suggests that increased research in the understanding of the patterns of spatial variation of parasite communities at the level of β-diversity will provide information that will aid in the interpretation of comparative results.
Among the factors listed above that can influence the composition of species in a helminths community in a given species of host, host size is considered to be an important predictor of helminth community structure, and a positive correlation between host size and helminth abundance is somewhat expected (Poulin, 1997; Williams & Jones, 1994). However, in this study correlation between the abundance of helminths and the snout-vent length of frogs was found only in females from Metznoxtla, even though, as mentioned above, on the average they were significantly smaller than those in Las Pilas. A correlation only in Metznoxtla could be because of the probable habitat-related differences in the diet and behavior of the frogs in this locality; both factors are important in the recruitment of helminths (Campião et al., 2014; Hamann et al., 2006b).
To date, there are no published studies of the helminths of frogs at Las Pilas; however, Pulido-Flores et al. (2009) carried out a pilot study of 49 individuals of L. spectabilis at Metznoxtla in 2003. They reported 4 species: G. quieta, H. complexus (Seely, 1906) Krull, 1933, Gorgoderina attenuata (Stafford, 1902) Stafford, 1905, and Ochoterenella digiticauda Caballero y Caballero, 1944. The specimens reported as H. complexus were reexamined for the present study and found to be H. caballeroi. The present study also reported G. quieta and H. caballeroi but did not report G. attenuata or O. digiticauda, both of which had low prevalence and abundance in the study of (Pulido-Flores et al., 2009). Despite the greater number of frogs collected in that study compared to the present study (49 vs. 37), the species richness was much lower (4 species vs. 7 species). Two species were reported by Pulido-Flores et al. (2009) that were not reported in the present study and 5 species were reported in the present study that were not reported in the previous one; 2 species were common to both studies. The species with the highest prevalence and abundance was G. quieta in both studies, but in the previous study the values (37.9% and 7.39, respectively) were lower than in the present study (51.35% and 11.03, respectively). In contrast, the prevalence and abundance of H. caballeroi in the previous study (13.9% and 0.44, respectively) were higher than in the present study (2.70% and 0.05, respectively). However, it is interesting to note that 584 individuals of G. quieta and 35 individuals of H. caballeroi were collected by Pulido-Flores et al. (2009) and only 408 of G. quieta and 2 individuals of H. caballeroi were collected in the present study. No data was collected between the dates of the 2 studies, so we have no information on which to base an interpretation of the differences between them.
As discussed above, Hill numbers are based on a sampling theory that proposes to produce indices that can provide a basis of comparison between the ecological parameters of communities with biological and environmental conditions that always are different. The Hill numbers are indices that smoothly link rarefaction (interpolation) and prediction (extrapolation) to standardize samples based on sample size and sample completeness and to facilitate the comparison of biodiversity data. Although the concept of Hill numbers is not new (Hill, 1973), they have not been widely applied in studies of parasite communities, notorious for having unique evolutionary and biological that affect the host-parasite interactions in distinctive environments.
The only study employing Hill numbers in describing communities of helminths in Mexican anurans is that of Espínola-Novelo et al. (2017). In that study, the authors quantified the ecological parameters of the helminth communities of Rhinella marina Linnaeus, 1758 and Incilius valliceps Wiegmann, 1833 from Lagunas de Yalahau, Yucatán, Mexico. The sample size of the helminths in these amphibians and the results of the richness and diversity estimators (Hill numbers; Table 5) were sufficient to suggest that the richness of species of helminth in that locality was relatively low, and the pattern proposed for amphibians is followed; i. e., nematodes are commonly more dominant, and the richness of other species is lower (Aho, 1990).
One of the important aspects of the Hill numbers is their usefulness for making predictions about the richness and diversity of communities of similar organisms (Jost, 2006, 2007). Based on the study of the helminth community of the 2 species of toads (I. valliceps and Rhinella marina) by Espínola-Novelo et al. (2017), the anurans in a more terrestrial environment would have q values lower than those in a combination of aquatic-terrestrial environments; in that case, the toads are in the same general locality, but they have different habitat preferences. In the present study, the frogs in Las Pilas are in an aquatic-terrestrial environment and those in Metznoxtla are in an aquatic one. Thus, it would be predicted that the q values of the helminth community of frogs in the in the more complex 2 component environment (Las Pilas) would have higher richness and diversity than the frogs in the single-component environment (Metznoxtla); in the present study, the species of host is the same in both localities, but because one is a flowing river and one is a small pond, the habitat usage and behavior is different. As predicted, the 3 q values were higher for Las Pilas than for Metznoxtla (Table 5). Of course, this validity and usefulness is supported by the results of 2 studies. The predictive value of q values must be evaluated using Hill numbers in future studies.

