











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Expectations of the alleged endemism in the Revillagigedo Archipelago for certain taxa (Aguirre-Muñoz et al., 2015), and particularities of benthic diatom taxocenoses from remote locations, prompted a floristic study of epiphytic diatom assemblages generating the first species list for the region (Siqueiros-Beltrones et al., 2019). Said study yielded a species richness of epiphytic diatoms on macroalgae from the Revillagigedo Archipelago of 202 diatom taxa. However, it relied on the inspection of only 2 macroalgae specimens of Laurencia sp. (Rhodophyta). It was thus assumed that further inspections comprising a larger sample of macroalgae specimens would yield at least twice the number of diatom taxa (Siqueiros-Beltrones et al., 2019, 2021). Notwithstanding, besides continuing the floristic account that will allow for the determination of ecological relations and biogeographic affinities of benthic diatom flora in the Western Pacific, more specific inventories dealing with certain taxa, mainly those that are conspicuous, either abundant or large forms, may be useful while addressing the above topics. Species of the genus Mastogloia Thwaites comply with such requirements.
The frustules of Mastogloia are naviculoid and isopolar, characterized by the presence of partecta, i. e., loculate marginal septa in the valvocopulae that bear 1 or many chambers (Round et al., 1990). Species are ubiquitous, mainly marine forms, although some are common in brackish water and freshwater habitats. Most are benthic, abundant in epipelic and epiphytic assemblages (Round et al., 1990), particularly in tropical and subtropical latitudes where they occur in high numbers and species richness (Frankovich et al., 2006).
The genus Mastogloia is frequently the most diverse within benthic marine assemblages from warm seas (Hafner et al., 2018; Siqueiros-Beltrones et al., 2020, 2021). Currently, 271 species are accepted out of 420 taxa proposed worldwide (Guiry & Guiry, 2021). Hustedt’s (1959) work, alone, provides descriptions for 140 Mastogloia taxa from various parts of the planet. In this sense, the above exploratory study for the Revillagigedo Archipelago included 28 Mastogloia species and infraspecific taxa (Siqueiros-Beltrones et al., 2019), besides many other taxa recorded from the tropics, that support the overall tropical affinity for the benthic diatom flora of the Revillagigedo Archipelago. A similar study carried out at Isla Guadalupe off Baja California, Mexico, where transitional tropical and temperate oceanographic conditions are characteristic, yielded less (119) taxa, although with many more samples (López-Fuerte et al., 2015). There, Mastogloia species comprised the higher number of species (13); however, in the Revillagigedo Archipelago twice the number of species (28) of this genus had been hitherto recorded (Siqueiros-Beltrones et al., 2019).
Elsewhere, in Atlantic shores of America, Hein et al. (2008) recorded 75 Mastogloia taxa from the Bahamas Archipelago, while Loir and Novarino (2013) recorded a higher number of species (> 80) for several substrata in islands of the Caribbean Sea, which they described and illustrated. Also, in other floristic studies of epiphytic diatoms of Thalassia testudinum Banks ex König off the coast of Quintana Roo (Caribbean Sea), López-Fuerte et al. (2013) recorded 30 Mastogloia taxa. And more recently a total of 76 Mastogloia species epiphytic on blades of the seagrass T. testudinum were observed off the coast of Campeche in the southern Gulf of México (Siqueiros-Beltrones et al., 2020), which is the highest number of Mastogloia taxa recorded for Mexican coasts. Albeit, in that study, only 51 Mastogloia species or infra-specific taxa were formally identified, while 25 remained as cf. or sp. status.
On the other hand, for the Mexican tropical Pacific, Meave-del Castillo et al. (2003) compiled 25 Mastogloia planktonic records, while Moreno et al. (1996) accounted for 38 Mastogloia taxa in plankton samples solely for the warm waters of the Gulf of California. Thus, although the Revillagigedo Archipelago is located within a transitional zone influenced by the California Current (Aguirre-Muñoz et al., 2015), the tropical component of the archipelago region seems to be a major factor enhancing the species diversity of epiphytic diatoms (Siqueiros-Beltrones et al., 2019). In particular, the Mastogloia species that currently comprise the higher number of species recorded for the archipelago (28 taxa) were here expected to at least approximate the number of species recorded recently for the southern Gulf of Mexico (Siqueiros-Beltrones et al., 2020).
The above accounts set a reference as to the expected diversity in any given area for species of the genus Mastogloia, particularly within tropical regions. However, they also show that much floristics and formal taxonomy are required on benthic diatoms from different types of substrata for Mexican littorals (Siqueiros-Beltrones et al., 2020). It thus seemed imperative to continue surveying the area in order to gather a precise reference on the diversity of Mastogloia taxa. Our aim was to attain a closer estimate of the species richness of Mastogloia living on macroalgae from the Revillagigedo Archipelago, inasmuch distribution data based on these taxa could be used to support further hypothesis driven studies on diatom ecology and biogeography for the Western Pacific.
Materials and methods
The Revillagigedo Archipelago is set under the jurisdiction of the state of Colima, México. It comprises 4 islands, San Benedicto, Socorro, Roca Partida, and Clarion, located 386 km south of Cabo San Lucas, Baja California Sur and has been recently declared as World Heritage Site (Conanp, 2017). Seasonal variations in the region seem to be determined by the alternating influence of the California Current and the North-equatorial current, whilst the rest of the year a transition between both states may be observed (Lluch-Cota et al., 1994). A rich macroalgal community of over 190 macroalgae taxa documented in the archipelago, that harbor highly diverse assemblages of epiphytic diatoms may be found in the rocky shores of the archipelago (León-Tejera et al., 1996; Siqueiros-Beltrones et al., 2019, 2021).
During an opportunity (recreational) trip to the Revillagigedo Islands within the 2019 February 10-15 period, 15 macroalgae thalli were collected manually by Scuba at 7-27 m depth from the rocky shore of San Benedicto and Socorro islands (Revillagigedo Archipelago); specific sampling points included El Boiler (19°19’38.30” N, 110°48’17.16” W), El Cañón (19°17′40.48” N, 110°48′40.92” W) located in the former and Cabo Pearce (18°45′57.24” N, 110°54′00.30” W) and Punta Tosca (18°46′33.46” N, 111°03′26.01” W) in the latter (Fig. 1). The specimens were sun-dried, transported in plastic bags, and identified in the laboratory following Abbot and Hollenberg (1976). Diatoms were brushed off from each algae specimen while rinsing with purified water. The brushed-off material was placed in a 150 mL test tube and left to settle. Thereafter, the precipitates were collected and oxidized with a mixture of commercial alcohol and nitric acid at a ratio of 1 (sample): 2 (alcohol): 5 (acid), according to Siqueiros-Beltrones (2002). The oxidized material was rinsed repeatedly with purified water until it reached a pH ≥ 6. For each sample 2 double permanent slides were mounted using Pleurax® (RI = 1.7) (made and distributed by Bill Daily, University of Pennsylvania), which were then inspected under several magnifications (250×, 400×, 630× and 1,000×) for general recognizance of diatoms.

The Mastogloia taxa were identified at 1,000× under an Olympus CH-2 (Olympus, Japan) compound microscope using clear field illumination, following Foged (1975, 1978, 1984), Hein et al. (2008), Hustedt (1959), Lobban et al. (2012), Loir and Novarino (2013), Moreno et al. (1996), Peragallo and Peragallo (1908), Schmidt et al. (1959), Siqueiros-Beltrones et al. (2017, 2019, 2020), Stidolph et al. (2012), Witkowski et al. (2000). Taxonomic names were updated according to Guiry and Guiry (2021). An iconographic catalog of the recorded Mastogloia taxa was assembled with specimens captured with a digital ocular Microcam 1.3, at 25×, 63× and mostly at 1,000× (Olympus, Japan).
Results
The inspected macroalgae specimens from the Revillagigedo Archipelago included phaeophytes (Canistrocarpus cervicornis (J. V. Lamouroux) De Paula & De Clerck, Dictyota crenulata J. Agardh, D. dichotoma (Hudson) J. V. Lamouroux, Sargassum liebmannii J. Agardh) and rhodophytes (Amphiroa beauvoisii J. V. Lamouroux, Asparagopsis sp., and cf. Laurencia sp.). Besides, several other macroalgae specimens not identified to genus or species level which were either phaeophytes or rhodophytes.
The floristic analysis of epiphytic Mastogloia found on the macroalgae yielded 51 taxa, including 23 new records and all those accounted for in the previous (only) survey, but no new taxa. Although no quantitative account was considered, frequencies of the various diatom taxa observed in the mounted slides were clearly heterogeneous. However, specimens of Mastogloia were present in all the samples, including 1 or more conspicuous (large) forms detected firstly under low magnification (250×) and later precisely identified under 1000×. Most conspicuous forms were frequent, and several taxa seemed to dominate the assemblages, e. gr. Mastogloia binotata, M. cocconeiformis, M. cuneata, M. fimbriata, M. inaequalis, M. ovum-paschale, M. punctatissima, and others recorded earlier. New records such as M. asperuloides, M. graciloides, M. peragalloi, and M. mauritiana occurred frequently. Because certain Mastogloia taxa are morphologically very similar, their images were placed together in the iconographic catalog (Figs. 2-26) which allowed for reliable discrimination between similar taxa and precise identification.

Figure 2 (a) Mastogloia horvathiana, (b) M. punctatissima, (c) M. ovata, (d) M. pseudolatecostata, (e) M. cocconeiformis, (f) M. ovum-paschale, (g) M. ovulum, (h) M. ovalis, (i) M. splendidula, (j) M. emarginata. Scale bar = 10 μm.


Figure 4 (a-d) Mastogloia labuensis var. lanceolata, (e, f) M. affinis, (g-n) M. apiculata. Scale bar = 10 μm.


Figure 6 (a-g) Mastogloia barbadensis, (h-l) M. binotata, (m, n) M. borneensis, (o, p) M. marginulata. Scale bar = 10 μm.

Figure 7 (a-e) Mastogloia cannii, (f-i) M. elegans, (j-m) M. quinquecostata, (n) M. grunowii. Scale bar = 10 μm.

Figure 8 (a-c) Mastogloia cocconeiformis, (d-f) M. cribrosa, (g) M. fimbriata, (h-k) M. corsicana. Scale bar = 10 μm.



Figure 11 (a-f) Mastogloia cyclops, (g, h) M. biapiculata, (i-p) M. subaffirmata, (q, r) M. parva. Scale bar = 10 μm.



Figure 14 (a-p) Mastogloia exilis, (q-t) M. tenuis, (u-w) M. pusilla var. subcapitata, (x, y) M. manokwariensis. Scale bar = 10 μm.
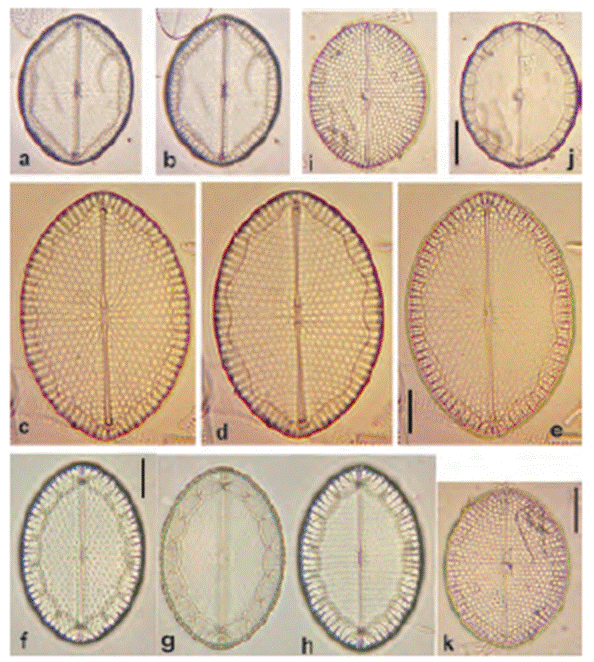



Figure 18 (a-e) Mastogloia ovalis, (f-p). M. ovulum, (q-u) M. emarginata, (v-z) M. splendidula. Scale bar = 10 μm.
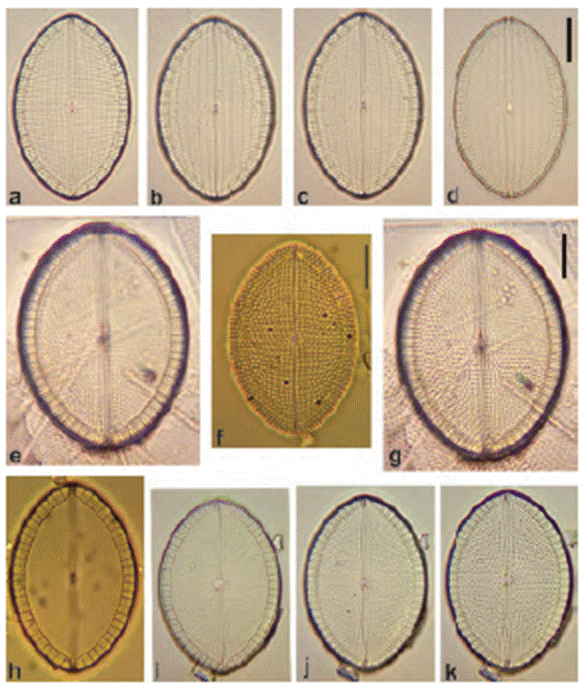



Figure 22 (a-c, e-k) Mastogloia peragalloi, (d, l) M. mauritiana, (m-p) M. japonica. Scale bar = 10 μm.


Figure 24 (a-d) Mastogloia punctatissima. Several depth perceptions are shown showing small partecta. Scale bar = 10 μm.

Figure 25 (a-d) Mastogloia pseudolatecostata. Several depth perceptions are shown where partecta are clearly defined. Scale bar = 10 μm.

Figure 26 (a-g) Mastogloia sublatericia, (h, i) M. gilberti, (j, k) M. lacrimata, (l-n) M. splendidula. Scale bar = 10 μm.
In what follows, the floristic list of epiphytic Mastogloia taxa on macroalgae thalli recorded at the Revillagigedo Archipelago is presented, along with references used to identify them and remarks. The 23 new records are marked (*) and include morphological data, while others previously recorded are referred to Siqueiros-Beltrones et al. (2019).
Bacillariophyceae E. Haeckel
Mastogloiales D. G. Mann
Mastogloiaceae C. Mereschkowsky
Mastogloia G. H. K. Thwaites
Mastogloia acutiuscula var. elliptica F. Hustedt (Fig. 3 a-l) References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: hard to distinguish from M. decipiens, but smaller, with more squared ends.
Mastogloia affinis P. T. Cleve (Fig. 4e, f).
References: Hustedt (1959); Loir and Novarino (2013).
Remarks: similar to M. varians, but with coarser striae and less partecta.
Mastogloia affirmata (G. Leudiger-Fortmore) P. T. Cleve (no image).
Reference: Hustedt (1959).
Remarks: very scarce, not found in these samples. Previously recorded in Revillagigedo Islands by Siqueiros-Beltrones et al. (2019).
Mastogloia apiculata W. Smith (Fig. 4g-n).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: a common taxon in most samples.
*Mastogloia asperuloides F. Hustedt (Fig. 5a-i)
Reference: Loir and Novarino (2013) p. 19, pl.3, figure b.
L = 48-55, W = 25-27 μm, 5 partecta/10 μm, 14 striae/10 μm.
Remarks: frequent in samples. Although it resembles a small M. horvathiana, it has square partecta, while in the latter these are narrow rectangular.
*Mastogloia barbadensis (R. K. Greville) P. T. Cleve (Fig. 6a-g).
Reference: Hustedt (1959) p. 472, figure 891; Loir and Novarino (2013) p. 20, pl. 3, figure c.
L = 35-40 μm, W = 20-27 μm, 3-3.5 partecta/10 μm, 11-12 striae/10 μm.
Remarks: few specimens observed.
*Mastogloia biapiculata F. Hustedt (Fig. 11g, h).
Reference: Hustedt (1959) p. 513, figure 943; Simonsen (1987) p. 113, pl.186, figure 8.
L = 35 μm, W = 16 μm, 6 partecta/10 μm, striae indistinguishable, with longitudinal striae.
Remarks: scarce and inconspicuous. In references images are confused with M. parva. First record for Mexican waters.
Mastogloia binotata (A. Grunow) P. T. Cleve (Fig. 6h-l). References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: a very common taxon showing much size variation.
Mastogloia borneensis F. Hustedt in A. Schmidt (Fig. 6m, n).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: common although not conspicuous because of its small size.
*Mastogloia cannii K. D. Kemp & T. B. B. Paddock (Fig. 7a-e).
Reference: Lobban et al. (2012) p. 267, pl. 24, figures 1-3.
L = 30-32 μm, W = 16-18 μm, 7 partecta/10 μm, 20 striae/10 μm.
Remarks: frequent. Morphologically similar to the larger M. elegans. First record for Mexican waters.
Mastogloia ciskeiensis M. H. Giffen (no image).
References: Foged (1975; 1978). Remarks: not found in these samples.
Mastogloia cocconeiformis A. Grunow (Figs. 2e, 8a-c). References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: a common and conspicuous (large) taxon. Mastogloia corsicana (Grunow) H. Peragallo & M. Peragallo (Figs. 8h-k and 9a-j).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: very common taxon, conspicuous in spite being relatively small.
*Mastogloia cribrosa A. Grunow (Figs. 8d-f and 15i-k) Reference: Hustedt (1959) p. 468, figure 887.
L = 32μm, W = 24μm, 3.5-4 partecta/10 μm, 12-14 striae/10 μm.
Remarks: scarce. Smaller, more rounded than M. fimbriata. Although currently regarded a synonym with Orthoneis cribrosa in the online platforms (Guiry & Guiry, 2021) M. cribrosa is still widely used in classic and recent literature. It thus awaits updating.
Mastogloia crucicula (A. Grunow) P. T. Cleve (no image).
References: Loir and Novarino (2013); Witkowski et al. (2000); Siqueiros-Beltrones et al. (2019).
Remarks: frequent and abundant; most of the time with a faint appearance. Although this name is considered invalid in online platforms, it is currently used as a reliable reference.
Mastogloia crucicula var. alternans V. Zanon (no image).
References: Loir and Novarino (2013); Witkowski et al. (2000); Siqueiros-Beltrones et al. (2019).
Remarks: frequent and most of the time with a faint appearance.
Mastogloia cuneata (F. Meister) R. Simonsen (Fig. 9p-v). References: Hustedt (1959); Loir and Novarino (2013).
Remarks: very common, although inconspicuous due to its small size.
*Mastogloia cyclops M. Voigt (Fig. 11a-f)
Reference: Lobban et al. (2012) p. 271, pl. 27, figures 4 and 5.
L = 38-40 μm, W = 19-20 μm, 8-10 partecta/10 μm, 16-17 striae/10 μm.
Remarks: frequent. Distinguished from M. subaffirmata by the stigma and smaller partecta.
*Mastogloia decipiens F. Hustedt (Fig. 3m-p) Reference: Hustedt (1959) p. 504, figure 929.
L = 28-38 μm, W = 13-17 μm, 6 partecta/10 μm, striae not visible in our specimens.
Remarks: scarce. Distinguished from M. acutiuscula var. elliptica by valve margin tapering into the capitate apices and more numerous partecta. Mastogloia delicatissima F. Hustedt (Fig. 12a-h).
References: Hustedt (1959); Witkowski et al. (2000).
Remarks: common but inconspicuous because of its small size.
*Mastogloia elegans C. Lewis (Fig. 7f-i).
Reference: Foged (1984) p. 52, pl. 35, figures 1 and 2.
L = 45-52, W = 18-21, 8 partecta/10 μm, 16-18 striae/10 μm.
Remarks: frequent. Resembles an elongated form of M. cannii. First record for Mexico.
Mastogloia emarginata F. Hustedt (Figs. 2j, 18q-u).
References: Hustedt (1959); Loir and Novarino (2013).
Remarks: common; distinguished from M. ovulum by longer partecta and longitudinal wavy lines.
Mastogloia erythraea A. Grunow (Fig. 13a-g).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: conspicuous but not common. Our specimens fit, with slight variations due to size that of Loir and Novarino’s form a.
Mastogloia exilis F. Hustedt (Fig. 14a-p).
References: Hustedt (1959); Loir and Novarino (2013).
Remarks: small and inconspicuous, frequent in some samples.
Mastogloia fimbriata (T. Brightwell) A. Grunow (Figs. 8g and 15a-h).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: very common and abundant taxon, easily distinguishable from the smaller M. cribrosa, by its more ovate valves and the varied appearance of the partecta.
*Mastogloia gilberti A. Schmidt (Figs. 12t-w and 26h, i).
Reference: Foged (1984) (as M. gracillima), p. 53, pl. 33, figure 6; Hustedt (1959) p. 479, figure 899; Peragallo and Peragallo (1897-1908) p. 31, pl. 5, figure 23.
L = 31 μm, W = 16 μm, 3.5-4 partecta/10 μm, 15-16 striae/10 μm.
Remarks: scarce. Resembles a smaller, narrower, and less capitated form of M. lacrimata.
First record for Mexican waters.
*Mastogloia graciloides F. Hustedt (Fig. 13h-t).
Reference: Hustedt (1959) p. 508, figure 934; Lobban et al. (2012) p.274, pl. 28, figures 12 and 13.
L = 28-40μm, W = 12-15 μm, 5.5-7 partecta/10 μm, 14-15 striae/10 μm.
Remarks: very frequent in samples.
Mastogloia grunowii A. Schmidt (Figs. 7n and 10a-h).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: a taxon appearing frequently, with valves like M. quinquecostata but distinguished by a very different partecta array.
Mastogloia horvathiana A. Grunow (Figs. 2a, 16a-e).
References: Hustedt (1959); Loir and Novarino (2013); Witkowski et al. (2000).
Remarks: common conspicuous taxon that may be confused with M. pseudolatecostata, but much bigger, with coarser striae, and narrower partecta.
Mastogloia inaequalis P. T. Cleve (Fig. 9k-o).
References: Hustedt (1959); Witkowski et al. (2000).
Remarks: may be confused with the lanceolate M. ciskeiensis but being wider (rhomboid).
*Mastogloia japonica P. T. Cleve (Fig. 22m-p).
Reference: Hustedt (1959) p. 565, figure 996.
L = 34μm, W = 16 μm, 5-8 partecta/10 μm, 15-16 striae/10 μm.
Remarks: only 1 specimen observed; partecta are like M. mauritiana but the arrangement is different. Striae undulate not crossed by longitudinal lines as in the latter. First record for Mexican waters.
*Mastogloia labuensis var. lanceolata F. Hustedt (Fig. 4a-d).
Reference: Hustedt (1959) p. 518, figure 950c; Loir and Novarino (2013) p. 37, pl. 15, figure a.
L = 25μm, W = 13 μm, 5 partecta/10 μm, 22 striae/10 μm.
Remarks: scarce. Very similar to M. adriatica M. Voight, as in Witkowski et al. (2000) but Hustedt’s image shows partecta not uniform and more like those in M. exilis.
*Mastogloia lacrimata M. Voigt (Figs. 12i-s and 26j, k).
Reference: Hein et al. (2008), p. 67, pl. 33, figure 7; Lobban et al. (2012), p. 276, pl. 30, figures 6-8.
L = 40-45 μm, W = 18-20 μm, 5 partecta/10 μm, 12-13 striae/10 μm.
Remarks: scarce. Similar to M. gilberti, albeit larger.
*Mastogloia lineata P. T. Cleve & E. Grove (Fig. 17a-c).
Reference: Hustedt (1959), p. 538, figure 971.
L = 65μm, W = 27 μm, 6 partecta/10 μm, 14 striae/10 μm.
Remarks: scarce. First record for Mexican waters.
*Mastogloia manokwariensis B. J. Cholnoky (Fig. 14x, y).
Reference: Lobban et al. (2012), p. 277, pl. 32, figures 1-3; Witkowski et al. (2000), p. 253, pl. 80, figure 11.
L = 22 μm, W = 10 μm, 3 partecta/10 μm, striae indistinguishable.
Remarks: scarce. Almost indistinguishable from M. pusilla var. capitata; the latter has 2 partecta/10 μm, and a wider valve, although cited descriptions say otherwise. First record for Mexican waters.
Mastogloia marginulata A. Grunow (Fig. 6o, p).
References: Peragallo and Peragallo (1897-1908); Witkowski et al. (2000).
Remarks: distinguished from M. borneensis, synonymized in Schmidt et al. (1874-1959), it is less lanceolate and fainter.
*Mastogloia mauritiana J. Brun (Figs. 17d-l and 22d, l).
Reference: Hustedt (1959), p. 563, figure 995; Lobban et al. (2012), p. 278, pl. 32, figures 4-10; Loir and Novarino(2013), p. 38, pl. 16, figure b.
L = 32-56 μm, W = 18-22 μm, 4-5 partecta/10 μm, 22-24 striae/10 μm.
Remarks: similar to M. peragalloi but this taxon has a more lemon-like appearance, while in M. mauritiana it is lanceolate, with longitudinal lines crossing the striae.
*Mastogloia ovalis A. Schmidt (Figs. 2h, 18a-e).
Reference: Hustedt (1959), p. 474, figure 893; Loir and Novarino (2013), p. 39, pl. 17, figure a.
L = 32-33 μm, W = 17-18 μm, 3-4 partecta/10 μm, 10 striae/10 μm.
Remarks: frequent in samples; several morphological variations observed.
*Mastogloia ovata A. Grunow (Figs. 2c, 19a-d).
Reference: Hustedt (1959), p. 476, figure 895; Loir and Novarino (2013), p. 40, pl. 17, figure b.
L = 42 μm, W = 27 μm, 3.5-4 partecta/10 μm, 17-18 striae/10 μm.
Remarks: frequent. May be confused with M. pseudolatecostata, but partecta are different.
Mastogloia ovulum F. Hustedt (Figs. 2g, 18f-p).
References: Hustedt (1959); Loir and Novarino (2013).
Remarks: similar to M. emarginata but partecta are squarer and more numerous.
Mastogloia ovum-paschale (A. Schmidt) D. G. Mann (Figs. 2f, 20a-g, 21a-k).
References: Hustedt (1959); Lobban et al. (2012); Loir and Novarino (2013).
Remarks: very common, showing great size variation: Its coarser striae clearly distinguish it from M. pseudolatecostata, and its partecta from M. horvathiana. Mastogloia parva F. Hustedt (Fig. 11q, r).
Reference: Hustedt (1959); Simonsen (1987).
Remarks: scarce and inconspicuous. In references images are confused with M. biapiculata.
*Mastogloia peragalloi P. T. Cleve (Fig. 22a-c, e-k).
Reference: Hustedt (1959), p. 561, figure 994; Witkowski et al. (2000), p. 257, pl. 78, figures 5 and 6.
L = 35-43 μm, W = 23-26 μm, 6-8 partecta/10 μm, 14-15 striae/10 μm.
Remarks: frequent. Distinct from M. mauritiana, evident in different sizes.
*Mastogloia pseudolatecostata T. A. Yohn & R. A. Gibson (Figs. 2d, 19e-k, 25a-d)
Reference: Hein et al. (2008), p. 70, pl. 45, figures 1 and 3; Lobban et al. (2012), p. 281, pl. 35, figures 1 and 2.
L = 43-53 μm, W = 29-39 μm, 5-5.5 partecta/10 μm, 12-14 striae/10 μm.
Remarks: frequent. Similar to M. ovata but with smaller partecta and less rounded apices.
Mastogloia punctatissima (R. K. Greville) M. Ricard (Figs. 2b, 23a-d, 24a-d).
Reference: Hein et al. (2008).
Remarks: common and conspicuous large form, mistaken for M. splendida in several references (Hein et al., 2008). Although in various literature this taxon is regarded a synonymy of Cocconeis punctatissima (Guiry & Guiry, 2021) this are monoraphid forms, whilst M. punctatissima is birhaphid a trait that can be noted by changing focus in complete frustules (epivalve and hypovalve) as in figures 23, 24, where partecta the distinctive character of Mastogloia are visible (24 c).
Mastogloia pusilla var. subcapitata F. Hustedt (Fig. 14u-w).
Reference: Hustedt (1959); Witkowski et al. (2000).
Remarks: may be confused with M. manokwariensis but having 2 partecta/10 μm and a wider valve, although other descriptions say otherwise.
Mastogloia quinquecostata A. Grunow (Figs. 7j-m and 10i-o).
References: Hustedt (1959); Witkowski et al. (2000).
Remarks: valves similar to M. grunowii, but partecta are much smaller and numerous.
*Mastogloia splendidula F. Hustedt (Figs. 2i, 18v-z, 26l-n).
Reference: Hustedt (1959), p. 468, figure 886; Simonsen (1987), p. 136, pl. 221, figures 38-40.
L = 26 μm, W = 17 μm, 1.5 partecta/10 μm, 14-16 striae/10 μm.
Remarks: scarce. May be mistaken for M. emarginata, albeit partecta are different.
Mastogloia subaffirmata F. Hustedt in A. W. F. Schmidt (Fig. 11i-p).
References: Hein et al. (2008); Hustedt (1959); Loir and Novarino (2013).
Remarks: similar to M. cyclops but lacks the stigma and has larger partecta. Although the name is regarded as invalid, Hustedt (1959) provides a good figure and a formal description.
*Mastogloia sublatericia F. Hustedt (Fig. 26a-g).
Reference: Hustedt (1959), p. 479, figure 900; Simonsen (1987), p. 137, pl. 222, figures 12-15.
L = 28-30 μm, W = 16-19 μm, 3 partecta/10 μm, 17-18 striae/10 μm.
Remarks: frequent but inconspicuous. First record for Mexican waters.
*Mastogloia tenuis F. Hustedt (Fig. 14q-t)
Reference: Hustedt (1959), p. 570, figure 1004; Loir and Novarino (2013), p. 49, pl. 23, figure c; Simonsen (1987), p. 145, pl. 234, figures 14-17.
L = 28-30 μm, W = 8-10 μm, 3-4 partecta/10 μm.
Remarks: inconspicuous (faint) but frequent.
Discussion
The 23 new records of Mastogloia added to the current floristic list for the Revillagigedo Archipelago pushes the total for this genus to 51 taxa (Siqueiros-Beltrones et al., 2019, 2021). These exhibit a wide variety of forms that range in size from just over 10 µm long and a very light structure (M. exilis, M. pusilla var. subcapitata) to over 80 µm, with a strong silicified frustule (M. horvathiana, M. ovum-paschale, M. punctatissima).
Morphological similarity between certain taxa poses difficulty in their discrimination and causes confusion, e.gr., in the previous report a specimen of M. pseudolatecostata was placed as M. horvathiana in the iconographic reference and was not hitherto noted (Siqueiros-Beltrones et al., 2019). Likewise, the references for identification of M. parva and M. biapiculata may also lead to confusion. Here they were discriminated on the basis of (Hustedt, 1959) original description, in contrast with more recent images (Simonsen, 1987). Also, specimens of M. decipiens may have been confused with M. acutiuscula var. elliptica, and vice versa. M. labuensis var. lanceolata possess a similar problem by resembling M. adriatica. On the one hand, difficulty in identifying may comprise other taxa like M. barbadensis, M. ovata, M. ovum-paschale, and M. pseudolatecostata. On the other hand, the taxonomic problems remarked in table 1 concerning old synonymies are surprising inasmuch they refer to long established common taxa. In this study, our floristic approach precludes us from further resolution and later studies are in hand.
Regarding the number of taxa, the works by Hein et al. (2008), Loir and Novarino (2013) recorded around 80 Mastogloia taxa each. However, these studies relied on much more complete samplings, whilst in this study for the Revillagigedo Archipelago, the collected material was scarce and thus the sample size much smaller, although not as small as in the previous study (Siqueiros-Beltrones et al., 2019). Elsewhere, the epilithic and epiphytic Mastogloia identified at the Adriatic Sea by Hafner et al. (2018) reached 46 taxa using 25 samples gathered for 15 months. Moreover, although the total number of Mastogloia taxa recorded until now for the Revillagigedo Archipelago is closer to the one reported (71) for the southern Gulf of Mexico (Siqueiros-Beltrones et al., 2020), the latter also included 25 unidentified taxa which indicates a higher floristic yield.
Most likely, a factor strongly influencing the Mastogloia species richness has more to do with a limitation of sampling effort. The relative remoteness of the archipelago largely precludes access to the island habitats for carrying out comprehensive samplings, that are representative of the target taxa. This, along with the restrictions for working within protected areas, explains why discrete samplings are still being considered in exploratory phases such as this one. Although no new taxa are presented here, more extensive sampling should yield higher, more representative richness of Mastogloia taxa, and of overall benthic diatom floristics. Only then the possibility that new Mastogloia taxa may be discovered in the archipelago habitats can be discarded. Also, in this way, expectations draw closer that distribution data based on these taxa be used to support hypothesis driven research on diatom ecology and biogeography for the Western Pacific.

