











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La subfamilia Dynastinae (Coleoptera: Scarabaeidae), uno de los grupos más conspicuos y atractivos de Coleoptera, incluye a los escarabajos conocidos comúnmente como rinocerontes, toritos, elefantitos, unicornios y hércules (Ratcliffe et al., 2013). Se distribuyen ampliamente en casi todas las regiones del planeta,, con excepción de los polos, y su mayor diversidad se concentra en los trópicos, especialmente en la región neotropical. En todo el mundo se conocen más de 1,600 especies incluidas en cerca de 200 géneros (Gasca-Álvarez y Amat-García, 2010; Gasca-Álvarez et al., 2018; Ratcliffe, 2003; Ratcliffe y Cave, 2006; Ratcliffe et al., 2013).
Los dinastinos adultos se distinguen por la siguiente combinación de estados de carácter (Fig. 1): mandíbulas generalmente expuestas en vista dorsal; antenas con 9 a 10 antenómeros, base del escapo antenal no visible en vista dorsal; escutelo claramente visible; base del pronoto y élitros casi del mismo tamaño; pigidio expuesto; esternitos abdominales no constreñidos en su línea media; procoxa transversa; meso y metatarsos con uñas simples, de tamaño similares; ápice de la metatibia siempre con 2 espolones; longitud corporal entre 4 a 160 mm (Endrödi, 1985; Morón, 2004; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013). La mayoría presenta dimorfismo sexual acentuado, con excepción de la tribu Phileurini y algunos Cyclocephalini y Pentodontini. Casi todas las especies pueden ser fácilmente sexadas, ya que los machos tienen el ápice del último esternito abdominal emarginado, mientras que en las hembras es redondeado (Ratcliffe, 2003; Ratcliffe et al., 2013). Además, en algunas especies los machos presentan cuernos prominentes o proyecciones cefálicas o pronotales (incluso ambas), con excepción de algunas especies como las del género Enema Hope, 1837, en que ambos sexos presentan un cuerno cefálico, aunque en los machos es más prominente que en las hembras.
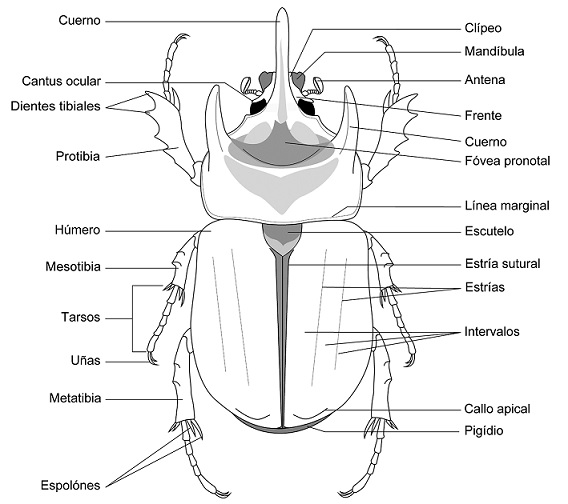
Figura 1 Vista dorsal de Strategus aloeus L. mostrando sus principales caracteres morfológicos (modificado de Ratcliffe et al., 2013).
Los adultos son principalmente de hábitos nocturnos o crepusculares y suelen ser atraídos por las luces durante la noche, esto debido a que presentan un fototropismo positivo (Gasca-Álvarez y Amat-García, 2010; Ratcliffe et al., 2013). Se alimentan de raíces, tubérculos, tallos, hojas tiernas, flores, frutos maduros o en descomposición, polen, néctar, savia, estiércol seco, madera podrida o materia orgánica humificada, incluso algunos se alimentan de otros escarabajos, por ejemplo, algunas especies de Hemiphileurus Kolbe, 1910 se han observado depredando larvas de Passalidae (Morón, 2004; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013). Las larvas son principalmente saprófagas, saproxilófagas o fitorrizófagas (Ratcliffe et al., 2013). Su ciclo de vida y estados inmaduros han sido poco estudiados, algunas especies documentadas son Ancongnatha sellata Arrow, 1911, Tomarus sallaei (Bates, 1888) (Ramírez-Salinas et al., 2004), Cyclocephala fulgurata Burmeister, 1947, C. lunulata Burmeister, 1847, C. gregaria Heney et Thaschenberg, 1907 (Bran et al., 2006), Coelosis biloba (Linnaeus, 1767) (PardoLocarno et al., 2006), Ancognatha ustulata Burmeister, 1847 (Neita-Moreno y Morón, 2008), Podischnus agenor (Olivier, 1789) (Pardo-Locarno et al., 2009), Tomarus nasutus (Burmeister, 1847) (Ramírez-Salinas et al., 2010), Dyscinetus dubius (Olivier, 1789) (Neita-Moreno y Yepes, 2011) y Phileurus valgus (Olivier, 1789) (Ibarra-Polesel et al., 2017). Se ha registrado que el ciclo completo de algunas especies, principalmente de talla grande, puede durar de 2 a 3 años. Morón (1987) señaló que el ciclo de una hembra de Dynastes hyllus Chevrolat, 1843 criada en cautiverio duró 2 años, mientras que el del macho fue de 3; resultados similares fueron registrados para Megasoma elephas (Fabricius, 1775) (Morón y Deloya, 2001) y Strategus aloeus (Linnaeus, 1758) (Ratcliffe et al., 2013).
Los dinastinos cumplen varias funciones importantes dentro de los ecosistemas, entre las principales se encuentra la aceleración en los procesos de descomposición de materia orgánica, reciclaje de nutrientes, fertilización del suelo (Morón, 1985; Morón y Aragón, 2003; Ratcliffe y Morón, 1997) y polinización (Guzmán-Vásquez et al., 2017; Moore y Jameson, 2013). Estas funciones hacen imprescindible la presencia de estos escarabajos en los ecosistemas tropicales (Iannacone-Oliver y Soras-Vega, 2010; Morón, 1985). Por lo anterior, también han sido propuestos como bioindicadores de la calidad de los ecosistemas (Otavo et al., 2013; Pardo-Locarno et al., 2011). En contraste, algunas especies de Cyclocephala Dejean, Dyscinetus Harold, Orizabus Fairmaire, Tomarus Erichson, Euetheola Bates y Strategus Kirby se incluyen en el llamado complejo “gallina ciega”, que se ha citado como el problema agrícola más frecuente en América (Morón, 2004; Morón y Aragón, 2003; Stechauner-Rohringer y Pardo-Locarno, 2010).
Actualmente, los sistemas de información geográfica (SIG) han tomado considerable relevancia en estudios de distribución y biodiversidad (IIRBA, 2006; Moreira, 1996). Esto debido a la relativa facilidad para manejar grandes cantidades de datos y obtener análisis precisos, lo cual tiene enormes ventajas sobre los métodos convencionales. Además, el uso de este tipo de herramientas permite ahorrar costos y reducir el tiempo de trabajo en campo. En ecología, los SIG se han utilizado para analizar la distribución espacial de diferentes grupos taxonómicos (por ejemplo, aves, mamíferos, arácnidos, insectos, etc.), lo cual permite inferir factores bióticos y abióticos que intervienen en el ensamblaje y agrupamiento de las especies. Además, el conocimiento de la distribución y localización de los taxones es indispensable para la conservación de especies, comunidades y ecosistemas (Moreira, 1996).
Estudios sobre distribución altitudinal se han realizado principalmente con escarabajos copronecrófagos (Scarabaeoidea), en gradientes altitudinales a escala local (Deloya et al., 2007, 2013; Halffter et al., 1995; Lobo y Halffter, 2000; Martín-Piera y Lobo, 1993). Un trabajo sobre distribución latitudinal a escala estatal se realizó con Passalinae (Passalidae) en Chiapas (Chamé-Vázquez et al., 2010). Sin embargo, con la subfamilia Dynastinae (Scarabaeidae), a la fecha no se han realizado estudios de distribución a escala estatal. Estudios similares en Oaxaca se han realizado principalmente con mamíferos (Briones-Salas et al., 2015; Martín et al., 2019; Santos-Moreno, 2014).
El estado de Oaxaca ocupa el primer lugar nacional en biodiversidad (García-Mendoza et al., 2004; Meave et al., 2012; Ordóñez y Rodríguez, 2008), a pesar de ésto, no se tiene consenso sobre la riqueza de Dynastinae (Guzmán-Vásquez, 2018). Los esfuerzos por enlistar las especies de esta subfamilia se centran en Ratcliffe y Morón (1997), quienes citan 174 especies para México y 47 para Oaxaca, posteriormente Ratcliffe et al. (2013) refieren 183 especies para el país y 89 para el estado. Por su parte, Morón et al. (2014) citan 185 especies para México y 48 para Oaxaca. Se han realizado solo 2 inventarios de faunas locales de Scarabaeoidea: el de Ramírez-Ponce et al. (2009) en Santiago Xiacui, Sierra Norte, y el de Guzmán-Vásquez (2016) en Villa de Zaachila y Santa Inés del Monte (Tabla 1). Sin embargo, debido a la gran extensión territorial del estado, al difícil acceso a muchas zonas aisladas y a la alta heterogeneidad ambiental en Oaxaca, estos esfuerzos aún son insuficientes, por lo cual, el conocimiento sobre la diversidad de Dynastinae en el estado continua escaso y desactualizado. El presente estudio contribuye con uno de los primeros esfuerzos para integrar el inventario de una de las subfamilias más conspicuas, interesantes e importantes de escarabajos.
Tabla 1 Dynastinae registrados en estudios de faunas locales de Scarabaeoidea en Oaxaca, México.
| Tribus | Géneros | Especies | Ramírez-Ponce et al. (2009) | Guzmán-Vásquez (2016) |
| Cyclocephalini | Ancognatha | A. falsa | x | |
| A. quadripunctata | x | x | ||
| Cyclocephala | C. lunulata | x | ||
| Pentodontini | Orizabus | O. clunalis | x | x |
| O. aff. batesi | x | |||
| O. sp. 1 | x | |||
| Tomarus | T. sallaei | x | x | |
| Oryctini | Xyloryctes | X. telephus | x | x |
| X. testhalus | x | |||
| Heterogomphus | H. chevrolati | x | x | |
| Strategus | S. aloeus | x | ||
| Dynastini | Dynastes | D. hyllus | x | x |
| Golofa | G. pizarro | x | x | |
| Totales | 9 gen. | 13 spp. | 9 spp. | 11 spp. |
Los objetivos de este estudio son: 1) aportar una lista actualizada de especies, obtenida de la revisión de especímenes provenientes de colecciones y recolectas en diferentes localidades, así como de la revisión de literatura, y 2) analizar la distribución espacial de las especies de Dynastinae mediante estratos o pisos altitudinales y subprovincias fisiográficas.
Materiales y métodos
El estado de Oaxaca se ubica al sureste de México (18°40’11” y 15°39’26” N; 93°52’03” y 98°33’10” O) (Fig. 2). Tiene una superficie territorial aproximada de 95,364 km2, equivalente a 4.8% de la superficie total del país (García-Mendoza et al., 2004; INEGI, 2016). Por sus características económicas, geográficas y culturales, se divide en 8 regiones: Cañada, Costa, Istmo, Mixteca, Papaloapan, Sierra Norte, Sierra Sur y Valles Centrales (Fig. 2). Éstas, a su vez, se dividen en 30 distritos, 570 municipios y más de 10,000 localidades (García-Mendoza et al., 2004). Por su rasgos geomorfológicos estructurales propios (orográficos, topográficos, geológicos e hidrográficos), se distinguen 12 regiones naturales o subprovincias fisiográficas (Ortiz-Pérez et al., 2004): Depresión del Balsas (DB), Depresión Ístmica de Tehuantepec (DIT), Fosa de Tehuacán (FT), Montañas y Valles del Centro (MVC), Montañas y Valles del Occidente (MVO), Planicie Costera de Tehuantepec (PCT), Planicie Costera del Golfo (PCG), Planicie Costera del Pacífico (PCP), Sierra Madre de Chiapas (SMC), Sierra Madre de Oaxaca (SMO), Sierra Madre del Sur (SMS) y Valles Centrales de Oaxaca (VCO).
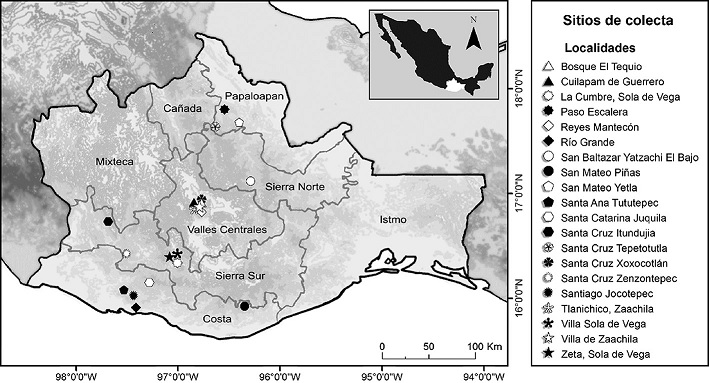
Figura 2 Localización de los sitios de colecta de escarabajos Dynastinae en Oaxaca, México y regiones geoeconómicas.
Se revisó material depositado en las siguientes colecciones locales: Instituto Tecnológico del Valle de Oaxaca (ITVO), Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Oaxaca del Instituto Politécnico Nacional (CIIDIR-OAX); colección particular Julián Hernández Cruz (CJHC); y colección particular Héctor Guzmán Vásquez (CHGV). No se revisaron colecciones nacionales (por ejemplo, UNAM, INECOL) debido a que ya han sido revisadas, la información se encuentra en Ratcliffe et al. (2013).
Durante 19 meses (septiembre 2016-abril 2018) se realizaron colectas en 20 localidades de Oaxaca (Tabla 2; Fig. 2). Se utilizó una trampa de luz tipo pantalla (lámpara con 2 tubos de luz negra (UV) de 20 watts cada uno, conectados a una fuente de energía). La trampa estuvo activa durante 12 h (19:00-7:00), durante 1-2 noches del periodo de luna nueva en una localidad distinta en cada evento de colecta. Adicionalmente, se realizaron algunas colectas manuales cerca del alumbrado público. El material recolectado está depositado en la colección CIIDIR-OAX.
Tabla 2 Localidades estudiadas (septiembre de 2016 - abril de 2018) en el estado de Oaxaca, México. Expresado por regiones geoeconómicas, tipo de vegetación, altitud, coordenadas y tipo de colecta. BP (bosque de Pinus spp.), BTP (bosque tropical perennifolio), AU (área urbana), BTC (bosque tropical caducifolio), BTsC (bosque tropical subcaducifolio), P (pastizal), MX (matorral xerófilo); metros sobre el nivel del mar (m snm); latitud norte (N), longitud oeste (O); trampa de luz (TL), colecta manual (CM), carpotrampa (CT).
| Localidad | Región geoeconómica |
Región natural |
Vegetación | Altitud (m snm) |
Coordenadas | Método de colecta |
| Cuilapam de Guerrero | Valles Centrales | VCO | BTC | 1,553 | 16°58’20” N, 96°46’48” O | CM |
| Bosque El Tequio | Valles Centrales | VCO | BTC | 1,528 | 17°00’47” N, 96°43’04” O | TL |
| La Cumbre, Sola de Vega | Sierra Sur | MVO | BP | 2,175 | 16°27’14” N, 97°00’12” O | TL |
| Paso Escalera, Usila | Papaloapan | PCG | BTP | 100 | 17°55’11” N, 96°29’04” O | TL |
| Reyes Mantecón | Valles Centrales | VCO | AU | 1,524 | 16°54’24” N, 96°43’26” O | CM |
| Río Grande | Costa | PCP | BTC | 28 | 16°00’49” N, 97°26’30” O | TL, CM |
| San Baltazar Yatzachi | Sierra Norte | SMO | BTC, BP | 1,575 | 17°13’17” N, 96°13’05” O | TL, CM |
| San Mateo Piñas | Costa | SMS | BTsC | 803 | 15°59’18” N, 96°19’34” O | TL, CM, |
| San Mateo Yetla | Papaloapan | PCG | BTP | 117 | 17°45’25” N, 96°19’03” O | TL, CM |
| Santa Ana Tututepec | Costa | SMS | BTsC | 442 | 16°10’53” N, 97°31’50” O | TL |
| Santa Catarina Juquila | Costa | SMS | BP | 1,442 | 16°14’15” N, 97°17’35” O | CM |
| Santa Cruz Itundujia | Sierra Sur | MVO | BP | 2,205 | 16°52’44” N, 97°38’13” O | TL, CM |
| Santa Cruz Tepetotutla | Papaloapan | SMO | BTP | 1,157 | 17°44’21” N, 96°33’29” O | CM, TL |
| Santa Cruz Xoxocotlán | Valles Centrales | VCO | BTC, AU | 1,532 | 17°01’44” N, 96°43’19” O | CM, TL |
| Santa Cruz Zenzontepec | Sierra Sur | MVO | BTC | 974 | 16°32’00” N, 97°29’43” O | TL |
| Santiago Jocotepec | Costa | SMS | BTC | 620 | 16°07’38” N, 97°26’17” O | TL |
| Tlanichico Zaachila | Valles Centrales | VCO | P, MX | 1,589 | 16°56’36” N, 96°48’08” O | CM, CT |
| Villa de Zaachila | Valles Centrales | VCO | AU | 1,521 | 16°57’26” N, 96°44’56” O | TL, CM |
| Villa Sola de Vega | Sierra Sur | MVO | BTC | 1,500 | 16°31’00” N, 96°59’04” O | TL |
| Zeta, Sola de Vega | Sierra Sur | MVO | BTsC | 1,599 | 16°29’13” N, 97°03’47” O | TL |
Para la determinación taxonómica se emplearon las claves de Endrödi (1985), Delgado et al. (2000) y Ratcliffe et al. (2013). Las observaciones se realizaron con los estereomicroscopios Zeiss Stemi 508 y Velab VE S-1. La lista de especies se elaboró a partir de la revisión de especímenes y de literatura. La búsqueda de literatura se realizó en las bases de datos SciELO, Redalyc, BIOONE, SpringerLink, Biodiversity Heritage Library, ScienceDirect y en los buscadores Google y Google Scholar. literatura valiosa también fue proporcionada por Héctor Gasca Álvarez y Brett Ratcliffe.
Las imágenes se tomaron con una cámara Canon EOS Rebel T6 adaptada a un microscopio Stemi 508 y con cámara digital Apple iOS 12.3.1 y se editaron con GIMP 2.10.18. (Creative Commons Attribution-ShareAlike 4.0). Los mapas se elaboraron con QGIS 3.10 (Creative Commons Attribution-ShareAlike 3.0). Las coordenadas geográficas se obtuvieron con Google Earth Pro 7.1. Los datos de altitud se obtuvieron con un modelo digital de elevación (MDE, www.inegi.org.mx). Los tipos de vegetación en los que se distribuyen las tribus se obtuvieron de capas shapefile descargadas de la Conabio, se trabajaron en QGIS 3.10 y se utilizó la clasificación de Rzedowski (1978).
Los registros taxonómicos se ordenaron de menor a mayor altitud y se clasificaron dentro de 7 estratos o pisos altitudinales (EA). Cada estrato representó un intervalo de 500 m de forma consecutiva. Posteriormente, se elaboraron las matrices de datos en Excel; para ésto, se siguió la metodología utilizada por Soberón et al. (2005), donde se colocan valores de 1 cuando hay presencia y valores de 0 para indicar ausencia. Las matrices se realizaron con base en los datos de distribución de las especies reales u observadas (DistObs), a estas se le agregaron valores potenciales de 1 en los vacíos entre el registro de menor al de mayor altitud en cada estrato, obteniendo así la matriz con la distribución potencial (DistPot).
Para analizar los datos se utilizaron índices de diversidad no paramétricos, que se basan en datos cualitativos (presencia-ausencia). Los índices más confiables o robustos para este caso son la riqueza específica como medida de diversidad alfa, el índice de similitud de Jaccard para la diversidad beta y el número total de especies presentes en el estado como medida de diversidad gamma (Moreno, 2001; Villarreal et al., 2004). Se utilizó el programa estadístico PAST 3.18 (Hammer et al., 2001) y el algoritmo UPGMA para estimar el índice de Jaccard, en donde se obtuvieron 2 dendrogramas de similitud. El primer dendrograma mostró la similitud entre los estratos altitudinales para las especies observadas y el segundo la similitud entre estratos altitudinales para la distribución potencial de las especies observadas. Para analizar los registros por subprovincias fisiográficas se realizó una matriz en Excel con valores de presencia-ausencia en cada subprovincia, esta matriz se corrió en PAST y se obtuvo el dendrograma de similitud de Jaccard.
Resultados
Se obtuvo una lista de 98 especies incluidas en 22 géneros y 6 tribus (Tabla 3). Se revisaron un total de 4,223 especímenes, 2,405 procedentes de colectas y 1,818 de colecciones. Se analizaron un total de 4,753 registros (530 de ellos son referencia de literatura) de 182 localidades (Fig. 3). De las 98 especies citadas para Oaxaca en este trabajo, se obtuvieron registros precisos para 97 de ellas. Solo de Podischnus agenor (Olivier, 1789) no se encontró algún registro preciso, por lo que es necesario confirmar su presencia en el estado. Ratcliffe et al. (2013) la mencionan para Oaxaca, pero sin datos de localidad. A las 89 especies referenciadas por Ratcliffe et al. (2013) se agregaron 9 especies, de éstas, 4 son nuevos registros publicados recientemente: Dynastes maya Hardy, 2003 (Moctezuma y Sánchez-Huerta, 2018), Tomarus selanderi (Cartwright, 1959) (López-García y Deloya, 2018), Cyclocephala alexi Ratcliffe y Delgado, 1990 y Stenocrates laevicollis Kirsch, 1871 (Delgado y Mora-Aguilar, 2020). Dos son nuevos registros para el estado (Golofa pusilla Arrow, 1911 y Cyclocephala comata Bates, 1888) y 3 son posiblemente nuevas especies ya que no se encontraron afines a ninguna especie conocida (Cyclocephala sp. 1, Cyclocephala sp. 2 y Orizabus sp. 1). Se reporta por primera vez la presencia de Golofa pusilla Arrow, 1911; se revisaron 173 especímenes provenientes de Putla Villa de Guerrero, Oaxaca. En la literatura no se encontraron localidades precisas para Cyclocephala comata Bates, 1888 en Oaxaca, aquí se proporcionan los primeros registros precisos; se revisaron 22 especímenes procedentes de San Lorenzo Cacaotepec (10), San Pablo Etla (2), San Andrés Huayapam (5), Santa Cruz Xoxocotlán (3), Villa de Zaachila (1) y Villa Sola de Vega (1).
Tabla 3 Lista de especies de Dynastinae con presencia en Oaxaca, México. Datos de distribución obtenidos de literatura con datos presentados en este estudio. *Especies endémicas de Oaxaca. **Especies endémicas de México.
| Especies | Distribución |
| Cyclocephalini Laporte 1840 | |
| Ancognatha Erichson, 1847 | |
| Ancognatha falsa Arrow, 1911** Elevación: 960-2,576 m | México: Chiapas, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Oaxaca, Puebla, Veracruz (Ratcliffe y Morón, 1997; Morón et al., 2003; Deloya et al., 2016; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Ancognatha quadripunctata Bates, 1888 Elevación: 1,200-2,100 m | México: Chihuahua, Colima, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Sinaloa, Sonora, Veracruz (Bates, 1888; Ratcliffe y Morón, 1997; Morón et al., 2003; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; Moore et al., 2018; presente estudio). |
| Ancognatha sellata Arrow, 1911 Elevación: 1,500-2,450 Aspidolea Bates, 1888 | México: Chiapas, Oaxaca (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Aspidolea fuliginea Burmeister, 1847 Elevación: 0-1,750 m | México: Chiapas, Guerrero, Hidalgo, Jalisco, Morelos, Oaxaca, Puebla, Tabasco, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; Moore et al., 2018; presente estudio). |
| Aspidolea singularis Bates, 1888 Elevación: 0-1,600 m | México: Chiapas, Oaxaca, Puebla, Tabasco, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala Dejean, 1821 | |
| Cyclocephala aequatoria Endrödi, 1963 Elevación: 10-1,500 m | México: Chiapas, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Veracruz (Endrödi, 1966, 1985; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala alexi Ratcliffe and Delgado, 1990 Elevación: 1,000-2,200 m | México: Chiapas, Oaxaca (Ratcliffe et al., 2013; Delgado y Mora-Aguilar, 2020). |
| Cyclocephala aulustjaorum Hielkema, 2017 Elevación: 0-1,900 m | México: Chiapas, Jalisco, Michoacán, Oaxaca, Veracruz (Endrödi, 1966, 1985; Morón et al., 2003; Ratcliffe et al. 2013; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala capitata Höhne, 1923** Elevación: 10-900 m | México: Chiapas, Colima, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Sinaloa (Endrödi 1966, 1985, Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala cerea Burmeister, 1847 Elevación: 83-2,300 m | México: Chiapas, Colima, Durango, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sinaloa, Tamaulipas, Veracruz (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003, Ratcliffe et al., 2013; Deloya et al., 2014; Ratcliffe y Cave, 2015; Moore et al., 2018; presente estudio). |
| Cyclocephala comata Bates, 1888** Elevación: 1,250-1,800 m | México: Durango, Edo. de México, Guanajuato, Jalisco, Michoacán, Oaxaca, San Luis Potosí, Tamaulipas (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013; Deloya et al., 2016; Moore et al., 2018; presente estudio). Nuevo registro estatal. |
| Cyclocephala complanata Burmeister, 1847 Elevación: 0-1,900 m | México: Chiapas, Morelos, Oaxaca, Quintana Roo, Tabasco, Veracruz, Yucatán (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013, Moore et al., 2018). |
| Cyclocephala concolor Burmeister, 1847 Elevación: 1,000-2,000 m | México: Chiapas, Oaxaca (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala confusa Endrödi, 1966 Elevación: 0-1,650 m | México: Chiapas, Oaxaca (Endrödi, 1966, 1985; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala curta Bates, 1888 Elevación: 100-1,200 m | México: Chiapas, Colima, Guerrero, Michoacán, Nayarit, Oaxaca, Sinaloa, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013, Moore et al., 2018; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala deceptor (Casey, 1915) Elevación: 0-2,300 m | México: Aguascalientes, Chiapas, Colima, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Sinaloa, Sonora, Tamaulipas, Veracruz (Endrödi, 1966, 1985; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018; presente estudio). |
| Cyclocephala discolor (Herbst, 1790) Elevación: 0-1,200 m | México: Chiapas, Colima, Jalisco, Michoacán, Nayarit, Oaxaca, San Luis Potosí (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013; Deloya et al., 2016; Moore et al., 2018). |
| Cyclocephala forcipulata Howden y Endrödi, 1966 ** Elevación: 650-1,650 m | México: Durango, Guerrero, Jalisco, Nayarit, Oaxaca, Sinaloa (Howden y Endrödi, 1966; Endrödi, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala freudei Endrödi, 1963 Elevación: 30-1,500 m | México: Chiapas, Chihuahua, Colima, Durango, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Puebla, Querétaro, Sinaloa, Sonora, Veracruz (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala fulgurata Burmeister, 1847 Elevación: 0-1,650 m | México: Chiapas, Ciudad de México, Hidalgo, Jalisco, Nayarit, Oaxaca, Puebla, Tamaulipas, Veracruz, Zacatecas (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala gravis Bates, 1888 Elevación: 0-1,800 m | México: Campeche, Chiapas, Guanajuato, Oaxaca, Quintana Roo, San Luis Potosí, Tabasco, Veracruz, Yucatán (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala guttata Bates, 1888 Elevación: 0-1,400 m | México: Chiapas, Morelos, Oaxaca, Puebla, San Luis Potosí, Tabasco, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala jalapensis Casey, 1915** Elevación: 1,369-2,040 m | México: Chiapas, Hidalgo, Oaxaca, Puebla, Querétaro, Veracruz (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala laminata Burmeister, 1847 Elevación: 0-945 m | México: Chiapas, Durango, Oaxaca (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala larssoni Endrödi, 1964 Elevación: 0-700 m | México: Guerrero, Jalisco, Nayarit, Oaxaca, Sonora (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; Moore et al., 2018). |
| Cyclocephala lunulata Burmeister, 1847 Elevación: 0-2,200 m | México: Aguascalientes, Baja California Sur, Campeche, Chiapas, Coahuila, Colima, Ciudad de México, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Querétaro, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán, Zacatecas (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; Moore et al., 2018; presente estudio). |
| Cyclocephala mafaffa Burmeister, 1847 Elevación: 0-2,170 m | México: Aguascalientes, Chiapas, Colima, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, San Luis Potosí, Sinaloa, Sonora, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala melanocephala (Fabricius, 1775) Elevación: 0-2,300 m | México: Aguascalientes, Baja California, Baja California Sur, Chiapas, Chihuahua, Colima, Ciudad de México, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán, Zacatecas (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018; presente estudio). |
| Cyclocephala mesophylla Mora-Aguilar y Delgado, 2012 Elevación: 600-1,750 m | México: Chiapas, Oaxaca, Veracruz (Mora-Aguilar y Delgado, 2012; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala multiplex Casey, 1915 Elevación: 0-1,800 m | México: Chiapas, Guerrero, Hidalgo, Oaxaca, Puebla, Quintana Roo, Tabasco, Veracruz (Endrödi, 1966, 1985; Ratcliffe et al., 2013, Moore et al., 2018; presente estudio). |
| Cyclocephala ovulum Bates, 1888 Elevación: 0-1,600 m | México: Chiapas, Guerrero, Jalisco, Morelos, Nayarit, Oaxaca, Puebla, Sinaloa, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe et al., 2013, Moore et al., 2018). |
| Cyclocephala picta Burmeister, 1847 Elevación: 900-2,000 m | México: Chiapas, Hidalgo, Jalisco, Michoacán, Oaxaca, Puebla, Veracruz, Yucatán (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2016; Moore et al., 2018; presente estudio). |
| Cyclocephala sanguinicollis Burmeister, 1847 Elevación: 40-2,100 m | México: Chiapas, Hidalgo, Nayarit, Oaxaca, Puebla, San Luis Potosí, Tabasco Veracruz, Yucatán (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Cyclocephala sexpunctata Laporte, 1840 Elevación: 0-2,000 m | México: Chiapas, Colima, Edo. de México, Guerrero, Hidalgo, Jalisco, Oaxaca, Puebla, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; Moore et al., 2018; presente estudio). |
| Cyclocephala sparsa Arrow, 1902 Elevación: 0-1,900 m | México: Campeche, Chiapas, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Quintana Roo, Veracruz, Yucatán (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018). |
| Cyclocephala stictica Burmeister, 1847 Elevación: 0-1,650 m | México: Chiapas, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, San Luis Potosí, Tabasco, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; Moore et al., 2018; presente estudio). |
| Cyclocephala weidneri Endrödi, 1964 Elevación: 70-2,200 m | México: Chiapas, Hidalgo, Oaxaca, Puebla, Veracruz (Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018; presente estudio). |
| Cyclocephala sp. 1 Elevación: 1,500 m | México: Oaxaca (presente estudio). |
| Cyclocephala sp. 2 Elevación: 440 m Dyscinetus Harold, 1869 | México: Oaxaca (presente estudio). |
| Dyscinetus dubius (Olivier, 1789) Elevación: 0-1,900 m | México: Campeche, Chiapas, Ciudad de México, Guerrero, Jalisco, Nayarit, Oaxaca, Quintana Roo, San Luis Potosí, Sinaloa, Tabasco, Tamaulipas, Tlaxcala, Veracruz (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; Moore et al., 2018). |
| Dyscinetus laevipunctatus Bates, 1888 Elevación: 0-1,500 m | México: Campeche, Chiapas, Ciudad de México, Coahuila, Colima, Guerrero, Jalisco, Michoacán, Nayarit, Nuevo León, Oaxaca, Puebla, Quintana Roo, San Luis Potosí, Sinaloa, Tabasco, Tamaulipas, Veracruz, Yucatán (Bates, 1888; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Moore et al., 2018; presente estudio). |
| Stenocrates Burmeister, 1847 | |
| Stenocrates bicarinatus Robinson, 1947 Elevación: 0-1,500 m | México: Campeche, Chiapas, Guerrero, Jalisco, Oaxaca, Puebla, San Luis Potosí, Tabasco, Veracruz (Robinson, 1947; Endrödi, 1966, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013; Moore et al., 2018). |
| Stenocrates canuli Delgado, 1991 Elevación: 0-1,000 m | México: Campeche, Chiapas, Oaxaca, Quintana Roo, Veracruz, Yucatán (Delgado, 1991; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Stenocrates duplicatus Endrödi, 1967 Elevación: 0-180 m | México: Chiapas, Jalisco, Oaxaca, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Moore et al., 2018). |
| Stenocrates laevicollis Kirsch, 1871 Elevación: 0-1,200 m Pentodontini Mulsant, 1842 Bothynus Hope, 1837 | México: Chiapas, Hidalgo, Oaxaca, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al, 2003; Ratcliffe et al., 2013; Delgado y Mora-Aguilar, 2020). |
| Bothynus complanus (Burmeister, 1847) Elevación: 0-2,600 m Euetheola Bates, 1888 | México: Chiapas, Colima, Guerrero, Jalisco, Michoacán, Oaxaca, Tabasco, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Euetheola bidentata (Burmeister, 1847) Elevación: 0-3,430 m Orizabus Fairmaire, 1878 | México: Chiapas, Hidalgo, Morelos, Oaxaca, Tamaulipas, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Orizabus batesi Prell, 1914** Elevación: 1,550-1,850 m | México: Chiapas, Chihuahua, Durango, Edo. de México, Morelos, Nayarit, Oaxaca, Sinaloa (Endrödi, 1985; Morón et al., 2003; Ratcliffe y Morón, 1997; Delgado, 2008; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013). |
| Orizabus clunalis (LeConte, 1856) Elevación: 450-3,000 m | México: Chiapas, Chihuahua, Coahuila, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nuevo León, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sonora, Tamaulipas, Tlaxcala, Veracruz, Zacatecas (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ordóñez y Deloya, 2004; Delgado 2008; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al, 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Orizabus delgadoi Ratcliffe and Cave, 2010* Elevación: 3,000 m | México: Oaxaca (Ratcliffe y Cave, 2010; Ratcliffe et al., 2013). |
| Orizabus fairmairei (Bates, 1888)** Elevación: 1,900-2,650 m | México: Coahuila, Ciudad de México, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nuevo León, Oaxaca, Puebla, San Luis Potosí, Tamaulipas, Veracruz, Zacatecas (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Delgado, 2008; Ratcliffe et al., 2013; presente estudio). |
| Orizabus isodonoides Fairmaire, 1878** Elevación: 1,200-2,600 m | México: Chihuahua, Ciudad de México, Durango, Edo. de México, Hidalgo, Jalisco, Michoacán, Morelos, Oaxaca, Puebla, San Luis Potosí, Sonora, Veracruz, Zacatecas (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Delgado, 2008; Ratcliffe et al., 2013; presente estudio). |
| Orizabus rawlinsi Dechambre, 1993* Elevación: 1,450-1,650 m | México: Oaxaca (Dechambre, 1993; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Orizabus subaziro Ratcliffe, 1994* Elevación: 400-1,650 m | México: Oaxaca (Ratcliffe, 1994; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Orizabus sp. 1 Elevación: 2,246 m Tomarus Erichson, 1847 | México: Oaxaca (presente estudio). |
| Tomarus bituberculatus: Palisot de Beauvois, 1811) Elevación: 0-2,800 m | México: Chiapas, Oaxaca, Tabasco, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Tomarus cicatricosus (Prell, 1937) Elevación: 0-1,600 m | México: Chiapas, Oaxaca, Tabasco, Veracruz, Yucatán (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Tomarus ebenus (De Geer, 1774) Elevación: 0-2,100 m | México: Chiapas, Ciudad de México, Nayarit, Oaxaca, Tabasco, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Tomarus laevicollis (Bates, 1888) Elevación: 0-2,100 m | México: Baja California Sur, Chiapas, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Quintana Roo, San Luis Potosí, Sinaloa, Tabasco, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Tomarus nasutus (Burmeister, 1847) Elevación: 0-1,800 m | México: Campeche, Chiapas, Colima, Guerrero, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Quintana Roo, San Luis Potosí, Tamaulipas, Veracruz, Yucatán (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; López-García et al., 2016; presente estudio). |
| Tomarus sallaei (Bates, 1888) Elevación: 0-2,500 m | México: Aguascalientes, Chiapas, Chihuahua, Coahuila, Colima, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán, Zacatecas (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Tomarus selanderi (Cartwright, 1959)** Elevación: 30-1,500 m | México: Colima, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Sinaloa, Sonora (Endrödi, 1985; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; López-García y Deloya, 2018). |
| Oryctini Mulsant, 1842 | |
| Coelosis Hope, 1837 | |
| Coelosis biloba (Linnaeus, 1767) Elevación: 0-2,600 m Enema Hope, 1837 | México: Campeche, Chiapas, Ciudad de México, Oaxaca, Quintana Roo, Tabasco, Veracruz, Yucatán (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Enema endymion Chevrolat, 1843 Elevación: 0-1,800 m | México: Campeche, Chiapas, Hidalgo, Oaxaca, Puebla, Quintana Roo, San Luis Potosí, Tabasco, Tamaulipas y Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; presente estudio). |
| Enema pan (Fabricius, 1775) Elevación: 0-2,800 m Heterogomphus Burmeister, 1847 | México: Chiapas, Hidalgo, Nayarit, Oaxaca, Puebla, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Heterogomphus chevrolati Burmeister, 1847 Elevación: 0-3,020 m | México: Chiapas, Edo. de México, Guerrero, Hidalgo, Morelos, Oaxaca, Puebla, San Luis Potosí y Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al., 2014; Guzmán-Vásquez, 2016; presente estudio). |
| Heterogomphus flohri (Kolbe, 1906) Elevación: 0-3,500 m | México: Chiapas, Edo. de México, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Podischnus Burmeister, 1847 | |
| Podischnus agenor (Olivier, 1789) Elevación: 0-1,500 m | México: Chiapas, Jalisco, Oaxaca, Tabasco, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al, 2003; Ratcliffe et al., 2013). |
| Strategus Kirby, 1828 | |
| Strategus aloeus (Linnaeus, 1758) Elevación: 0-2,800 m | México: Aguascalientes, Campeche, Chiapas, Chihuahua, Coahuila, Colima, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán, Zacatecas (Ratcliffe, 1976; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Strategus hipposiderus Ratcliffe, 1976 Elevación: 0-1,500 m | México: Chiapas, Oaxaca, Veracruz (Ratcliffe 1976; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Strategus jugurtha Burmeister, 1847 Elevación: 0-2,600 m | México: Chiapas, Chihuahua, Jalisco, Michoacán, Oaxaca, Sinaloa, Veracruz (Ratcliffe, 1976; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Strategus longichomperus Ratcliffe, 1976 Elevación: 0-1,000 m Xyloryctes Hope, 1837 | México: Chiapas, Oaxaca, Tamaulipas, Veracruz (Ratcliffe, 1976; Endrödi, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013). |
| Xyloryctes corniger Bates, 1888 Elevación: 960-2,680 m | México: Chiapas, Coahuila, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Querétaro, San Luis Potosí, Sinaloa, Sonora, Veracruz, Zacatecas (Bates, 1888; Endrödi, 1976, 1985; Delgado y Nájera-Rincón, 1992; Ratcliffe y Morón, 1997; Morón et al., 2003; Lugo-García et al., 2013; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016). |
| Xyloryctes ensifer Bates, 1888 Elevación: 1,540-3,089 m | México: Chiapas, Chihuahua, Coahuila, Ciudad de México, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Tlaxcala, Veracruz, Zacatecas (Bates, 1888; Endrödi, 1976, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016; presente estudio). |
| Xyloryctes furcatus Burmeister, 1847** Elevación: 0-2,860 m | México: Chiapas, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Oaxaca, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016). |
| Xyloryctes lobicollis Bates, 1888 Elevación: 0-2,500 m | México: Chiapas, Ciudad de México, Hidalgo, Jalisco, Michoacán, Oaxaca, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016). |
| Xyloryctes telephus Burmeister, 1847 Elevación: 0-3,400 m | México: Ciudad de México, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Oaxaca, Puebla, Veracruz (Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Xyloryctes teuthras Bates, 1888 Elevación: 580-2,400 m | México: Chiapas, Guerrero, Hidalgo, Jalisco, Michoacán, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sinaloa, Sonora, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2016; presente estudio). |
| Xyloryctes thestalus Bates, 1888 Elevación: 0-2,900 m | México: Aguascalientes, Chiapas, Chihuahua, Coahuila, Ciudad de México, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Querétaro, San Luis Potosí, Sinaloa, Sonora, Tamaulipas, Veracruz, Zacatecas (Bates, 1888; Endrödi, 1985; Delgado y Nájera-Rincón, 1992; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Bitar y Morón, 2014; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Phileurini Burmeister, 1847 | |
| Hemiphileurus Kolbe, 1910 | |
| Hemiphileurus dejeani (Bates, 1888) Elevación: 0-2,164 m | México: Chiapas, Hidalgo, Oaxaca, Puebla, Veracruz (Bates, 1888; Endrödi, 1978; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Hemiphileurus flohri (Kolbe, 1910)** Elevación: 1,000-1,550 m | México: Guerrero, Jalisco, Oaxaca, Veracruz (Endrödi, 1978; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; presente estudio). |
| Hemiphileurus laevicauda (Bates, 1888) Elevación: 60-1,500 m | México: Chiapas, Guerrero, Morelos, Oaxaca (Bates, 1888; Endrödi, 1978; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014). |
| Hemiphileurus microps (Burmeister, 1847) Elevación: 80-1,000 m Homophileurus Kolbe, 1910 | México: Chiapas, Hidalgo, Nuevo León, Oaxaca, Puebla, San Luis Potosí, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1978, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Homophileurus tricuspis Prell, 1914 Elevación: 0-1,800 m Phileurus Latreille, 1807 | México: Campeche, Chiapas, Hidalgo, Oaxaca, Puebla, Quintana Roo, San Luis Potosí, Tabasco, Veracruz, Yucatán (Endrödi, 1978, 1985; Ratcliffe y Morón, 1997; Ratcliffe et al., 2013). |
| Phileurus didymus (Linnaeus, 1758) Elevación: 0-2,070 m | México: Campeche, Chiapas, Guerrero, Hidalgo, Jalisco, Nayarit, Oaxaca, Quintana Roo, Puebla, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán (Bates, 1888; Endrödi, 1978, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Phileurus truncatus (Palisot de Beauvois, 1807) Elevación: 0-1,600 m | México: Chiapas, Hidalgo, Jalisco, Michoacán, Nayarit, Nuevo León, Oaxaca, Puebla, Quintana Roo, Sinaloa, Sonora, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1978, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016). |
| Phileurus valgus (Olivier, 1789) Elevación: 0-1,850 m | México: Chiapas, Ciudad de México, Durango, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, Quintana Roo, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Yucatán (Bates, 1888; Endrödi, 1978, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016). |
| Phileurus voirinae Elevación: 0-900 m Agaocephalini Burmeister, 1847 Spodistes Burmeister, 1847 | México: Chiapas, Oaxaca, Veracruz (Endrödi, 1985; Ratcliffe et al., 2013). |
| Spodistes mniszechi (Thomson, 1860) Elevación: 80-1,300 m | México: Chiapas, Guerrero, Hidalgo, Oaxaca, Veracruz (Bates, 1888; Endrödi, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; presente estudio). |
| Spodistes monzoni Warner, 1992 Elevación: 90-1,800 m Dynastini MacLeay, 1819 Dynastes MacLeay, 1819 | México: Chiapas, Guerrero, Oaxaca (Warner, 1992; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014; presente estudio). |
| Dynastes hyllus Chevrolat, 1843 Elevación: 0-2,870 m | México: Chiapas, Durango, Edo. de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, San Luis Potosí, Sinaloa, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1976; Endrödi, 1985; Lachaume, 1985; Ratcliffe y Morón, 1997; Morón et al., 2003; Morón, 2009; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Dynastes maya Hardy, 2003 Elevación: 30-2,000 m | México: Chiapas, Oaxaca (Hardy, 2003; Morón, 2009; Ratcliffe et al., 2013; Moctezuma y Sánchez-Huerta, 2018). |
| Golofa Hope, 1837 | |
| Golofa incas Hope, 1837 Elevación: 850-2,800 m | México: Chiapas y Oaxaca (Endrödi, 1977, 1985; Lachaume, 1985; Morón, 1995; Ratcliffe y Morón, 1997; Morón et al., 2003; Ordóñez y Deloya, 2003; Ratcliffe et al., 2013). |
| Golofa pizarro Hope, 1837 Elevación: 0-2,850 m | México: Chiapas, Coahuila, Colima, Ciudad de México, Durango, Edo. de México, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla, Querétaro, San Luis Potosí, Tamaulipas, Veracruz (Bates, 1888; Endrödi, 1976, 1977, 1985; Lachaume, 1985; Morón, 1995; Ratcliffe y Morón, 1997; Morón et al., 2003; Ramírez-Ponce et al., 2009; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; Guzmán-Vásquez, 2016; presente estudio). |
| Golofa pusilla Arrow, 1911** Elevación: 40-1,800 m | México: Colima, Chihuahua, Edo. de México, Guerrero, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, San Luis, Potosí, Sinaloa, Sonora (Endrödi, 1977, 1985; Lachaume, 1985; Morón, 1995; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
| Golofa tersander Burmeister, 1847 Elevación: 0-1,600 m | México: Chiapas, Edo. de México, Jalisco, Nayarit, Oaxaca, San Luis Potosí, Sinaloa, Veracruz (Endrödi, 1977, 1985; Lachaume, 1985; Morón, 1995; Ratcliffe y Morón, 1997; Morón et al., 2003; Ratcliffe et al., 2013). |
| Megasoma Kirby, 1825 Megasoma elephas (Fabricius, 1775) Elevación: 0-1,500 m | México: Campeche, Chiapas, Oaxaca, Quintana Roo, Tabasco, Veracruz, Yucatán (Bates, 1888; Hardy, 1972; Endrödi, 1977, 1985; Lachaume, 1985; Ratcliffe y Morón, 1997; Morón y Gómez-Anaya, 2002; Morón et al., 2003; Morón, 2005; Ratcliffe y Morón, 2005; Ratcliffe et al., 2013). |
| Megasoma occidentale Bolívar y Pieltain, Jiménez-Asúa y Martínez, 1963** Elevación: 0-1,100 msnm | México: Colima, Guerrero, Jalisco, Michoacán, Nayarit, Oaxaca, Sinaloa (Bolívar y Pieltain et al., 1963; Hardy, 1972; Endrödi, 1977, 1985; Lachaume, 1985; Ratcliffe y Morón, 1997; Morón y Gómez-Anaya, 2002; Morón et al., 2003; Morón, 2005; Ratcliffe y Morón, 2005; Ratcliffe et al., 2013; Deloya et al., 2014, 2016; presente estudio). |
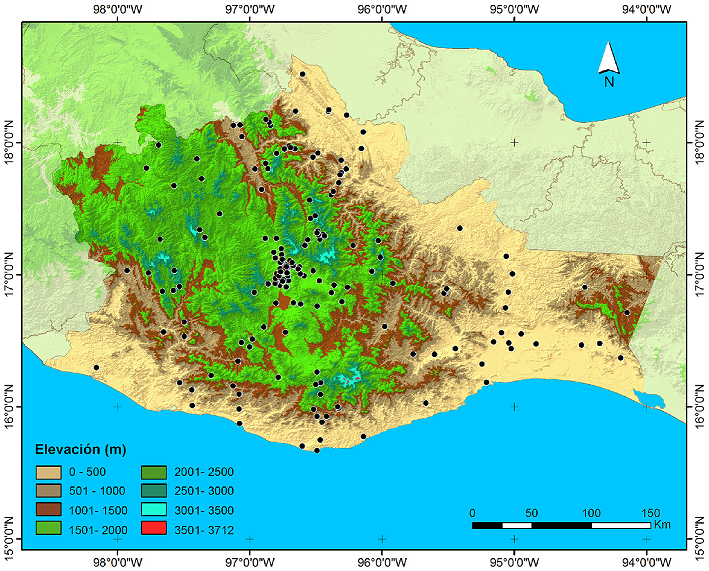
Figura 3 Modelo digital de elevación (MDE) que muestra las localidades con registros de distribución de escarabajos Dynastinae en Oaxaca, México.
La tribu mejor representada es Cyclocephalini, con 5 géneros (22.73% de la riqueza genérica) y 45 especies, que equivale a 45.92% de la riqueza de especies de dinastinos registrados en este estudio para Oaxaca. Le siguen Oryctini con 6 géneros (27.27%) y 17 especies (17.35%) y Pentodontini con 4 géneros (18.18%) y 17 especies (17.35%). La tribu con menor representación es Agaocephalini, con 1 género (4.55%) y 2 especies (2.04%) (Fig. 4). El género con mayor riqueza es Cyclocephala Dejean, 1821, con 34 especies (34.69% de la riqueza total), seguido por Orizabus Fairmaire, 1878, con 8 especies (8.16%), Tomarus Erichson, 1847 y Xyloryctes Hope, 1837, con 7 especies (7.14%) cada uno (Fig. 4). Dieciséis especies en este estudio son endémicas de México, de éstas, 3 lo son de Oaxaca (Tabla 3).

Los adultos de Cyclocephalini se encuentran activos de marzo a octubre, pero su mayor actividad es entre abril y junio (Fig. 5a). Se distribuyen entre los 0 y 2,600 m snm (Fig. 5b), aunque la mayoría de las especies se presentan entre los 0 y 1,000 m; habitan principalmente en bosque tropical caducifolio, bosque tropical perennifolio, bosque tropical subcaducifolio y bosque de pino-encino. Algunos especímenes de Cyclocephala lunulata Burmeister, 1847 (Fig. 5c) fueron recolectados con trampas cebadas con plátano o mango fermentado. Además, se los observó alimentarse del mismo fruto de guayaba junto con Cotinis mutabilis (Gory y Percheron, 1833) y Hologymnetis cinerea (Gory y Percheron, 1833) (Scarabaeidae: Cetoniinae).
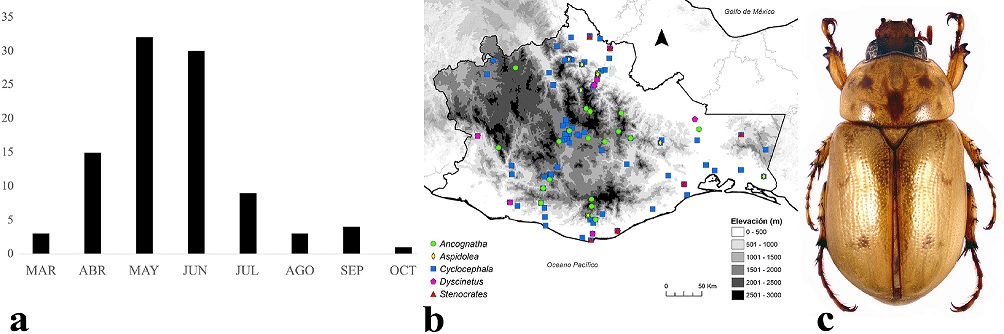
Figura 5 a) Distribución temporal de Cyclocephalini en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Cyclocephala lunulata Burmeister, 1847.
Los adultos de Pentodontini se encuentran activos de abril a noviembre, pero su mayor actividad es entre abril y junio (Fig. 6a), coincidiendo con el patrón fenológico de los Cyclocephalini. Se distribuyen entre los 0 y 3,000 m snm (Fig. 6b), aunque la mayoría de las especies lo hacen entre los 0 y 2,000 m, principalmente en bosque tropical caducifolio, bosque de pino-encino, bosque tropical subcaducifolio y bosque tropical perennifolio. Una especie representativa es Tomarus sallaei (Bates, 1888) (Fig. 6c), la cual tiene amplia distribución estatal y es común encontrarla debajo del alumbrado público junto con T. nasutus (Burmeister, 1847).
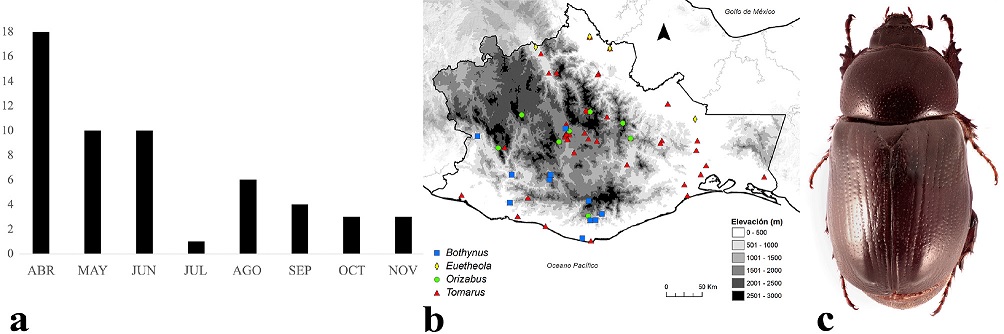
Figura 6 a) Distribución temporal de Pentodontini en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Tomarus sallaei (Bates, 1888).
Los adultos de Oryctini se encuentran activos de febrero a diciembre, pero su mayor actividad se registró en mayo y junio (Fig. 7a). Se distribuyen entre los 0 y 2,900 m snm (Fig. 7b), aunque la mayoría de las especies lo hacen entre los 0 y 2,000 m, principalmente en bosque tropical caducifolio, bosque de pino-encino, bosque tropical perennifolio y bosque tropical subcaducifolio. Una especie representativa es Xyloryctes telephus Burmeister, 1847 (Fig. 7c); tiene amplia distribución estatal y se le ha observado en junio y julio volando en grandes cantidades debajo del alumbrado público, en zonas urbanas y suburbanas de los Valles Centrales, donde suelen ser aplastados masivamente por los automóviles.
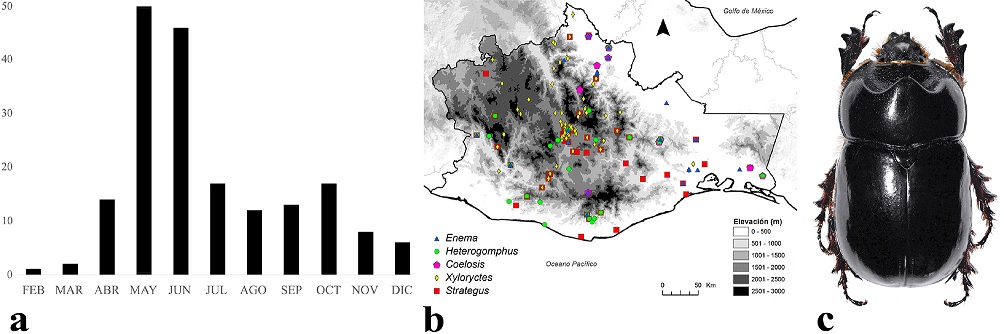
Figura 7 a) Distribución temporal de Oryctini en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Xyloryctes telephus Burmeister, 1847.
Los adultos de Phileurini se encuentran activos de abril a julio, pero su mayor actividad se registró en junio (Fig. 8a). Se distribuyen entre los 0 y 2,100 m snm (Fig. 8b), aunque la mayoría se encuentran entre los 0 y 1,000 m, principalmente en bosque tropical perennifolio, bosque tropical caducifolio y bosque tropical subcaducifolio. Una especie representativa es Phileurus didymus (Linnaeus, 1758) (Fig. 8c), la cual es atraída por la luz durante la noche, tiene amplia distribución y se le ha encontrado debajo de tocones en descomposición.
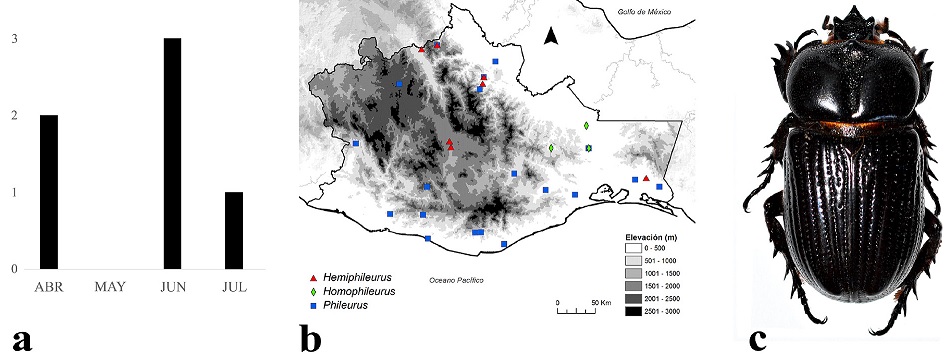
Figura 8 a) Distribución temporal de en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Phileurus didymus (Linnaeus, 1758).
Los adultos de Agaocephalini se encuentran activos de abril a junio, su mayor actividad se registró en junio (Fig. 9a). Se distribuyen entre los 80 y 1,680 m snm (Fig. 9b), aunque mayormente se les encuentra entre los 80 y 1,000 m, principalmente en bosque tropical perennifolio, bosque de pino-encino y bosque tropical subcaducifolio. De esta tribu, solamente se distribuye un género con 2 especies; la más representativa es Spodistes monzoni Warner, 1992 (Fig. 9c).
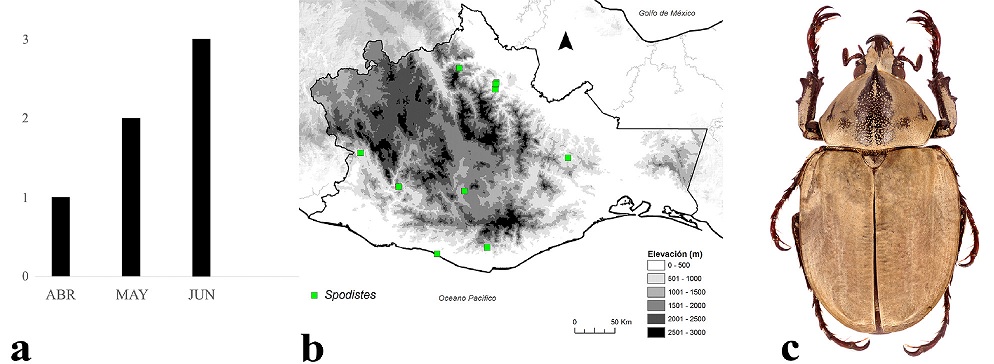
Figura 9 a) Distribución temporal de Agaocephalini en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Spodistes monzoni Warner, 1992.
Los adultos de Dynastini se encuentran activos de mayo a diciembre, pero su mayor actividad se registró en octubre (Fig. 10a). Se distribuyen entre los 0 y 2,870 m snm (Fig. 10b), aunque la mayoría lo hace entre los 0 y 1,500 m, principalmente en bosque tropical caducifolio, bosque de pino-encino, bosque tropical perennifolio y bosque tropical subcaducifolio. Una especie representativa es Dynastes hyllus Chevrolat, 1843 (Fig. 10c), tiene amplia distribución en la subprovincia Valles Centrales de Oaxaca. Se la ha encontrado durante la noche debajo del alumbrado público, en parques, escuelas y tiendas comerciales, atraídos por las luces de las lámparas. En Villa de Zaachila se recolectó 1 espécimen atraído por una carpotrampa cebada con una mezcla fermentada de plátano y cerveza.
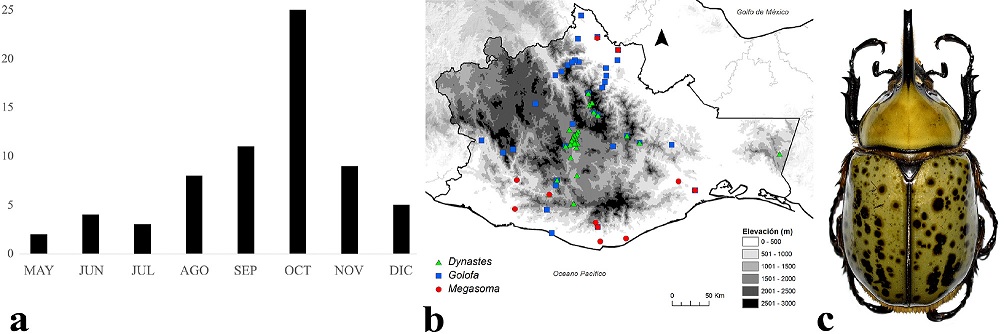
Figura 10 a) Distribución temporal de Dynastini en Oaxaca, México. b) Distribución espacial. c) Aspecto dorsal del macho de Dynastes hyllus Chevrolat, 1843.
Subprovincias fisiográficas. Se encontraron registros para 11 de las 12 subprovincias fisiográficas presentes en Oaxaca, solo la Depresión del Balsas no contó con registros (Tabla 4; Fig. 11a). La subprovincia con el mayor número de registros fue la Sierra Madre de Oaxaca con 2,523, que representa 53.08% del total de registros obtenidos en el estudio. Le siguieron Montañas y Valles del Occidente con 1,002 registros (21.08%), Valles Centrales de Oaxaca con 907 registros (19.08%). Las subprovincias menos representadas fueron la Depresión Ístmica de Tehuantepec, Fosa de Tehuacán y Sierra Madre de Chiapas con 11 (0.23%), 9 (0.19%) y 4 (0.08%) registros, respectivamente (Tabla 4). La subprovincia Sierra Madre de Oaxaca presentó la mayor riqueza con 53 especies, que equivale a 18.40% del total de la riqueza estatal de dinastinos, seguida de Valles Centrales de Oaxaca con 38 (13.19%), Montañas y Valles del Occidente con 34 (11.81%), Planicie Costera de Tehuantepec con 32 (11.11%), Planicie Costera del Golfo y Sierra Madre del Sur con 31 (10.76%) cada una. La Sierra Madre de Chiapas tiene la menor representatividad (1.39%) con 4 especies (Tabla 4).
Tabla 4 Subprovincias fisiográficas con número de tribus, géneros y especies de escarabajos Dynastinae que ocurren en Oaxaca, México. DB (Depresión del Balsas), DIT (Depresión Ístmica de Tehuantepec), FT (Fosa de Tehuacán), MVC (Montañas y Valles del Centro), MVO (Montañas y Valles del Occidente), PCT (Planicie Costera de Tehuantepec), PCG (Planicie Costera del Golfo), PCP (Planicie Costera del Pacífico), SMC (Sierra Madre de Chiapas), SMO (Sierra Madre de Oaxaca), SMS (Sierra Madre del Sur), VCO (Valles Centrales de Oaxaca).
| Subprovincia fisiográfica |
Superficie territorial en Km2 |
Registros | Tribus | Géneros | Especies | Porcentaje de especies (%) |
| SMS | 12,350 | 78 | 6 | 16 | 31 | 10.76 |
| DB | 1,788 | 0 | 0 | 0 | 0 | 0 |
| MVO | 21,263 | 1,002 | 6 | 15 | 34 | 11.81 |
| FT | 1,134 | 9 | 5 | 6 | 7 | 2.43 |
| PCG | 7,976 | 65 | 5 | 17 | 31 | 10.76 |
| SMC | 5,816 | 4 | 1 | 3 | 4 | 1.39 |
| PCT | 4,299 | 77 | 5 | 15 | 32 | 11.11 |
| DIT | 2,114 | 11 | 4 | 7 | 10 | 3.47 |
| SMO | 17,520 | 2,523 | 6 | 17 | 53 | 18.40 |
| VCO | 2,267 | 907 | 6 | 13 | 38 | 13.19 |
| MVC | 6,663 | 29 | 5 | 9 | 19 | 6.60 |
| PCP | 9,262 | 48 | 6 | 11 | 29 | 10.07 |
| 4,753 | 100 |
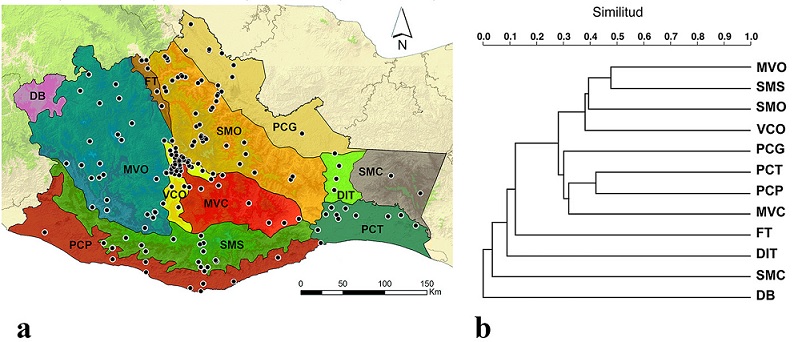
Figura 11 a) Localidades con registros de escarabajos en cada subprovincia fisiográfica de Oaxaca, México. DB (Depresión del Balsas), DIT (Depresión Ístmica de Tehuantepec), FT (Fosa de Tehuacán), MVC (Montañas y Valles del Centro), MVO (Montañas y Valles del Occidente), PCT (Planicie Costera de Tehuantepec), PCG (Planicie Costera del Golfo), PCP (Planicie Costera del Pacífico), SMC (Sierra Madre de Chiapas), SMO (Sierra Madre de Oaxaca), SMS (Sierra Madre del Sur), VCO (Valles Centrales de Oaxaca). b) Similitud de las subprovincias fisiográficas mediante el índice de Jaccard en la composición de escarabajos Dynastinae.
De acuerdo con el índice de Jaccard, basado en el algoritmo UPGMA y con un coeficiente de correlación de 0.972, se reconocen 3 principales grupos: el formado por las principales sierras (Montañas y Valles del Occidente, Sierra Madre del Sur y Sierra Madre de Oaxaca) y la subprovincia Valles Centrales de Oaxaca; el que integran las planicies costeras (Planicie Costera de Tehuantepec, Planicie Costera del Pacífico, Planicie Costera del Golfo y la subprovincia Montañas y Valles del Centro; y el conformado por el Istmo de Tehuantepec (Depresión Ístmica de Tehuantepec y Sierra Madre de Chiapas) y las subprovincias Depresión del Balsas y Fosa de Tehuacán (Fig. 11b). Las subprovincias Montañas y Valles del Occidente y Sierra Madre del Sur presentan la mayor similitud en la composición de especies de escarabajos con 48%. La Sierra Madre del Sur y la Sierra Madre de Oaxaca comparten el 42% de similitud. La Planicie Costera del Golfo y la Planicie Costera del Pacífico también presentan 42% de similitud. Las Montañas y Valles del Occidente tienen 41% de similitud con los Valles Centrales de Oaxaca (Fig. 11b). La Sierra Madre de Chiapas y la Sierra Madre de Oaxaca presentaron la menor similitud con solo 4%.
Se obtuvieron nuevos registros de distribución altitudinal para 27 especies (Tabla 5). Se consideraron únicamente aquellos registros que se extendieron como mínimo 100 m por debajo o encima de su rango altitudinal. Los nuevos registros altitudinales se obtuvieron con las localidades de colecta o registros encontrados en la literatura y obtenidas las altitudes con el MDE. Xyloryctes Hope, 1837 presentó el rango de distribución altitudinal más amplio, de 0 a 2,900 m snm. A nivel de especie, X. telephus Burmeister, 1847 tiene el mayor rango altitudinal, ya que se le puede encontrar desde 0 hasta 2,870 m snm. Otra especie con amplio rango de distribución altitudinal (70-2,600 m) fue Ancognatha falsa Arrow, 1911. Orizabus delgadoi Ratcliffe y Cave, 2010 es el único dinastino que se ha reportado a 3,000 m de altitud en Oaxaca. Los estratos altitudinales con mayor número de registros fueron EA500 con 2,601 que equivale a 54.72% del total de registros (4,753), seguido de EA2000 con 995 registros que representa 20.93%. Los estratos con menor número de registros fueron EA3500 y EA3000 con 1 y 5 registros, respectivamente (Tabla 6).
Tabla 5 Nuevos registros altitudinales para 27 especies de Dynastinae con distribución en Oaxaca, México. RangL (rango reportado en literatura); RangE (rango obtenido en este estudio); Dif-deb (diferencia obtenida por debajo del rango); Dif-enc (diferencia obtenida por encima del rango).
| Tribu | Especies | RangL (m) | RangE (m) | Dif-deb (m) | Dif-enc (m) | |
| 1 | Cyclocephalini | Ancognatha falsa | 960-2360 | 70-2,600 | 890 | 240 |
| 2 | Cyclocephalini | Ancognatha quadripunctata | 1,200-2,100 | 80-2,250 | 1,120 | 150 |
| 3 | Cyclocephalini | Ancognatha sellata | 1,500-2,450 | 80-1,630 | 1,420 | - |
| 4 | Cyclocephalini | Aspidolea singularis | 0-1,600 | 60-2,030 | - | 430 |
| 5 | Cyclocephalini | Cyclocephala concolor | 1,000-2,000 | 40-1,820 | 960 | - |
| 6 | Cyclocephalini | Cyclocephala curta | 100-1,200 | 0-900 | 100 | - |
| 7 | Cyclocephalini | Cyclocephala jalapensis | 1,369-2,040 | 80-1550 | 1,289 | - |
| 8 | Cyclocephalini | Cyclocephala larssoni | 0-700 | 0-2,040 | - | 1340 |
| 9 | Cyclocephalini | Cyclocephala picta | 900-2,000 | 0-1,610 | 900 | - |
| 10 | Cyclocephalini | Cyclocephala stictica | 0-1,650 | 0-2,030 | - | 380 |
| 11 | Cyclocephalini | Dyscinetus laevipunctatus | 0-500 | 0-800 | - | 300 |
| 12 | Oryctini | Enema endymion | 35-1,700 | 0-1,800 | 35 | 100 |
| 13 | Oryctini | Heterogomphus flohri | 500-3,500 | 0-1,650 | 500 | - |
| 14 | Oryctini | Xyloryctes ensifer | 1,800-3,089 | 1,540-2,340 | 260 | - |
| 15 | Oryctini | Xyloryctes furcatus | 375-2,860 | 0-1,880 | 375 | - |
| 16 | Oryctini | Xyloryctes lobicollis | 330-2,500 | 0-1,880 | 330 | - |
| 17 | Oryctini | Xyloryctes telephus | 530-3,400 | 0-2,900 | 530 | - |
| 18 | Oryctini | Xyloryctes teuthras | 800-2,400 | 580-2,090 | 220 | - |
| 19 | Oryctini | Xyloryctes thestalus | 550-2,900 | 0-2,250 | 550 | - |
| 20 | Phileurini | Hemiphileurus flohri | 1,500-1,940 | 1,000-1,550 | 500 | - |
| 21 | Phileurini | Hemiphileurus microps | 550-1,000 | 80 | 470 | - |
| 22 | Phileurini | Phileurus didymus | 0-1,800 | 0-2,070 | - | 270 |
| 23 | Phileurini | Phileurus voirinae | 100-900 | 0-210 | 100 | - |
| 24 | Agaocephalini | Spodistes mniszechi | 300-1,300 | 80-1,200 | 220 | - |
| 25 | Agaocephalini | Spodistes monzoni | 500-1,800 | 90-1,680 | 410 | - |
| 26 | Dynastini | Dynastes hyllus | 0-2,100 | 860-2,870 | - | 770 |
| 27 | 2Dynastini | Golofa pizarro | 200-2,850 | 0-2,330 | 200 | - |
Tabla 6 Clasificación de los registros de escarabajos Dynastinae por estrato altitudinal en Oaxaca, México. EA = estrato altitudinal; msnm = metros sobre el nivel del mar.
| Número | Rango altitudinal (m snm) |
Estrato | Registros | Porcentaje (%) |
| 1 | 0-500 | EA500 | 2601 | 54.72 |
| 2 | 5,001-1,000 | EA1000 | 615 | 12.94 |
| 3 | 1,001-1,500 | EA1500 | 203 | 4.27 |
| 4 | 1,501-2,000 | EA2000 | 995 | 20.93 |
| 5 | 2,001-2,500 | EA2500 | 333 | 7.01 |
| 6 | 2,501-3,000 | EA3000 | 5 | 0.11 |
| 7 | 3,001-3,500 | EA3500 | 1 | 0.02 |
| 4,753 | 100 |
La mayor riqueza para la distribución observada la presentó el estrato EA500 con 74 especies que equivale a 30% de la riqueza total, seguido del estrato EA1000 y EA2000, ambos con 47 especies que equivale a 19.50% para cada estrato (Tabla 7). En cuanto a la distribución potencial, los estratos con mayor riqueza fueron EA500 y EA1000 con 74 (27.50%) y 61 (22.68%) especies, respectivamente. En contraste, los estratos con menor riqueza para ambas distribuciones fueron EA3500 y EA3000 con 1 y 4 especies respectivamente, lo que representa 0.41% y 1.66% de la riqueza total para la distribución observada y 0.37% y 1.49% para la distribución potencial (Tabla 7). Los estratos de los extremos no difieren para ninguna de las distribuciones, esto se debe a que son los estratos de apertura y cierre de todo el rango altitudinal estudiado (0-3,500 m), por lo cual solo se encontraron diferencias para los estratos intermedios. La comparación entre la distribución observada y la distribución potencial muestra que, para esta última los estratos EA1000 y EA1500 presentaron el mayor incremento en sus riquezas con 14 y 12 especies, respectivamente. El EA2000 solo incrementó 2 especies (Tabla 7). Se observó que los estratos con datos de distribución potencial mostraron mayor riqueza que sus homólogos para la distribución observada, a excepción de EA500, EA2500, EA3000 y EA3500 que permanecieron iguales para ambas distribuciones (Tabla 7; Fig. 12). Por otra parte, se pueden observar claramente 2 picos de riqueza (EA500 y EA2000) para la distribución observada. En cambio, en la distribución potencial solo hubo un pico máximo de riqueza en EA500, el cual fue disminuyendo paulatinamente conforme la altitud incrementó.
Tabla 7 Distribución observada y potencial de la riqueza de escarabajos Dynastinae por estratos altitudinales en Oaxaca, México. DistOBS = distribución observada; DistPOT = distribución potencial.
| DistOBS | DistPOT | Diferencia (Spp.) |
|||
| Estratos | Riqueza (Spp.) |
Porcentaje (%) |
Riqueza (Spp.) |
Porcentaje (%) |
|
| EA500 | 74 | 30.71 | 74 | 27.51 | 0 |
| EA1000 | 47 | 19.50 | 61 | 22.68 | 14 |
| EA1500 | 40 | 16.60 | 52 | 19.33 | 12 |
| EA2000 | 47 | 19.50 | 49 | 18.22 | 2 |
| EA2500 | 28 | 11.62 | 28 | 10.41 | 0 |
| EA3000 | 4 | 1.66 | 4 | 1.49 | 0 |
| EA3500 | 1 | 0.41 | 1 | 0.37 | 0 |
| 100 | 100 | ||||
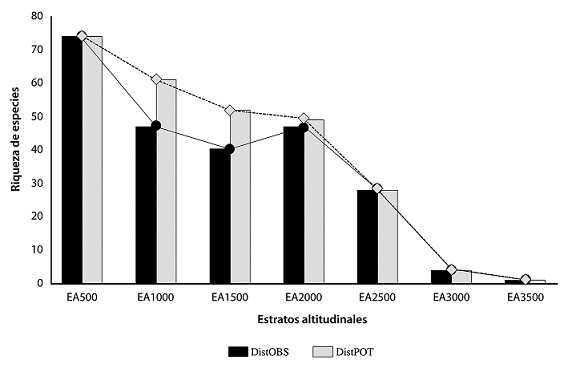
Figura 12 Distribución observada y potencial de la riqueza de Dynastinae por estrato altitudinal en Oaxaca, México.
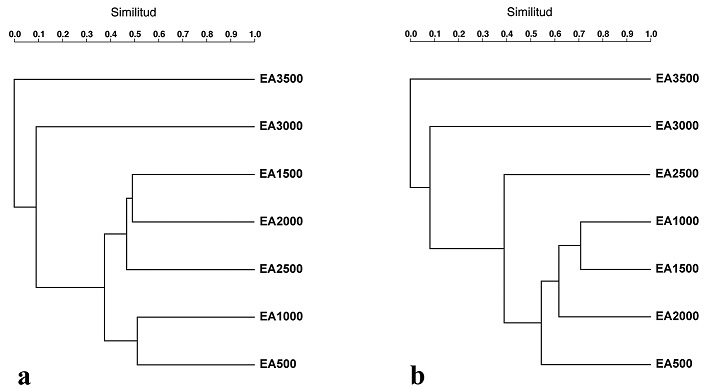
El dendrograma de similitud basado en el índice de Jaccard para la distribución observada mostró muy poca semejanza entre los estratos, ya que el valor más alto de similitud (51%) se encontró entre los estratos EA1000 y EA500, seguido de EA1500, EA2000 y EA2500, que presentaron valores de similitud entre 47 y 49% (Fig. 13a). Por su parte, el índice de Jaccard para la distribución potencial de la riqueza de especies mostró que los valores más altos de similitud (71%) se encontraron entre los estratos EA1000 y EA1500, y EA1500 y EA2000, seguidos de EA500 y EA1000 con 69% de similitud. Para ambas distribuciones, la menor similitud se observó entre los estratos EA500 y EA3000 con apenas 4% (Fig. 13a, b).
Discusión
Como consecuencia de varios trabajos recientemente publicados, en donde se reportan nuevos registros de distribución, nuevas especies y sinonimias (Delgado y Mora-Aguilar, 2020; Hielkema, 2017; López-García y Deloya, 2018; López-García et al., 2016, 2018; Moore et al., 2015, 2018; Ratcliffe y Cave, 2015; Ratcliffe et al., 2013; Romero-López y Morón, 2017), se tiene que a la fecha, la fauna de Dynastinae de México está integrada por 29 géneros y 188 especies, de los cuales Oaxaca cuenta con 22 géneros y 98 especies, que representan 75.86% y 52.12% de la riqueza genérica y específica nacional. Esto significa que Oaxaca alberga poco más de la mitad de las especies de México, ocupando el tercer lugar a nivel nacional, después de Chiapas y Veracruz, con 103 y 102 especies, respectivamente (Tabla 8). La riqueza faunística de Oaxaca también es comparable con la riqueza que presentan países como Honduras (100 spp., Ratcliffe y Cave, 2006) y Guatemala (112 spp., Ratcliffe et al., 2013), incluso es mucho mayor que la riqueza reportada para Belice (51 spp., Ratcliffe et al., 2013), El Salvador (59 spp.) y Nicaragua (78 spp., Ratcliffe y Cave, 2006). Esta alta riqueza se debe principalmente a la compleja heterogeneidad ambiental que presenta el estado (Lavariega et al., 2016; Martín et al., 2019; Ordoñez y Rodríguez, 2008; Santos-Moreno, 2014), esto como consecuencia de la accidentada orografía, con hábitats montañosos de tierras altas, bajas y valles (García-Mendoza et al., 2004; Peterson et al., 2003), que junto con la posición geográfica, que lo ubica en una zona estratégica con influencia de las corrientes de viento del golfo de México y del Pacífico (Peterson et al., 2003), permiten una gran variedad de climas, microclimas, tipos de vegetación, y condiciones físicas y ecológicas que favorecen la diversidad de escarabajos, condiciones (por ejemplo, altitud, tipo de suelo, precipitación, humedad, etc.) que ya han sido mencionadas como causantes de alta biodiversidad en México (Morón, 2004; Trujillo-Miranda et al., 2016). Además, ocurre un efecto de ecotono que permite la convergencia de la fauna proveniente de las regiones Neotropical y Neártica (Martín et al., 2019; Morón, 2004; Peterson et al., 2003).
Tabla 8 Número de tribus, géneros y especies de Dynastinae por entidad federativa en México. Datos recopilados de Delgado y Mora-Aguilar, 2020; Deloya y Ponce, 2016; López-García et al., 2016, 2018; López-García y Deloya, 2018; Lugo-García et al., 2013; Minor y Morón, 2016; Moore et al., 2015, 2018; Ratcliffe et al., 2013; Ratcliffe y Cave, 2015; Ratcliffe y Morón, 1997; Romero-López y Morón, 2017; Zaragoza-Caballero et al., 2019.
| Estado | Tribus | Géneros | Especies | Estado | Tribus | Géneros | Especies |
| Aguascalientes | 3 | 6 | 13 | Morelos | 5 | 14 | 37 |
| Baja California | 4 | 5 | 11 | Nayarit | 5 | 17 | 57 |
| Baja California Sur | 4 | 5 | 11 | Nuevo León | 5 | 11 | 23 |
| Campeche | 5 | 10 | 16 | Oaxaca | 6 | 22 | 98 |
| Chiapas | 6 | 24 | 103 | Puebla | 5 | 17 | 53 |
| Chihuahua | 5 | 12 | 26 | Querétaro | 4 | 8 | 17 |
| Ciudad de México | 5 | 10 | 26 | Quintana Roo | 5 | 10 | 20 |
| Coahuila | 5 | 10 | 20 | San Luis Potosí | 5 | 17 | 43 |
| Colima | 5 | 10 | 25 | Sinaloa | 5 | 16 | 43 |
| Durango | 5 | 12 | 38 | Sonora | 5 | 17 | 45 |
| Estado de México | 4 | 11 | 38 | Tabasco | 5 | 15 | 33 |
| Guanajuato | 4 | 8 | 16 | Tamaulipas | 5 | 17 | 40 |
| Guerrero | 6 | 18 | 60 | Tlaxcala | 3 | 4 | 7 |
| Hidalgo | 6 | 17 | 49 | Veracruz | 6 | 23 | 102 |
| Jalisco | 5 | 21 | 75 | Yucatán | 5 | 9 | 22 |
| Michoacán | 5 | 14 | 52 | Zacatecas | 3 | 8 | 19 |
La subprovincia Sierra Madre de Oaxaca concentra la mayor riqueza (53 especies) y número de registros (2523) debido principalmente al gradiente de pisos altitudinales (0-3,500 m), climas (desde cálidos húmedos y subhúmedos, hasta templados y fríos) y gran cantidad de bosques bien conservados, entre los que destacan los de pino, encino, mixto, mesófilo de montaña y bosques tropicales (INEGI, 2016). Se ha documentado que los sistemas montañosos contribuyen desproporcionadamente a la biodiversidad, ya que contienen más de 85% de especies de anfibios, aves y mamíferos a nivel mundial. Además, las características climáticas complejas que presentan estos sistemas son responsables del mantenimiento y diversidad biológica que albergan (Arriaga-Jiménez et al., 2020; Rahbek et al., 2019). Por su atractivo, esta región (SMO) ha sido más explorada que otras a lo largo del tiempo, lo que se traduce en un mayor número de registros o esfuerzo de muestreo (Briones-Salas et al., 2015). Además, la Sierra Madre de Oaxaca presenta una compleja historia biogeográfica de formación, aislamiento geográfico y climático que favorecen los procesos de especiación y endemismo, por lo cual ha sido considerada una zona de importancia biológica y región prioritaria para la conservación (Arriaga et al., 2000; Briones-Salas et al., 2015; Lavariega et al., 2016). Resultados similares de riqueza también se han encontrado en estudios con otros taxones como mamíferos, aves, reptiles, anfibios y plantas, por lo cual no es de sorprender que esta región (SMO) es una de las más diversas del estado (Briones-Salas et al., 2015; García-Grajales, 2008; García-Mendoza, 2004; Lavariega et al., 2016; Martín et al., 2019). En contraste, la baja representatividad (4 registros y 4 especies) de la Sierra Madre de Chiapas se debe principalmente a la falta de muestreo, debido al difícil acceso a diferentes zonas, por ejemplo, Los Chimalapas, una de las regiones más biodiversas de México y centro importante de endemismos, considerada también región prioritaria para la conservación (Arriaga et al., 2000; Olguín-Monroy et al., 2008; Peterson et al., 2003). En este contexto, se puede señalar que, así como Los Chimalapas, hay otras regiones en el estado que no han sido exploradas, muestra de ésto es la subprovincia Depresión del Balsas (Briones-Salas et al., 2015; Tabla 4, Fig. 11b), donde no se encontró registro alguno, por lo cual es necesario muestrear estas regiones y documentar la diversidad de Dynastinae, ya que se asume que aún hay especies por registrar o describir en el estado. Por otra parte, el análisis de similitud muestra que hay un fuerte recambio de especies entre subprovincias fisiográficas, ya que la mayor similitud fue de apenas 48%, en otras palabras, la baja similitud (0.04-0.48) es responsable de la gran diversidad de escarabajos en Oaxaca, así como se ha reportado para otros grupos, como los mamíferos (Briones-Salas et al., 2015). Además, es importante mencionar que los registros representan principalmente colectas esporádicas no intensivas ni sistemáticas, y existen muchas áreas que no han sido exploradas. Debido a sus características fisiográficas, climáticas, edáficas y de vegetación, son regiones con enorme potencial de estudio, principalmente aquellas que son consideradas áreas prioritarias para la conservación (Arriaga et al., 2000).
Se puede señalar que los nuevos registros altitudinales se deben a tres factores: 1) la falta histórica de muestreos, ésto porque son pocos los investigadores interesados en el grupo, en comparación con otros taxones como aves, mamíferos, reptiles, anfibios, etc. (Briones-Salas et al., 2015; García-Grajales, 2008; Lavariega et al., 2016; Martín et al., 2019), que son más apreciados, incluso por la sociedad; 2) especímenes con falta de datos (altitud y/o coordenadas) o mal etiquetados (datos ambiguos, generales o localidades erróneas); 3) la acelerada pérdida de su hábitat natural, como consecuencia de las actividades antropogénicas y el cambio climático (Macedo et al., 2018; Maciel-Mata et al., 2015). Aquí hemos documentado que los escarabajos son sensibles a la perturbación (Otavo et al., 2013; Trujillo-Miranda et al., 2016), por consiguiente, ésto los obliga a desplazarse a zonas con ambientes más favorables, ya sea a mayor o menor altitud según los requerimientos y tolerancia ecológica de cada especie; por ejemplo, si una especie X se distribuye en una zona de baja altitud con clima cálido y la temperatura aumenta considerablemente, ella se desplazará a mayor altitud en busca de condiciones ambientales óptimas (Foden et al., 2007; Maciel-Mata et al., 2015). Es importante conocer la distribución altitudinal de estas especies, ya que el ensamble varía conforme la altitud cambia (Stevens, 1992; Trujillo-Miranda et al., 2016), esto nos puede ayudar a entender el comportamiento o respuesta de las especies ante condiciones ecológicas adversas, así como conocer los patrones de distribución de la riqueza de especies, el estado de supervivencia a lo largo del tiempo (dinámica poblacional) y el papel que éstas desempeñan en los ecosistemas. Por ejemplo, en el caso de los Cyclocephalini, se ha reportado que son importantes polinizadores de más de 161 especies de plantas (Moore y Jameson, 2013), con excepción de algunas especies de Cyclocephala y Dyscinetus, cuyas larvas pueden constituir plagas agrícolas importantes (Morón, 2004; Morón y Aragón, 2003). Por su parte, las especies de Dynastini, Oryctini y Phileurini son importantes degradadoras de materia orgánica (Morón, 1985, 2004). En el caso de Spodistes, no se conoce mucho de su historia natural (Ratcliffe et al., 2013).
Xyloryctes Hope, 1837 fue el género con mayor amplitud de rango altitudinal, esta amplitud se debe al traslape de los rangos en el que se presenta cada una de sus especies, por ejemplo X. telephus Burmeister, 1847, que se distribuye entre 0 y 2,870 m de altitud, especie que tiene el rango más amplio en este estudio. Bitar y Morón (2014) ya habían reportado que la distribución altitudinal de esta especie es una de las más amplias del género, 530-2,275 m. Por su parte, Chacón et al. (2013) registraron esta especie del Iztaccíhuatl, a 3,356 m, todavía a una altitud mucho mayor a la reportada en este estudio. También se ha dicho que las características ambientales favorecen la dispersión de estos escarabajos, los cuales, además, presentan alta tolerancia ecológica (Bitar y Morón, 2014), por consiguiente, son menos sensibles a la perturbación antrópica de su hábitat, ya que incluso en este estudio, se les encontró con frecuencia en grandes cantidades debajo del alumbrado público en zonas urbanas y suburbanas en la parte central del estado. Otra especie con un amplio rango de distribución altitudinal es Ancognatha falsa Arrow, 1911 (70-2,600 m). El hecho de ocupar un amplio rango se debe, además de la tolerancia ecológica, a la capacidad de movilidad, ya que estos escarabajos pueden desplazarse grandes distancias, como se ha reportado para otras especies de Cyclocephalini del género Cyclocephala Dejean, 1821, las cuales pueden llegar a recorrer distancias de hasta 1.5 km de planta a planta en 24 hrs. (Morón, 1997).
En cuanto a la similitud de la distribución observada y potencial de Dynastinae, los valores bajos de similitud (menores a 51%) para la distribución observada revelan que hay un fuerte recambio de especies entre estratos, quizás esto se deba en parte a la intensidad de muestreo, aunque también coincide con los valores bajos de similitud encontrados entre subprovincias. Mientras aumenta la altitud, la similitud en la riqueza de especies entre los estratos disminuye. En contraste con la distribución observada, la distribución potencial presentó valores más altos de similitud, esto se debe a que hay especies que tienen amplios rangos altitudinales de distribución. Por otra parte, de acuerdo con lo observado en este estudio, al colocar valores potenciales de distribución se pueden obtener resultados más cercanos a la distribución real de cada especie de Dynastinae. Esta distribución potencial nos permite llenar vacíos de información por falta de registros. En general, ambas distribuciones muestran un patrón similar que indica una relación inversamente proporcional entre la diversidad de escarabajos y la altitud, ya que a mayor altitud menor fue la diversidad; por el contrario, a menor altitud mayor fue la diversidad de estos insectos (Fig. 12). Este patrón de distribución parece ajustarse más a un modelo lógico clásico que coincide con la regla altitudinal de Rapoport propuesta por Stevens (1992), la cual señala que la riqueza de especies de insectos y otros taxones como árboles, mamíferos, aves, reptiles y anfibios, será mayor en las zonas bajas que en las partes altas de las montañas, mientras más se incremente la elevación, menos especies habrá pero su rango altitudinal incrementará. Por el contrario, especies de zonas bajas tendrán rangos de distribución altitudinal menores o estrechos, lo que a su vez permitirá la coexistencia de muchas especies, lo cual reflejará mayor diversidad (McCain y Bracy-Knight, 2012; Rahbek, 1995; Šizling et al., 2009). La explicación dada por Stevens para entender este fenómeno es que las condiciones climáticas (temperatura, humedad, precipitación) serán las determinantes de la riqueza local de especies, ya que especies de zonas altas tendrán mayor tolerancia a los cambios climáticos, por lo cual, sus rangos de distribución altitudinal serán mayores, pues estarán mejor adaptadas a condiciones adversas. En contraste, especies de zonas bajas tendrán menos tolerancia y sus rangos de distribución altitudinal serán menores, ya que serán más sensibles a las condiciones climáticas (Stevens, 1992).

