











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
A nivel mundial, se han descrito 49,118 especies de arañas agrupadas en 4,206 géneros y 128 familias (World Spider Catalog, 2021). Únicamente 6 géneros con 212 especies (4%) se consideran de importancia médica debido a los efectos nocivos que ocasiona su mordedura venenosa a la salud humana: Sicarius Walckenaer, 1847 (familia Sicariidae) (21 especies); Hexophthalma Karsh, 1879 (Sicariidae) (8); Loxosceles Heineken y Lowe, 1832 (Sicariidae) (140); Atrax O. Pickard-Cambridge, 1877 (Atracidae) (3); Phoneutria Perty, 1833 (Ctenidae) (8), y Latrodectus Walckenaer, 1805 (Theridiidae) (32), (Müller, 1993; Valdez-Mondragón et al., 2018, 2019; World Spider Catalog, 2021). Al menos otros 20 géneros de arañas pueden requerir algún tipo de atención médica, pero sin poner en riesgo la vida humana en comparación con los 6 géneros anteriormente mencionados (Müller, 1993; Valdez-Mondragón et al., 2018). Algunos ejemplos son los géneros Lycosa Latrielle, 1804 (Lycosidae); Sericopelma Ausserer, 1875; Poecilotheria Simon, 1885; Phamphobeteus Pocock, 1901 (Theraphosidae), Cheiracanthuium C. L. Koch, 1839 (Eutichuridae) y Argyroneta Latreille, 1804 (Dictynidae) (Barreto y Barreto, 1994; Foelix, 2011; World Spider Catalog, 2021). Las arañas del género Latrodectus son consideradas de importancia médica debido a que su veneno contiene la enzima α-latrotoxina, que en vertebrados tiene una gran afinidad por receptores presinápticos, ocasionando la liberación masiva de neurotransmisores, lo que genera parálisis del sistema nervioso central (Aguilera, 2016; Garb et al., 2004; Kaslin, 2013; Müller, 1993).
En México, actualmente se encuentran 2,349 especies de arañas, agrupadas en 426 géneros y 69 familias, lo que representa 4.8%,10.2% y 57.5% de la diversidad mundial respectivamente (Jiménez e Ibarra, 2008; World Spider Catalog, 2021). Los géneros Loxosceles con 40 especies y Latrodectus con 3 especies son los únicos de importancia médica, lo que representa 1.8% de la diversidad total de arañas del país (Desales-Lara et al., 2018; Valdez-Mondragón et al., 2018, 2019).
La familia Theridiidae actualmente ocupa el quinto lugar en cuanto a diversidad de especies de arañas en el mundo, con 2,516 clasificadas en 124 géneros (World Spider Catalog, 2021). De esta familia, México registra 34 géneros y 223 especies (27% y 9%), respectivamente a nivel mundial (Jiménez e Ibarra, 2008). El género Latrodectus está conformado por 32 especies, distribuidas en todos los continentes a excepción de los polos. Argentina con 9 especies, es el país con mayor riqueza mundial (Cabrera-Espinosa, 2020; World Spider Catalog, 2021). Latrodectus geometricus C. L. Koch, 1841 tiene una distribución cosmopolita, y ha sido introducida en varios países incluido México (Cabrera-Espinosa, 2020; Chamberlin e Ivie, 1935; Garb et al., 2004; Kaslin, 2013; World Spider Catalog, 2021).
Los análisis filogenéticos del género Latrodectus con el gen mitocondrial CO1 (Aguilera et al., 2009; Garb et al., 2004), sugieren que las especies se agrupan en 2 clados principales: el clado geometricus, que incluye a L. geometricus, Latrodectus rhodesiensis Mackay, 1972 y Latrodectus umbukwaneWright, Wright, Lyle y Engelbrecht, 2019, y el clado mactans que incluye al resto de las especies. Estos 2 clados son soportados morfológicamente por la posición de las espermatecas de las hembras, en forma de “V” en el clado mactans y casi paralelas en el clado geometricus (Aguilera et al., 2009; Garb et al., 2004).
Conocidas coloquialmente como “viudas negras” o “black widow spiders” en inglés, “arañas capulinas”, “casampulga” o “cintlatlahua” (la del trasero rojo en náhuatl) en México, actualmente se conocen 3 especies del género Latrodectus: Latrodectus mactans (Fabricius, 1775), Latrodectus hesperusChamberlin e Ivie, 1935 y L. geometricus. Hasta ahora, L. mactans es la especie con mayor distribución en el territorio nacional (Cabrera-Espinosa, 2020; Cabrera-Espinosa y Valdez-Mondragón, 2019; Monjaraz-Ruedas, 2012; REDTOX, 2021). Los registros de L. hesperus son escasos y recientes en nuestro país (Desales-Lara et al., 2018; Jiménez et al., 2015). Esto se debe a que morfológicamente, presenta un solapamiento de caracteres diagnósticos como pedipalpos en machos y ductos copulatorios en hembras con L. mactans (Cabrera-Espinosa, 2020).
En los primeros estudios taxonómicos realizados con este género, entre 1902 y 1958, la identificación de especies estaba basada únicamente en estructuras somáticas como patrones de coloración y tipos de sedas del opistosoma, estableciéndose variedades o subespecies para L. mactans: L. mactans mactans, L. mactans hesperus, L. mactans mexicanus, L. mactans texanus y L. mactans albomaculatus (Chamberlin e Ivie, 1935; Levi, 1959; Pickard-Cambridge, 1902; World Spider Catalog, 2021). Levi (1959) utiliza estructuras sexuales primarias (pedipalpos en machos y epiginios en hembras), y se invalidan las subespecies que pertenecían en realidad a L. mactans con excepción de L. mactans hesperus, reconocida como una especie válida: L. hesperus.
Actualmente, la determinación de especies de Latrodectus se realiza utilizando la taxonomía tradicional que hace uso de caracteres sexuales primarios y patrones de coloración del opistosoma. Sin embargo, estos caracteres no siempre son confiables debido al amplio grado de variación morfológica e incluso solapamiento de caracteres (Cabrera-Espinosa, 2020; Cabrera-Espinosa y Valdez-Mondragón, 2019; Levi, 1959; Levi y Randolph, 1975; Levy y Amitai, 1983; Ubick et al., 2005).
El género Latrodectus en México presenta una amplia distribución a nivel nacional, aunado a la gran cantidad de registros provenientes de diversas fuentes de información (colecciones científicas, literatura y bases de datos en línea). A pesar de esto, la distribución actual de las 3 especies del género es incierta y desactualizada (Monjaraz-Ruedas, 2012). Además, los patrones dorsales de variación sobre todo en L. mactans, nunca han sido estudiados biogeográficamente, por lo que es necesario conocer si existe un patrón biogeográfico de éstas.
Debido a lo anterior, el objetivo de este trabajo fue actualizar los registros de las especies de Latrodectus de México, así como estudiar los patrones de variación dorsales y ventrales del opistosoma de las hembras dentro de un contexto biogeográfico, así como la distribución real y potencial de las 3 especies del género en el país. Se discute la variación morfológica de estructuras diagnósticas, tales como ductos copuladores en las hembras y pedipalpos en los machos, y sus implicaciones taxonómicas.
Materiales y métodos
Se revisaron ejemplares depositados en las siguientes colecciones nacionales: Colección Nacional de Arácnidos (CNAN), Instituto de Biología (IBUNAM), UNAM, México (curador: Dr. Oscar F. Francke Ballvé); Laboratorio de Aracnología (LATLAX), del Laboratorio Regional de Biodiversidad y Cultivo de Tejidos Vegetales (LBCTV), Instituto de Biología UNAM, sede Tlaxcala (curador Dr. Alejandro Valdez Mondragón); y Colección Aracnológica (CARCIB), del Centro de Investigación Biológica del Noreste S. C. (CIBNOR) (curadora Dra. María Luisa Jiménez Jiménez) (Tabla 1).
Tabla 1 Total de registros obtenidos de las distintas fuentes de información para cada una de las especies de Latrodectus de México.
| Colección | L. mactans | L. geometricus | L. hesperus | L. sp. | Número de registros |
|---|---|---|---|---|---|
| GBIF | 448 | 57 | 1 | - | 506 |
| CNAN | 249 | 24 | - | 26 | 299 |
| Naturalista | - | 166 | - | - | 166 |
| Literatura | 46 | 28 | 3 | 1 | 78 |
| LATLAX | 37 | 15 | 2 | 21 | 75 |
| 2 | 20 | - | - | 22 | |
| CARCIB | - | - | 1 | - | 1 |
| Totales | 782 | 310 | 7 | 48 | 1,147 |
Se realizaron 17 salidas al campo para la recolecta de ejemplares de Latrodectus durante los años 2017 a 2019 en los siguientes estados de la República Mexicana: Baja California, Baja California Sur, Estado de México, Guerrero, Hidalgo, Jalisco, Michoacán, Nayarit, Oaxaca, Puebla y Tlaxcala. Los ejemplares se recolectaron de forma manual con pinzas aracnológicas y frascos de plástico. Todos los ejemplares se etiquetaron con sus datos correspondientes, colocados en viales y frascos de vidrio en alcohol etílico al 80% y depositados en la colección aracnológica del LATLAX.
La identificación de ejemplares se realizó con los trabajos de Levi (1959), Levi y Randolph (1975) y Levy y Amitai (1983). Para la categoría de especie, se disectaron epiginios en hembras y pedipalpos en machos adultos, siguiendo a Levi (1959) y Valdez-Mondragón (2009). Los epiginios fueron sumergidos en KOH (10%) durante 10 minutos para remover y limpiar tejido blando alrededor de las espermatecas y ductos copuladores. Las disecciones se realizaron con ayuda de un microscopio estereoscópico Zeiss Stemi 508. Los ejemplares y las estructuras disectadas, fueron observadas bajo alcohol etílico al 80% en una caja Petri con arena blanca de playa, bajo un microscopio estereoscópico Zeiss Stemi 508. Para las fotografías se utilizó una cámara Axiocam 506 acoplada a un microscopio estereoscópico Zeiss AXIO Zoom V16. Las imágenes fueron procesadas con el software Zen 2 (blue edition). Para fotografiar pedipalpos y ductos copuladores, se colocaron en alcohol etílico (70%) en gel, cubierto por alcohol etílico al 80% para evitar difracción, y se editaron en el programa Photoshop CS6 Extended. La variación de los patrones dorsales del opistosoma fue observada únicamente en hembras adultas siguiendo a Levi (1959). El criterio para determinar los diferentes tipos de patrones dorsales del opistosoma, se tomaron en cuenta aquellos que contaran con la presencia en al menos 10 individuos. Los patrones dorsales que contaran con un número menor a 10 se agruparon con el patrón dorsal más parecido. Dado que no existe una relación aparente entre el patrón ventral y el patrón dorsal del opistosoma, la variación del “reloj de arena” se registró sin seguir ningún criterio específico.
Se elaboró una base de datos en Excel con los datos obtenidos tanto de colecciones científicas revisadas, como de ejemplares recolectados en campo, registros de literatura y grupos especializados de redes sociales en línea (Facebook) como: Insectos y otros artrópodos de México, Arácnidos de la República Mexicana, Arañas de México y Reivindicando a las arañas. También se utilizaron las bases de datos del GBIF (GBIF.org, 2019), Naturalista (Naturalista, 2019) y del Portal de Datos Abiertos de la UNAM (2018). Para considerar los registros obtenidos de dichas fuentes (Facebook, GBIF y Naturalista), se tomaron en cuenta los siguientes criterios: 1) pertenecer a un ejemplar depositado en alguna colección científica perteneciente a alguna institución académica reconocida (museo o universidad) (GBIF); 2) el ejemplar debe haber sido identificado por algún especialista reconocido de dicho portal (Naturalista y Facebook); 3) en caso de ser fotografía, la especie debe poder ser determinada mediante caracteres visibles (patrón dorsal, coloración de las patas u ovisaco) (Naturalista y Facebook); 4) contar con datos completos de la localidad exacta donde el ejemplar fue observado (Facebook). Basado en los criterios anteriores, la base de datos obtenida del portal de Naturalista y Facebook cuenta únicamente con los registros de L. mactans y L. geometricus, mientras que de GBIF se descargaron los registros de las 3 especies para México. La georreferenciación se realizó con el programa Google Earth Pro v.7.3.2.5491 y el sistema de coordenadas geográficas WGS 84. Los registros georreferenciados fueron los pertenecientes a las colecciones científicas y literatura, así como los de Facebook que únicamente contaran con la localidad de recolecta pero sin coordenadas geográficas.
El modelaje de nicho ecológico (MNE) nos permiten crear una estimación de idoneidad (distribución potencial) estableciendo una relación entre las condiciones bióticas y abióticas donde se sabe que se distribuye una especie (Galante et al., 2018). Los análisis de distribución potencial se realizaron para todo el género (todos los registros de México), para L. mactans y L. geometricus con el programa Maxent v.3.4.1 (Phillips et al., 2006), utilizando un total de 21 capas ambientales con una resolución de 30 segundos de arco: 19 de WorldClim (https://www.worldclim.org/) (BIO1-BIO19) (Fick y Hijamas, 2017), y 2 de Conabio (http://www.conabio.gob.mx/informacion/gis/) (CON1, CON2) (Conabio, 2019) (Apéndice). Para determinar las variables ambientales con mayor aporte al MNE, en cada modelaje se realizó la ejecución de 10 réplicas estadísticas de bootstrap, en cada réplica se utilizó, de forma aleatoria, 60% de registros de entrenamiento y 40% de registros restantes como puntos de validación. Las configuraciones restantes del análisis se dejaron de manera predeterminada en el programa Maxent. Se llevó a cabo un segundo análisis únicamente con las capas ambientales de mayor aporte para cada modelaje y con la misma configuración que el primer MNE, discutiendo las 3 capas ambientales con mayor explicación para cada caso (Tabla 2). Los MNE obtenidos fueron evaluados por su valor de área bajo la curva: AUC (por sus siglas en inglés), que mide la capacidad de los modelos para discriminar positivos verdaderos y positivos falsos para el MNE (Illoldi-Rangel et al., 2008; Phillips et al., 2006; Phillips y Dudík, 2008). Este análisis está incluido en el programa MaxEnt (Phillips et al., 2006). Debido al escaso número de registros en México de L. hesperus (Tabla 1), no se realizó MNE para esta especie ya que los modelados con número pequeño de muestras son inexactos (Hanberry et al., 2012). Los mapas de distribución real y potencial fueron editados usando el programa Photoshop CS6 Extended.
Tabla 2 Variables ambientales de mayor aporte en el análisis de modelaje de nicho ecológico (MNE) para todo el género Latrodectus y cada especie en México.
| Análisis de modelaje de nicho ecológico | Capas ambientales de mayor aporte | Valor de aporte (%) |
|---|---|---|
| Latrodectus spp. AUC = 0.898 |
|
|
| L. mactans AUC = 0.928 |
|
|
| L. geometricus AUC = 0.933 |
|
|
Resultados
Se obtuvo un total de 1,147 registros de las 3 especies de Latrodectus de México: L. mactans, L. geometricus y L. hesperus (Tabla 1). Los registros que no fueron georreferenciados (73) y las localidades repetidas por especie (149), fueron eliminados para los análisis de MNE. Cuarenta y ocho registros fueron de ejemplares juveniles, por lo que la identificación se dejó hasta género (Latrodectus sp.) y se utilizaron únicamente para el MNE a nivel genérico (Tabla 3). La base de datos depurada contó con 925 registros, con la que se realizaron los mapas de distribución real y el MNE. Todos los estados de la República Mexicana y las 14 provincias biogeográficas presentaron registros de alguna especie de Latrodectus (Fig. 1). Las provincias biogeográficas que presentaron el mayor número de registros fueron el Desierto Chihuahuense (DC), Cinturón Volcánico Transmexicano (CVT) y la Cuenca del Balsas (CB) (Tabla 4; Fig. 1).
Tabla 3 Registros totales por especie del género Latrodectus y por estado en México.
| Estado | L. mactans | L. geometricus | L. hesperus | L. sp. | Registros totales por estado (%) |
|---|---|---|---|---|---|
| Aguascalientes | 10 | 10 | - | - | 20 (1.7) |
| Baja California | 9 | 22* | - | - | 31 (2.7) |
| Baja California Sur | 7 | 3 | 2 | - | 12 (1) |
| Campeche | 2 | 1** | - | - | 3 (0.2) |
| Chiapas | 5 | 5 | - | - | 10 (0.8) |
| Chihuahua | 8 | 7* | 2* | - | 17 (1.4) |
| Ciudad de México | 19 | - | - | 2 | 21 (1.8) |
| Coahuila | 6 | 4 | 2 | - | 12 (1) |
| Colima | 1 | 5 | - | - | 6 (0.5) |
| Durango | 6 | 6 | - | 2 | 14 (1.2) |
| Estado de México | 39 | 12 | - | 3 | 54 (4.7) |
| Guanajuato | 343 | 49 | - | - | 392 (34.1) |
| Guerrero | 25 | 18 | - | 5 | 48 (4.1) |
| Hidalgo | 15 | 3 | 1* | 2 | 21 (1.8) |
| Jalisco | 73 | 28 | - | - | 101 (8.8) |
| Michoacán | 7 | 15 | - | - | 22 (1.9) |
| Morelos | 67 | 17 | - | 6 | 90 (7.8) |
| Nayarit | 5 | 2 | - | 1 | 8 (0.6) |
| Nuevo León | 5 | 20 | - | - | 25 (2.1) |
| Oaxaca | 38 | 11 | - | 3 | 52 (4.5) |
| Puebla | 11 | 5 | - | 3 | 19 (1.6) |
| Querétaro | 3 | 13 | - | - | 16 (1.3) |
| Quintana Roo | 2 | 3 | - | - | 5 (0.4) |
| San Luis Potosí | 11 | 7 | - | 2 | 20 (1.7) |
| Sinaloa | 3 | 13 | - | - | 16 (1.3) |
| Sonora | 6 | 9* | - | - | 15 (1.3) |
| Tabasco | 1** | - | - | - | 1 (0.08) |
| Tamaulipas | 5 | 5 | - | - | 10 (0.8) |
| Tlaxcala | 31 | 5 | - | 16 | 52 (4.5) |
| Veracruz | 16 | 9 | - | 2 | 27 (2.3) |
| Yucatán | 1 | 2 | - | - | 3 (0.2) |
| Zacatecas | 2 | 1 | - | 1 | 4 (0.3) |
| Registros totales | 782 | 310 | 7 | 48 | 1,147 |
* = Registros nuevos para la entidad;
** = registros únicos de literatura que no pudieron ser georreferenciados.
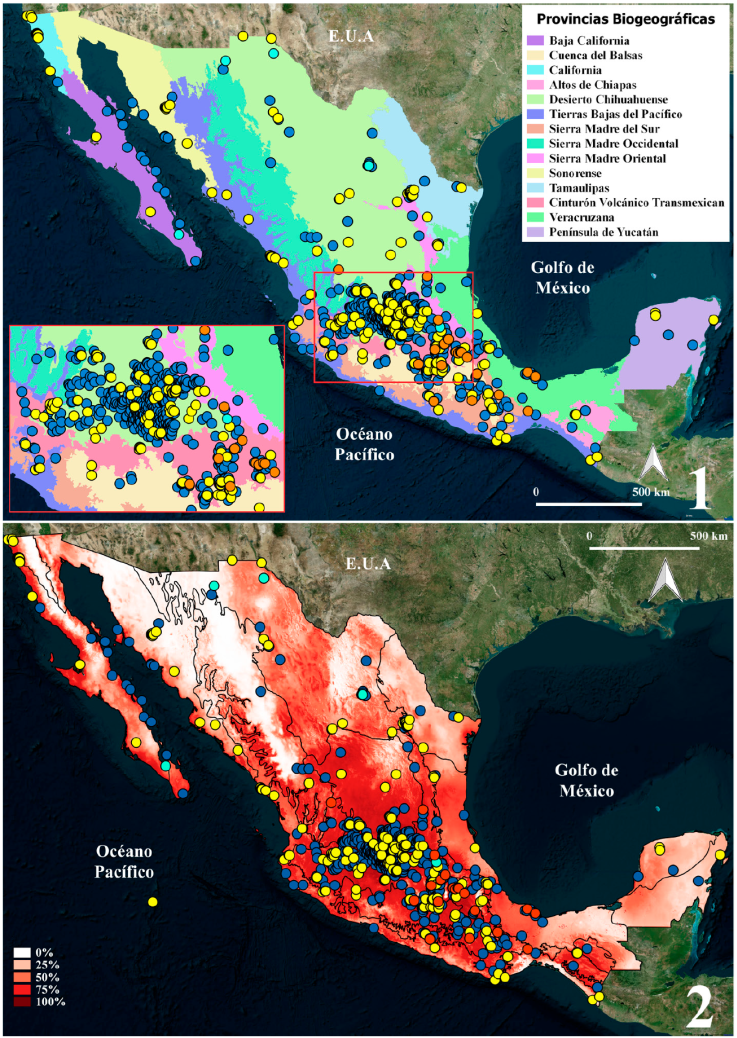
Figuras 1, 2 . Registros y distribución potencial de las 3 especies del género
Latrodectus de México: L.
geometricus (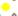 ),
L. hesperus (
),
L. hesperus (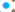 ), L. mactans (
), L. mactans ( ) y
Latrodectus sp. (
) y
Latrodectus sp. ( )
(ejemplares juveniles que no pudieron identificarse). 1) Registros
totales con base en las provincias biogeográficas propuestas por Morrone et al. (2017). 2) Modelaje
de nicho ecológico (MNE) de Latrodectus (todos los
registros) bajo el algoritmo de Maxent. Escala de colores: colores rojos
más oscuros muestran áreas con mejores condiciones de predicción
(0-100). Las líneas muestran las provincias biogeográficas.
)
(ejemplares juveniles que no pudieron identificarse). 1) Registros
totales con base en las provincias biogeográficas propuestas por Morrone et al. (2017). 2) Modelaje
de nicho ecológico (MNE) de Latrodectus (todos los
registros) bajo el algoritmo de Maxent. Escala de colores: colores rojos
más oscuros muestran áreas con mejores condiciones de predicción
(0-100). Las líneas muestran las provincias biogeográficas.
Tabla 4 Registros totales georreferenciados de las especies del género Latrodectus en México para cada región y provincias biogeográficas propuestas por Morrone et al. (2017).
| Región | Provincia biogeográfica | L. mactans | L. geometricus | L. hesperus | L. sp. | Total |
|---|---|---|---|---|---|---|
| Neártica | Desierto Chihuahuense (DC) | 381 | 100 | 4 | 1 | 486 |
| Baja California (BC) | 12 | 2 | 1 | - | 15 | |
| California (Cal) | 2 | 22 | - | - | 24 | |
| Sonorense (Son) | 6 | 9 | - | - | 15 | |
| Tamaulipas (Tam) | 3 | 21 | - | - | 24 | |
| Zona de transición mexicana | Sierra Madre Occidental (SMOc) | 12 | 7 | - | - | 19 |
| Sierra Madre Oriental (SMOr) | 41 | 4 | 1 | 2 | 48 | |
| Cinturón Volcánico Transmexicano (CVT) | 114 | 36 | - | 26 | 176 | |
| Cuenca del Balsas (CB) | 88 | 34 | - | 7 | 129 | |
| Sierra Madre del Sur (SMS) | 26 | 10 | - | 6 | 42 | |
| Neotropical | Tierras Bajas del Pacífico (TBP) | 22 | 40 | - | - | 62 |
| Veracruzana (Ver) | 17 | 6 | - | 2 | 25 | |
| Altos de Chiapas (AC) | 2 | - | - | - | 2 | |
| Península de Yucatán (PY) | 3 | 4 | - | - | 7 | |
| Totales | 729 | 295 | 6 | 44 | 1,074 |
El análisis de MNE para todos los registros del género Latrodectus con un valor de AUC de 0.898, mostró predicción en la mayor parte del país, modelando en todas las provincias biogeográficas (Fig. 2). Sin embargo, las provincias biogeográficas de la Sierra Madre Occidental (SMOc) y Sonorense (Son) mostraron una limitada predicción. La zona centro-sur de México es la región que presentó los valores más altos de predicción (Fig. 2). Las 3 capas ambientales con el mayor aporte fueron: tipo de vegetación y uso de suelo (CON2), temperatura estacional (BIO4) y precipitación del cuarto más frío (BIO19) (Tabla 2).
Latrodectus mactans: especie con la mayor distribución en México, con la mayoría de los registros en la región centro-sur del país (Figs. 1, 3; Tabla 3). Los estados que cuentan con la mayor cantidad de registros son Guanajuato, Jalisco, Morelos, Estado de México, Oaxaca y Tlaxcala (Tabla 3), mientras que los estados con el menor número de registros fueron Querétaro, Sinaloa, Campeche, Quintana Roo, Zacatecas, Colima, Tabasco y Yucatán (Tabla 3). Latrodectus mactans está reportada en las 14 provincias biogeográficas de México, siendo las provincias del DC y el CVT las que cuentan con el mayor número de registros (Fig. 3; Tabla 4). Respecto al MNE, con un valor de AUC de 0.928, las variables de temperatura estacional (BIO4), precipitación del cuarto más frío (BIO19) y tipo de vegetación, y uso de suelo (CON2) fueron las que tuvieron un mayor valor predictivo (Tabla 2). Las provincias biogeográficas con alta probabilidad son las correspondientes a la región centro del país, siendo las provincias del CVT, CB, SMOc y la zona sur del DC, (Fig. 3). Las provincias biogeográficas de la Sierra Madre Occidental, Sonorense, gran parte del DC, sur de la provincia Veracruzana y la Península de Yucatán (PY) mostraron una limitada o nula predicción (Fig. 2).

Figuras 3, 4 . Modelaje de nicho ecológico (MNE) bajo el algoritmo de Maxent. 3) MNE basado en los
registros ( ) de
L. mactans. 4) MNE basado en los registros
(
) de
L. mactans. 4) MNE basado en los registros
( ) de
L. geometricus. Escala de colores: colores rojos
más oscuros muestran áreas con mejores condiciones de predicción
(0-100). Las líneas muestran las provincias biogeográficas de acuerdo
con la clasificación de Morrone et al.
(2017).
) de
L. geometricus. Escala de colores: colores rojos
más oscuros muestran áreas con mejores condiciones de predicción
(0-100). Las líneas muestran las provincias biogeográficas de acuerdo
con la clasificación de Morrone et al.
(2017).
Latrodectus geometricus: esta especie presenta una amplia distribución en México con 310 registros (Tabla 3; Fig. 4). Las provincias que presentan el mayor número de registros hasta ahora fueron el DC, Tierras Bajas del Pacífico (TBP), CVT y la CB (Tabla 4). El valor de AUC para el MNE de L. geometricus fue de 0.933, siendo el tipo de vegetación y uso de suelo (CON2), rango de temperatura anual (BIO7) y la precipitación estacional (BIO15) las capas con el mayor aporte (Tabla 2). El programa Maxent estimó una amplia distribución en México para L. geometricus, con un valor de predicción mayor al 50% en la mayor parte del territorio mexicano (Fig. 4), principalmente hacia la parte centro-sur del país. De forma contraria, la predicción hacia las provincias de Baja California (BC), California (Cal), Son, la SMOc, mitad de la provincia del DC, provincia de Tamaulipas (Tam), parte sur de la provincia Ver y PY fue baja (Fig. 4).
Latrodectus hesperus: los registros de esta especie en México son recientes, para los estados de Baja California Sur y Coahuila (Jiménez et al. (2015) y Desales-Lara et al. (2018), respectivamente). El registro de Baja California por Jiménez et al. (2015) no cuenta con coordenadas, por lo que el de Coahuila, por Desales-Lara et al. (2018), es el primero bien documentado. Los registros de esta especie corresponden a la zona norte y centro del país y abarcan los estados de Baja California Sur, Coahuila, Chihuahua e Hidalgo (Fig. 1), con 9 registros en estos 2 últimos estados (Tabla 3). Hasta ahora, la provincia biogeográfica del DC cuenta con 4 de los 6 registros de L. hesperus en el país, mientras que los otros 2 pertenecen a las provincias de BC y SMOr (Tabla 4; Fig. 1). Cabe mencionar, que todos los registros se encontraron en pastizales y matorrales con clima semiárido a muy árido principalmente (Conabio, 2019; Espinosa et al., 2008; Morrone, 2005).
Variación geográfica de patrones de coloración dorsal del opistosoma. Se revisó el patrón dorsal de 384 ejemplares hembras adultas pertenecientes a las 3 especies, de 270 localidades distribuidas en 29 estados del país (Figs. 5-7). Trescientos cincuenta y dos ejemplares corresponden a L. mactans, 27 a L. geometricus y 5 a L. hesperus. En algunas localidades se encontró más de un tipo de patrón dorsal para una especie, y se identificaron 8 patrones dorsales de coloración para L. mactans (Figs. 5, 6, material suplementario: S1). Latrodectus geometricus presentó únicamente 3 patrones de coloración dorsal del opistosoma (Fig. 7, material suplementario: S2). Dada la poca cantidad de ejemplares de L. hesperus, fue difícil analizar variación, por lo que se reconocieron únicamente 2 patrones diferentes (Fig. 7; S2), similares a los patrones tipo 4 y tipo 6 de L. mactans.
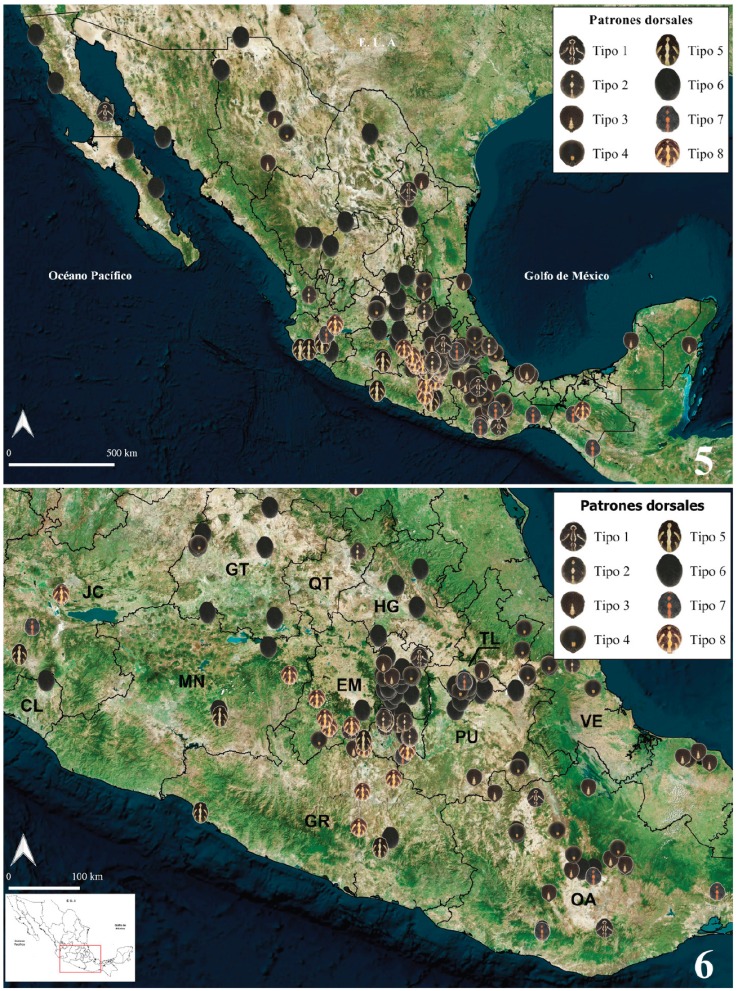
Figuras 5, 6 . 5) Registros de los 8 tipos de patrón dorsal del opistosoma de las hembras de L. mactans en México. 6) Patrones dorsales mostrados a detalle para la región central.
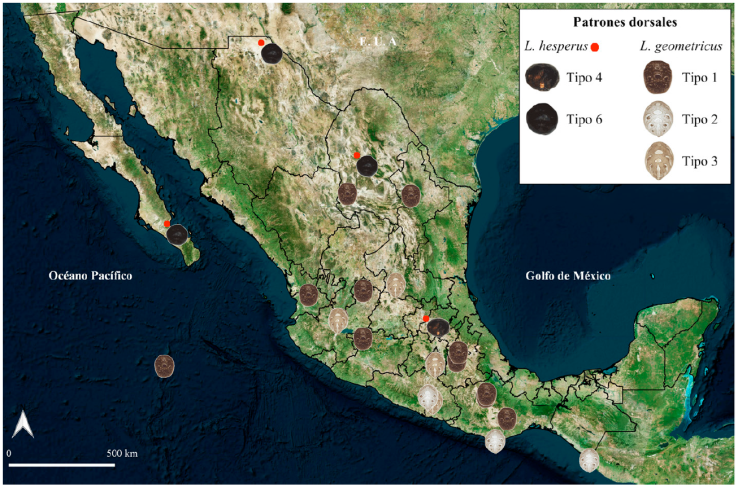
Figura 7 Registros de los tipos de patrón dorsal del opistosoma de las hembras de L. hesperus y L geometricus en México.
Levi (1959) analizó la variación de los patrones del opistosoma de L. mactans en el mundo, con 11 patrones dorsales diferentes para México. En el presente trabajo se encontraron 6 de los 11 patrones dorsales reportados por Levi (1959) y se reportan 2 nuevos para el país: tipos 1 y 3 (Tabla 5). El patrón tipo 6 fue el que se observó en el mayor número de ejemplares (144) y la mayor distribución geográfica (Tabla 5; Figs. 5, 6). Los estados con la mayor variación en los patrones dorsales de L. mactans fueron Guerrero y Morelos, con todos los tipos de patrón dorsal a excepción del tipo 4 (Tabla 5).
Tabla 5 Presencia (*)-ausencia por estado de los 8 tipos de patrones dorsales del opistosoma registrados en hembras de L. mactans en México. En negritas, estados que tienen el mayor número de patrones dorsales observados.
| Estado | Tipo 1 | Tipo 2 | Tipo 3 | Tipo 4 | Tipo 5 | Tipo 6 | Tipo 7 | Tipo 8 |
|---|---|---|---|---|---|---|---|---|
| Baja California | * | * | ||||||
| Baja California Sur | * | |||||||
| Campeche | * | |||||||
| Chiapas | * | * | ||||||
| Chihuahua | * | * | * | |||||
| Ciudad de México | * | * | ||||||
| Coahuila | * | |||||||
| Colima | * | |||||||
| Durango | * | |||||||
| Estado de México | * | * | * | * | * | |||
| Guanajuato | * | * | * | |||||
| Guerrero | * | * | * | * | * | * | * | |
| Hidalgo | * | |||||||
| Jalisco | * | * | * | |||||
| Michoacán | * | * | * | |||||
| Morelos | * | * | * | * | * | * | * | |
| Nayarit | ||||||||
| Nuevo León | * | * | * | * | ||||
| Oaxaca | * | * | * | * | * | * | * | |
| Puebla | * | * | * | |||||
| Querétaro | * | |||||||
| Quintana Roo | * | |||||||
| San Luis Potosí | * | * | * | |||||
| Sonora | * | |||||||
| Tamaulipas | * | |||||||
| Tlaxcala | * | * | * | * | ||||
| Veracruz | * | * | * | * | ||||
| Zacatecas | * |
Latrodectus geometricus presentó poca variación en los patrones dorsales del opistosoma (S2). Dicha variación se basó en 27 ejemplares de 24 localidades (Fig. 7). El más común para L. geometricus en México fue el tipo 1, presente en Aguascalientes, Coahuila, Colima, Jalisco, Michoacán, Morelos, Nayarit, Nuevo León, Oaxaca, Puebla y Tlaxcala. El patrón tipo 2 fue el de menor distribución, presente en Chiapas, Guerrero y Oaxaca (Fig. 7).
La variación del patrón dorsal de L. hesperus se observó en ejemplares de 4 estados de México, y se incluyeron los ejemplares de Coahuila (Fig. 7; Desales-Lara et al., 2018). El patrón más común fue el tipo 6, presente en los ejemplares de Baja California Sur, Chihuahua y Coahuila (Fig. 7). El patrón tipo 4 únicamente se encontró en el ejemplar de Hidalgo (Fig. 7).
Los machos de las 3 especies presentaron un patrón dorsal en el opistosoma similar (Figs. 8, 11, 14), la variación interespecífica radicó en la coloración general del opistosoma (Figs. 8-16). En L. mactans se presentan tonos de café obscuro a negro (Figs. 8-10), en L. hesperus tonalidades de café claro o crema (Figs. 11-13), mientras en L. geometricus tonalidades de café o gris (Figs. 14-16). Los machos de L. mactans, además, presentan tonos rojizos en la línea media del opistosoma (Figs. 8-10), lo cual no se observa en las otras 2 especies (Figs. 11-16).

Figuras 8-16 Variación de los patrones dorsales observados del opistosoma y coloración general de los machos de Latrodectus de México. 8-10) Latrodectus mactans. 11-12) Latrodectus hesperus. 14-16) Latrodectus geometricus.
Variación del patrón ventral “reloj de arena” del opistosoma. El patrón ventral del opistosoma fue revisado en las mismas 384 hembras en las que se analizó el patrón dorsal. De las 3 especies para México, el patrón de “reloj de arena” de L. mactans fue el que presentó la mayor variación, con 40 patrones diferentes, sin contar la ausencia total de éste (Fig. 17). Con estos resultados, a los 12 patrones ventrales reportados por Levi (1959) aumentan a 21 patrones. La ausencia del “reloj de arena” fue el patrón que más se presentó en el mayor número de ejemplares de L. mactans. En Latrodectus geometricus se observó dicho patrón siempre completo y poco variable, de color rojo o naranja, con un par de marcas pequeñas posteriores al “reloj de arena” cerca de las hileras (Figs. 18-20). En el caso de L. hesperus, con el limitado material revisado no se analizó dicha variación, encontrando tanto ejemplares con el patrón completo, como el patrón ausente.

Figuras 17-20 Variación observada de los tipos de patrones ventrales (40) del opistosoma en forma de “reloj de arena” en ejemplares hembra de México. 17) Latrodectus mactans. 18-20) Latrodectus geometricus, la coloración blanca es debido a que los ejemplares observados en alcohol etílico pierden sus tonalidades rojas o anaranjadas. Escalas: 2 mm.
Variación morfológica de caracteres taxonómicos. Las hembras de L. mactans y L. hesperus presentan un solapamiento de caracteres en estructuras sexuales y somáticas debido a su variabilidad (Figs. 21-26). Estas especies al pertenecer al clado mactans, presentan una morfología de estructuras sexuales internas (espermatecas y ductos copuladores) muy similares, con 4 o 5 vueltas del ducto copulador en el caso de L. mactans (Figs. 21-23), y 3 o 4 en de L. hesperus (Figs. 24-26).
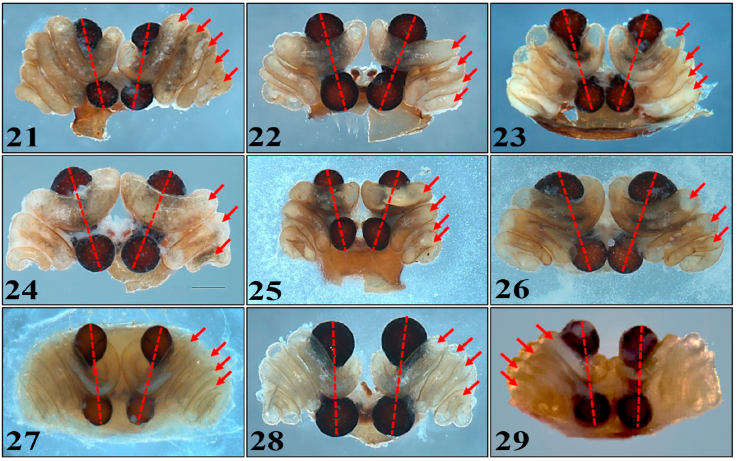
Figuras 21-29 Vista dorsal de los epiginios de hembras de las tres especies de arañas del género Latrodectus de México. 21-23) Latrodectus mactans. 24-26) Latrodectus hesperus. 27-29) Latrodectus geometricus. Flechas señalan las vueltas del ducto copulador. Líneas punteadas señalan el ángulo de las espermatecas entre sí. Escalas: 0.5mm.
Las hembras de L. geometricus presentan posición casi paralela entre las espermatecas (Figs. 27-29, líneas punteadas) y el número de vueltas del ducto copulador con menor variación (Figs. 27-29, flechas). Otro carácter importante para identificar a esta especie son las patas, con anillos de tonos obscuros en las uniones distales de todos los artejos, a excepción de la coxa y el trocánter, siendo la patela completamente oscura. La forma del ovisaco es distintivo de la especie, siendo circular y con proyecciones conspicuas en forma de espinas, característico de L. geometricus.
En el caso de los machos revisados, el número de vueltas del émbolo del pedipalpo presenta variación interespecífica entre las 3 especies, en L. mactans 3 vueltas (Figs. 30-33, línea punteada y flechas), en L. hesperus poco más de 2 vueltas (Figs. 34-37, línea punteada y flechas) y en L. geometricus poco más de 3 vueltas (Figs. 38-41).

Figuras 30-41 Comparación del número de vueltas del émbolo de los pedipalpos de machos de las 3 especies de arañas del género Latrodectus de México. 30-33) Latrodectus mactans. 34-37) Latrodectus hesperus. 38-41) Latrodectus geometricus. 34, 36) Tomado y modificado de Kaston (1970). 30, 32, 38, 40) Tomado y modificado de Levi (1959). Escalas: 0.5 mm. Flechas indican el número de vueltas del émbolo, las líneas discontinuas indican dónde comienza y dónde termina cada vuelta.
Discusión
A pesar de que el género Latrodectus cuenta con registros en todas las provincias biogeográficas de México, las provincias que contaron con el mayor número de registros georreferenciados abarcan también los estados con la mayor densidad poblacional del país (Tabla 4), tales como la Ciudad de México, Estado de México, Morelos, Tlaxcala, Aguascalientes, Guanajuato, Puebla, Querétaro, Hidalgo, Colima y Veracruz (INEGI, 2019; Figs. 1-4). Esta región se caracteriza por presentar solapamiento de elementos Neotropicales y Neárticos, por lo que se presentan diversos tipos de vegetación, predominando matorrales y bosque de pino-encino principalmente (Morrone, 2005).
Latrodectus mactans y L. hesperus se distribuyen de manera natural en zonas de pastizales, matorrales y bosque de pino-encino principalmente, sin embargo, han sido encontradas también en zonas de cultivo, pastoreo y en los alrededores e interior de las casas (Cabrera-Espinosa, 2020; Cabrera-Espinosa y Valdez-Mondragón, 2020). Las actividades humanas en la región centro del país, parecen un factor importante en la dispersión de estas arañas, por traslado de materiales donde se refugian, además de brindar el microhábitat ideal para su establecimiento y reproducción dada su alta capacidad sinantrópica (Cabrera-Espinosa, 2020; Cabrera-Espinosa y Valdez-Mondragón, 2020; Garb et al., 2004). Aunado a esto, las especies de Latrodectus también se dispersan mediante ballooning, que podría ser un factor importante que influye en su amplia distribución, como es el caso de L. mactans y L. geometricus (Johnson et al., 2015).
Latrodectus mactans ha sido recolectada en diferentes tipos de vegetación y clima en México, con un amplio gradiente altitudinal. En la zona centro del país, ejemplares han sido encontrados dentro y alrededor de casas, o en zonas perturbadas y conservadas de bosque de pino y pino-encino (obs. pers.). Esto está relacionado con el MNE de esta especie, con una alta probabilidad de distribución donde predominan los pastizales y bosques de pino (Fig. 3). En este análisis de MNE, la temperatura estacional (BIO4), fue la variable que más aportó en el modelaje de la especie (Tabla 2). Se ha observado en algunas especies de Latrodectus que la temperatura estacional es un factor importante para su reproducción y desarrollo, y que el calor extremo frena el crecimiento de las arañas en estadios juveniles y aumenta su mortalidad (Jonhson et al., 2019). Esto podría explicar el porqué de la baja predicción del MNE para la especie al norte del país (Fig. 3), donde las temperaturas en ciertas etapas del año suelen ser extremadamente cálidas, además de ser una región de baja densidad poblacional, lo que podría ser un factor importante que delimita su dispersión (Conabio, 2019; INEGI, 2019).
El análisis de MNE también presentó una baja predicción hacia el sureste de México (Fig. 3), que es la región con el mayor valor de evapotranspiración del país, con un clima de tipo cálido húmedo y una precipitación anual total promedio mayor a los 2,000 mm, con terrenos inundables y pantanos en algunas zonas como Tabasco, hábitat inadecuado para L. mactans (Conabio, 2019). Lo anterior hace suponer que estos registros, principalmente para el sur de la provincia de Ver y de PY podrían tratarse de arañas introducidas en zonas urbanas, y por ello una sobre predicción del MNE. Algo importante de mencionar, es que la mayoría de los registros de la especie son más numerosos hacia la parte central del país, lo cual puede influir en el MNE dada la presencia de más puntos georreferenciados.
Latrodectus geometricus, al ser una especie introducida y con rangos medioambientales más amplios que L. mactans, mostró una predicción mayor en el país (Fig. 4). La importancia de la variable ambiental del tipo de vegetación y uso de suelo (CON2) en el análisis de MNE (Tabla 2), nos habla claramente de dependencia a los ambientes antropizados al ser una especie introducida, encontrada principalmente en los alrededores de las casas y ambientes urbanos, con mayor densidad poblacional como es la región central del país (Conabio, 2019; Fig. 4). En recientes expediciones y observaciones de campo por parte del LATLAX a diferentes estados del país (Tabla 3), permite suponer que esta especie prefiere climas cálidos de bajas altitudes, por lo que su presencia en climas templados con altitudes mayores a los 2,000 m podría deberse principalmente a dependencia a hábitats proporcionado por el hombre. Al ser una especie dependiente de ambientes antropizados, la baja predicción del MNE de L. geometricus hacia el sureste de México, sugiere que las altas condiciones de humedad y temperatura, aunado a altas precipitaciones probablemente sean factores importantes que limitan la distribución en esos ambientes naturales (Fig. 4). Debido a ésto, los registros de L. geometricus en la provincia de PY deben de provenir de ejemplares encontrados en zonas urbanas o antropizadas. Se ha observado que al ser una especie introducida y de fácil aclimatación, ha provocado el desplazamiento de poblaciones nativas de algunas especies como L. hesperus (Cabrera-Espinosa y Valdez-Mondragón, 2019; Vetter et al., 2012) y poblaciones de L. mactans (Dr. Ponce Saavedra, com. pers.)
A pesar de que actualmente se cuenta con pocos registros de L. hesperus en México, esta distribución concuerda con la mencionada en la descripción original de la especie realizada por Chamberlin e Ivie (1935), principalmente al norte de México, ya que lo que menciona hacia al sur del país corresponde a L. mactans (Figs. 1, 2). Todos los ejemplares han sido recolectados asociados a matorral xerófilo y pastizales (Desales-Lara et al., 2018; Jiménez et al., 2015). La provincia del DC (con el mayor número de registros), se caracteriza por presentar clima semiárido, árido y muy árido, con una precipitación anual total de 500 mm, lo que favorece la presencia de matorrales xerófilos y pastizales (Conabio, 2019; Espinosa et al., 2008; Morrone, 2005). Los ejemplares recolectados en Baja California Sur e Hidalgo fueron encontrados en una zona árida de clima seco y vegetación de matorral. Podemos inferir que esta especie en ambientes silvestres prefiere hábitats secos y áridos. Estas condiciones ambientales y tipo de vegetación donde existen registros de L. hesperus hasta ahora, es el tipo de vegetación en la que el MNE de L. mactans no presenta predicción (Fig. 3), por lo que aparentemente no hay un solapamiento de distribución entre estas 2 especies en México. Sin embargo, se requiere un mayor número de registros de L. hesperus para poderlo analizar de manera más robusta y corroborarlo.
Como lo menciona Kaston (1970), especies como L. hesperus, L. mactans y L. variolus, tanto machos como hembras a nivel intraespecífico, pueden requerir diferente número de mudas para alcanzar la madurez sexual. Con cada muda, van adquiriendo tonalidades más obscuras en su coloración, por lo que un ejemplar que tardará más mudas que el promedio en alcanzar la madurez sexual, tendrá patrones de coloración rojizo en el opistosoma más pequeños que el resto de los ejemplares de su especie. Sin embargo, durante la revisión del material de las colecciones, se observó que, en algunos casos, los juveniles ya presentan el mismo patrón dorsal en el opistosoma que los adultos. En el caso de algunos patrones dorsales con distribución geográfica más regionalizada (ej. tipos 5 y 8 de L. mactans; Figs. 5, 6), existe la posibilidad de que se trataran de especies diferentes, aunque estudios taxonómicos más profundos utilizando evidencia molecular son necesarios para ser conclusivos. Este supuesto se apoya en que hace más de 60 años, Levi (1959) reporta el mismo patrón dorsal para ejemplares de la misma región de la costa del Pacífico, indicando que es un patrón conservado en ciertas poblaciones hacia el occidente de México (Figs. 5, 6).
A pesar de ser patrones de coloración poco variables, se observó que los 3 patrones dorsales de L. geometricus pueden variar en la tonalidad general del opistosoma. Esta variación de tonalidad es constante en las poblaciones, por lo que podría estar influida por factores ambientales o por la alimentación como se ha observado en otras arañas (Vetter, 2015). Keegan (1955) menciona que, a pesar de la amplia variación de coloración, las diferencias en los patrones del opistosoma en Latrodectus pueden ser excelentes criterios para la separación de especies. Como en otras arañas, los patrones dorsales y laterales suelen coincidir con puntos de inserción de la musculatura interna, como las especies de coloraciones más claras como el caso de L. geometricus o Latrodectus pallidus O. Pickard Cambridge, 1872, donde incluso no hay solapamiento de patrones. Sin embargo, ésto no pasa para las especies más oscuras como por ejemplo L. mactans o Latrodectus hasselti Thorell, 1870, donde las poblaciones provenientes de áreas geográficas ampliamente separadas presentan amplia variación en patrones de coloración, que quizá podría hablar de especies distintas (Keegan, 1955).
El patrón ventral del opistosoma en forma de “reloj de arena” rojo es un rasgo característico que permite la fácil identificación del género Latrodectus en México, sirviendo antiguamente en su taxonomía, como un carácter de identificación a nivel de especie y subespecie (Chamberlin e Ivie, 1935; Kaston, 1970; Levi, 1959). La amplia variación del patrón ventral del opistosoma del L. mactans no parece estar relacionada con la variación en los patrones dorsales. En los ejemplares recolectados en las grutas de Tolantongo, Hidalgo, por ejemplo; todos los ejemplares presentaban el mismo patrón dorsal, pero diferentes patrones ventrales, encontrando ejemplares con el reloj completo, partido, sólo una parte o completamente ausente (Fig. 17). Esto nos habla de la gran plasticidad y variación de este carácter, incluso en ejemplares de una misma población. En el caso de L. hesperus, Chamberlin e Ivie (1935) mencionan que esta especie presenta una amplia variedad de patrones ventrales, siendo la forma de reloj de arena completo el más común. Tanto la variación en los patrones dorsales de coloración del opistosoma como en los patrones ventrales del “reloj de arena”, fueron revisados solo en hembras, ya que presentaron una mayor variación que en machos (Figs. 8-16).
Se considera que las estructuras sexuales de las arañas (espermatecas en las hembras y pedipalpos en los machos) son caracteres morfológicos robustos, que permiten la identificación a nivel de especie en la mayoría de los grupos debido a su evolución más rápida en comparación con estructuras somáticas (Huber, 2003; Huber et al., 2005). Sin embargo, estos caracteres parecen no ser tan robustos en algunas especies del género Latrodectus, ya que tanto L. mactans como L. hesperus presentaron un solapamiento en el número de vueltas del ducto copulador en hembras (Cabrera-Espinosa, 2020; Levi, 1958) (Figs. 21-26). Lo anterior presenta un problema taxonómico, dificultado la identificación de los ejemplares de L. hesperus en México. Chamberlin e Ivie (1935) describen el epiginio de las hembras de L. hesperus con 4 vueltas del ducto copulador (Fig. 25), similar al de L. mactans (Fig. 22). Posteriormente, Levy y Amitai (1983) reportaron en Israel a L. hesperus (especie introducida) con 3 vueltas en el ducto copulador, lo que coincide con ejemplares recolectados por Desales-Lara et al. (2018) en Coahuila (Fig. 26) y en Hidalgo (Fig. 24).
En el caso de los machos, Kaston (1970) indica que el émbolo del pedipalpo de L. mactans presenta poco más de 3 vueltas (Figs. 30, 32) y el pedipalpo del macho tipo de L. hesperus, únicamente 2 vueltas (Figs. 34, 36), mientras que el de L. geometricus tiene casi 4 vueltas (Figs. 38, 40). Sin embargo, este carácter parece ser variable incluso dentro de una misma especie, en L. geometricus puede variar de 3 a casi 4 vueltas (Figs. 40, 41). De esta manera, el número de vueltas del émbolo debe de ser considerado con precaución para identificación a nivel de especie. El uso de caracteres somáticos también puede aportar información relevante en conjunto con los caracteres sexuales para la identificación de especies, incluso caracteres ultra morfológicos (sedas del opistosoma) han sido caracteres útiles, como se observó en L. umbukwane (Wright et al., 2019).
Latrodectus mactans es la especie con la mayor variación en las estructuras sexuales y somáticas dada una supuesta amplia distribución, además de la aparente regionalización en México de algunos de los patrones dorsales del opistosoma de las hembras. Esto nos habla de que posiblemente se trate de un complejo de especies, por lo que la diversidad del género en el país podría estar subestimada. Actualmente, el uso de una taxonomía integradora ha ayudado en la delimitación de complejos de especies en arañas, al usar de diferentes evidencias (ecológicas, biológicas, ADN, conducta, biogeografía, etc.) en conjunto con diferentes técnicas y métodos de delimitación de linajes (Hamilton et al., 2014; Navarro-Rodríguez y Valdez-Mondragón, 2020; Padial et al., 2010; Valdez-Mondragón, 2020; Valdez-Mondragón et al., 2019). Este es el caso de Latrodectus thoracicus Nicolet, 1849, dicha especie fue revalidada después de un estudio con caracteres morfológicos y moleculares, utilizando el gen mitocondrial CO1 (Aguilera et al., 2009). Latrodectus umbukwane de África es última especie descrita del género hasta el momento por Wright et al. (2019), utilizando caracteres morfológicos, ultra morfológicos y moleculares (CO1) como evidencias para su descripción. Esto nos habla que no solamente en México, sino quizá en otras regiones del mundo, la diversidad de especies de Latrodectus podría ser mayor a la conocida actualmente.
A pesar del amplio conocimiento que se tiene del género Latrodectus en el mundo y en Norte América y ser arañas de importancia médica, los aspectos biológicos y taxonómicos siguen siendo poco estudiados, y las poblaciones/especies de México no son la excepción. Es necesario abordar estudios a fondo desde distintos aspectos con este grupo de arañas, como biogeográficos, de su historia natural, ecológicos y, sobre todo, taxonómicos en un contexto integrador; ésto aportaría información valiosa para conocer más sobre el género y la posible diversidad subestimada en el país.

