











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
The family Asteraceae has one of its main diversification centers in Mexico and constitutes the family with the largest number of species in the country (Villaseñor, 1990, 2016, 2018). It shows a great ecological tolerance, since its species are found from the coastal vegetation to the high mountains (Ortiz-Bermudez et al., 1998; Villaseñor, 2018; Villaseñor et al., 1998). The family has an important center of diversification in central Mexico, with an important number of endemic species (1,988; Villaseñor, 2018). This region lies basically in 2 biogeographic provinces (Fig. 1), the Balsas Basin (BB) and the Transmexican Volcanic Belt (TVB) (Morrone, 2006, 2014a, 2019; Morrone et al., 2017).

Figure 1 Collecting localities of 151 species of Asteraceae endemic to the BB (orange) and TVB (blue). The study area was divided in grid-cells of 30’ latitude and longitude (A) and according to the hydrological basins (B).
The BB is located between the TVB and the Sierra Madre del Sur, mainly in the states of Guerrero, Jalisco, Estado de México, Michoacán, Morelos, Oaxaca, and Puebla, at altitudes below 1,600 m (Morrone, 2006, 2019). Fernández-Nava et al. (1998) reported a total of 4,442 vascular plants species for this province, indicating that 537 are Asteraceae, 48 of them endemic to the province (Rodríguez-Jiménez et al., 2005). The TVB is located northwards of the BB and covers part of the states of Aguascalientes, Guanajuato, Jalisco, Mexico City, Michoacán, Oaxaca, Puebla, Estado de México, Tlaxcala, and Veracruz, being defined by altitudes above 1,600-1,800 m (Morrone, 2006, 2019). For this province, 5,229 species of vascular plants have been recorded (Bonilla-Barbosa & Villaseñor, 2003), 709 corresponding to Asteraceae and 20 of them being endemic to the province (Villaseñor & Ortiz, 2007).
One way to understand the distributional patterns of the species of Asteraceae is through the identification of their areas of endemism. An area of endemism is a non-random region of congruent distribution of 2 or more species or supra-specific taxa (Morrone, 1994). Although several methods have been proposed for the identification of areas of endemism (Morrone, 2007), 2 stand out as the most robust approaches (Munguía-Lino et al., 2016): Parsimony Analysis of Endemicity (PAE; Morrone, 1994, 2014b; Rosen, 1988) and Endemicity Analysis (EA; Szumik & Goloboff, 2004; Szumik et al., 2002). PAE has been frequently used to evaluate biogeographic relationships and to identify areas of endemism and generalized tracks, whereas EA has been used explicitly for the recognition of areas of endemism (Morrone, 2014a; Munguía-Lino et al., 2016). Several discussions have arisen on which of these methods is better for the identification of areas of endemism; however, Escalante et al. (2013) consider that both methods offer different advantages, and their results can be complementary. EA can recognize areas of endemism and PAE allows to organize them hierarchically. Our objective was to identify areas of endemism within the BB and TVB based on the species of Asteraceae endemic to these provinces. It was expected that these areas of endemism provide relevant information on the biogeographic history of the region and for future studies aimed at the conservation of their floristic richness.
Materials and methods
The study area corresponds to the biogeographic provinces BB and TVB (Morrone, 2006, 2014, 2019; Morrone et al., 2017). The BB is located between the TVB and the Sierra Madre del Sur, and it is characterized especially by the occurrence of seasonally dry tropical forests at altitudes below 1,800 m (Morrone, 2006). The TVB is located towards the center of the country, being characterized by the occurrence of temperate forests at altitudes above 1,800 m (Morrone, 2006).
To carry out the analyzes, the aforementioned provinces were divided first into units of defined size with the same surface (grid-cells), and later according to the hydrological sub-basins (Conabio, 1998) in which these provinces are encompassed. The sub-basins were used in order to know if the Asteraceae endemism patterns are maintained in the geographic space, regardless of the size or scale of areas used in the analysis. The use of hydrological sub-basins allows to circumscribe more naturally the distribution of endemic Asteraceae species to the BB and TVB provinces, constituting predefined areas of endemism that allow determining the relationships between the areas (Morrone, 2014b).
In this way, 216 grid-cells of 30’ latitude and longitude were obtained (with an approximate area of 3,000 km2), of which only 53 grids-cells contained records of endemic species of Asteraceae. In addition, both provinces correspond to 33 hydrological sub-basins, 27 of them with information about endemic species (Fig. 1, Table 1). The grid-cell size used was determined following the strategy of Suárez-Mota et al. (2013), based on IUCN recommendations (IUCN, 2012), and representing about 10% of the total distance between the extreme points of the total surface under study.
Table 1 Hydrological sub-basins occurring within of the BB and the TVB.
| Identifier | Hydrological subbasin |
|---|---|
| 1 | La Vega-Cocula |
| 2 | Lago Chapala |
| 3 | Lago de Pátzcuaro-Cuitzeo y Lago de Yuriria |
| 4 | Río Ameca-Atenguillo |
| 5 | Río Ameca-Ixtapa |
| 6 | Río Armería |
| 7 | Río Atoyac-A |
| 8 | Río Atoyac-B |
| 9 | Río Balsas-Infiernillo |
| 10 | Río Balsas-Mezcala |
| 11 | Río Balsas-Zirándalo |
| 12 | Río Coahuayana |
| 13 | Río Cutzamala |
| 14 | Río Grande de Amacuzac |
| 15 | Río Huicicila-San Blas |
| 16 | Río Ixtapa y otros |
| 17 | Río Jamapa y otros |
| 18 | Río Lerma-Chapala |
| 19 | Río Lerma-Salamanca |
| 20 | Río Lerma-Toluca |
| 21 | Río Moctezuma |
| 22 | Río Nautla y otros |
| 23 | Río Nexpe y otros |
| 24 | Río Papagayo |
| 25 | Río Papaloapan |
| 26 | Río Santiago Guadalajara |
| 27 | Río Tacámbaro |
| 28 | Río Tecolutla |
| 29 | Río Tehuantepec |
| 30 | Río Tepalcatepec |
| 31 | Río Tepalcatepec-Infiernillo |
| 32 | Río Tlapaneco |
| 33 | Santiago-Aguamilpa |
We analyzed the distribution data of 148 species of Asteraceae endemic both to BB and TVB. Of these species, 79 are distributed only in TVB, 39 occur only in BB and 30 are shared between both provinces (Table 2). To carry out the analyses, the collection records occurring within a maximum distance of 2 contiguous cells (approximately 111 km), out of the study biogeographic provinces limits were considered, with the aim of including the full records of the endemic species in order to fulfill their complete distribution. The information about the species and their collection sites come from the personal database of one of the authors (JLV), although most of the records are available in public data bases, such as GBIF (http://www.gbif.org), UNIBIO of the Instituto de Biología, UNAM (http://www.unibio.unam.mx), and SNIB of the Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio, http://www.conabio.gob.mx). However, the database and matrices with which this study was carried out will be available from the authors upon request. In total, 901 unique records were evaluated, to which geographic coordinates were assigned when the records lacked them, in addition to corroborating that the georeferenced records were within the biogeographic provinces analyzed or no more than 2 cells away from them.
Table 2 Species of Asteraceae endemic to the BB and the TVB biogeographic provinces analyzed in this study.
| Species* | Endemic | ||
|---|---|---|---|
| BB | TVB | ||
| 1 | Acourtia lepidopoda (B.L. Rob.) Reveal & R.M. King | X | X |
| 2 | Acourtia michoacana (B.L. Rob.) Reveal & R.M. King | X | X |
| 3 | Acourtia pringlei (B.L. Rob. & Greenm.) Reveal & R.M. King | X | |
| 4 | Ageratina arsenei (B.L. Rob.) R.M. King & H. Rob. | X | |
| 5 | Ageratina colimana B.L. Turner | X | |
| 6 | Ageratina enixa (B.L. Rob.) R.M. King & H. Rob. | X | |
| 7 | Ageratina iltisii B.L. Turner | X | |
| 8 | Ageratina jocotepecana B.L. Turner | X | |
| 9 | Ageratina lasia (B.L. Rob.) R.M. King & H. Rob. | X | X |
| 10 | Ageratina moorei B.L. Turner | X | |
| 11 | Ageratina neohintoniorum B.L. Turner | X | X |
| 12 | Ageratina pendula Panero & Villaseñor | X | |
| 13 | Ageratina perezii B.L. Turner | X | |
| 14 | Ageratina photina (B.L. Rob.) R.M. King & H. Rob. | X | X |
| 15 | Ageratina spooneri B.L. Turner | X | |
| 16 | Ageratum platypodum B.L. Rob. | X | |
| 17 | Aldama subcanescens (S.F. Blake) E.E. Schill. & Panero | X | |
| 18 | Alloispermum longiradiatum (Urbatsch & B.L. Turner) B.L. Turner | X | |
| 19 | Archibaccharis simplex S.F. Blake | X | |
| 20 | Archibaccharis veracruzana G.L. Nesom | X | |
| 21 | Astranthium laetificum De Jong | X | |
| 22 | Astranthium reichei Rzed. | X | |
| 23 | Baccharis erosoricola Rzed. | X | |
| 24 | Baccharis macrocephala Sch. Bip. ex Greenm. | X | |
| 25 | Bidens colimana Melchert | X | |
| 26 | Bidens gracillima Sherff | X | |
| 27 | Bidens gypsophila Miranda | X | |
| 28 | Bidens hintonii (Sherff) Melchert | X | |
| 29 | Bidens ocellatus (Greenm.) Melchert | X | X |
| 30 | Bidens pringlei Greenm. | X | X |
| 31 | Bidens purpusorum Bitter & Petersen | X | |
| 32 | Brickellia huahuapana B.L. Turner | X | |
| 33 | Brickellia jimenezii Hinojosa & Cruz-Durán | X | |
| 34 | Brickellia leonis Rzed. & Calderón | X | |
| 35 | Brickellia serboana B.L. Turner | X | |
| 36 | Carminatia papagayana B.L. Turner | X | |
| 37 | Cirsium cernuum Lag. | X | |
| 38 | Cirsium jorullense (Kunth) Spreng. | X | |
| 39 | Cirsium liebmannii (Sch. Bip. ex Klatt) Petr. | X | |
| 40 | Cirsium lomatolepis (Hemsl.) Petr. | X | |
| 41 | Cirsium nivale (Kunth) Sch. Bip. | X | |
| 42 | Cirsium pascuarense (Kunth) Spreng. | X | |
| 43 | Coreocarpus ixtapanus B.L. Turner | X | X |
| 44 | Cosmos deficiens (Sherff) Melchert | X | |
| 45 | Cosmos jaliscensis Sherff | X | |
| 46 | Cosmos landii Sherff | X | |
| 47 | Cosmos longipetiolatus Melchert | X | |
| 48 | Cosmos nitidus Paray | X | |
| 49 | Cosmos pseudoperfoliatus Art. Castro, Harker & Aarón Rodr. | X | |
| 50 | Cosmos ramirezianus Art. Castro, Harker & Aarón Rodr. | X | |
| 51 | Chaptalia hintonii Bullock | X | X |
| 52 | Chionolaena lavandulifolia (Kunth) Benth. & Hook. f. ex B.D. Jacks. | X | |
| 53 | Chrysanthellum involutum Paul G. Wilson | X | |
| 54 | Dahlia barkeriae Knowles & Westc. | X | |
| 55 | Dahlia brevis P.D. Sorensen | X | |
| 56 | Dahlia parvibracteata Saar & P.D. Sorensen | X | X |
| 57 | Dahlia rudis P.D. Sorensen | X | |
| 58 | Davilanthus huajuapanus (Panero & Villaseñor) E.E. Schill. & Panero | X | |
| 59 | Davilanthus veracruzanus B.L. Turner | X | |
| 60 | Dendroviguiera guerrerana (Panero & E.E. X Schill.) E.E. Schill. & Panero | X | |
| 61 | Dendroviguiera mirandae (Panero & E.E. Schill.) E.E. Schill. & Panero | X | X |
| 62 | Dendroviguiera splendens (Panero & E.E. Schill.) E.E. Schill. & Panero | X | X |
| 63 | Electranthera mutica (DC.) Mesfin, D.J. Crawford & Pruski | X | |
| 64 | Eremosis solorzanoana (Rzed. & Calderón) Pruski | X | |
| 65 | Erigeron annuactis G.L. Nesom | X | |
| 66 | Erigeron morelensis Greenm. | X | |
| 67 | Eryngiophyllum pinnatisectum Paul G. Wilson | X | |
| 68 | Flaveria kochiana B.L. Turner | X | |
| 69 | Grindelia nelsonii Steyerm. | X | |
| 70 | Grindelia sublanuginosa Steyerm. | X | |
| 71 | Guardiola pappifera Paul G. Wilson | X | |
| 72 | Guardiola thompsonii Van Faasen | X | X |
| 73 | Gutierrezia dunalii (Speng.) G.L. Nesom | X | X |
| 74 | Heliopsis brachactis Standl. ex T.R. Fisher | X | |
| 75 | Hymenostephium woronowii (S.F. Blake) E.E. Schill. & Panero | X | X |
| 76 | Hymenothrix greenmanii (Heiser) B.L. Turner | X | |
| 77 | Jaegeria robustior Rzed. | X | X |
| 78 | Jaegeria sterilis McVaugh | X | |
| 79 | Lagascea heteropappus Hemsl. | X | X |
| 80 | Lasianthaea squarrosa (Greenm.) K.M. Becker | X | |
| 81 | Leiboldia guerreroana (S.B. Jones) H. Rob. | X | |
| 82 | Liabellum hintoniorum (B.L. Turner) H. Rob. | X | |
| 83 | Mexerion sarmentosum (Klatt) G.L. Nesom | X | X |
| 84 | Microspermum debile Benth. | X | |
| 85 | Microspermum flaccidum Paul G. Wilson | X | X |
| 86 | Nelsonianthus tapianus (B.L. Turner) C. Jeffrey | X | |
| 87 | Neurolaena balsana B.L. Turner | X | |
| 88 | Oritrophium orizabense G.L. Nesom | X | |
| 89 | Pectis exilis D.J. Keil | X | |
| 90 | Pectis holochaeta (S.F. Blake) D.J. Keil | X | |
| 91 | Pectis leavenworthii Standl. | X | |
| 92 | Perymenium episcopale J.J. Fay | X | |
| 93 | Perymenium garciaruizii Rzed. | X | |
| 94 | Perymenium ibarrarum Rzed. & Calderón | X | |
| 95 | Perymenium paneroi B.L. Turner | X | |
| 96 | Perymenium reticulatum J.J. Fay | X | |
| 97 | Perymenium rogmacvaughii Rzed. & Calderón | X | |
| 98 | Philactis fayi A.M. Torres | X | |
| 99 | Piqueria glandulosa B.L. Turner | X | X |
| 100 | Porophyllum warnockii R.R. Johnson | X | X |
| 101 | Psacalium hintonii (Pippen) H. Rob. & Brettell | X | |
| 102 | Psacalium matudae H. Rob. & Brettell | X | X |
| 103 | Psacalium napellifolium (Schauer) H. Rob. & Brettell | X | |
| 104 | Pseudognaphalium altamiranum (Greenm.) Anderb. | X | |
| 105 | Pseudognaphalium oxyphyllum (DC.) Kirp. | X | |
| 106 | Roldana glinophylla H. Rob. & Brettell | X | |
| 107 | Roldana hederifolia (Hemsl.) H. Rob. & Brettell | X | X |
| 108 | Roldana hintonii H. Rob. & Brettell | X | |
| 109 | Rumfordia revealii H. Rob. | X | |
| 110 | Selloa plantaginea Kunth | X | |
| 111 | Senecio mulgediifolius S. Schauer | X | |
| 112 | Senecio orizabensis Sch. Bip. ex Hemsl. | X | |
| 113 | Senecio procumbens Kunth | X | |
| 114 | Senecio roseus Sch. Bip. | X | |
| 115 | Simsia tenuis (Fernald) S.F. Blake | X | |
| 116 | Stevia cordifolia Benth. | X | |
| 117 | Stevia hintonii (Grashoff) B.L. Turner | X | |
| 118 | Stevia hypomalaca B.L. Rob. | X | X |
| 119 | Stevia macvaughii Grashoff | X | |
| 120 | Stevia mascotensis Soejima & Yahara | X | |
| 121 | Stevia mitopoda B.L. Rob. | X | |
| 122 | Stevia totalcoana B.L. Turner | X | |
| 123 | Stevia vernicosa Greenm. | X | |
| 124 | Tridax trilobata (Cav.) Hemsl. | X | X |
| 125 | Trigonospermum alexandri Rzed., Calderón& Pérez-Calix | X | |
| 126 | Trigonospermum auriculatum B.L. Turner | X | |
| 127 | Trixis alata D. Don | X | |
| 128 | Verbesina calzadae Panero & Villaseñor | X | |
| 129 | Verbesina cuautlensis McVaugh | X | |
| 130 | Verbesina fusiformis McVaugh | X | |
| 131 | Verbesina juxtlahuacensis Panero & Villaseñor | X | |
| 132 | Verbesina linearis (McVaugh) B.L. Turner | X | |
| 133 | Verbesina machucana B.L. Turner | X | |
| 134 | Verbesina paneroi B.L. Turner | X | |
| 135 | Verbesina pietatis McVaugh | X | X |
| 136 | Verbesina pterocaula DC. | X | X |
| 137 | Verbesina reyesii Panero & Villaseñor | X | |
| 138 | Verbesina seatonii S.F. Blake | X | |
| 139 | Verbesina stenophylla Greenm. | X | X |
| 140 | Verbesina villasenorii B.L. Turner | X | |
| 141 | Verbesina xicoana B.L. Turner | X | |
| 142 | Viguiera moreliana B.L. Turner | X | |
| 143 | Viguiera sultepecana Paray | X | |
| 144 | Viguiera tepoxtlensis Paray | X | X |
| 145 | Villanova achillaeoides (Less.) Less. | X | |
| 146 | Wedelia cordiformis McVaugh | X | |
| 147 | Wedelia keilii B.L. Turner | X | X |
| 148 | Wedelia simsioides McVaugh | X | |
* The database and matrices with which this table was built will be available from the authors upon request.
The EA was performed using NDM/VNDM version 3.0 (Szumik & Goloboff, 2004), which uses an optimality criterion and calculates an endemicity index (E), that takes into account the number of endemics and the distributional restriction of the taxa in a given area. In this way, areas with a higher index value are postulated as areas of endemism (Escalante, 2009, 2015; Szumik & Goloboff, 2004; Szumik et al., 2002). Individually, a taxon will have a maximum e value (= 1) if it is present in all the grid-cells evaluated; on the contrary, if the taxon is found only in 1 or few of the evaluated cells, the e value will be lower. The value of E depends on the number of taxa included in the study area; that is, the more taxa that are considered endemic (high e values), the greater the value of E (Noguera-Urbano & Escalante, 2015; Szumik & Goloboff, 2004). EA requires that the study area is divided into grids previously defined by the user. In this way, the areas (sets of grid-cells) that are congruent with the distribution of as many species as possible are evaluated (Munguía-Lino et al., 2016). The surface of BB and TVB, as well as the surface of the hydrological sub-basins that include these provinces, were divided into grid-cells of 30’ latitude and longitude (Fig. 1). In summary, NDM/VNDM generated 216 cells for both provinces and 242 cells considering hydrological sub-basins.
NDM/VNDM also requires a list of species with their respective geographic coordinates. Such a list is automatically transformed into a matrix of presences (1) and absences (0) with respect to each of the grid-cells of the study area (Szumik & Goloboff, 2004; Szumik et al., 2002). The configuration of the program was carried out following the recommendations of Munguía-Lino et al. (2016). The areas of endemism were selected using the search ran 1,000 times and keeping overlapping as subsets when 98% of species superposition was recorded as unique; in addition, the edge effect was activated. From the subsets of areas obtained, those species with a minimum score of 0.6 were chosen and the consensus of these selected areas were calculated considering 30% species similarity under a strict consensus.
To carry out the PAE, a matrix of 151 species (columns) by 53 grid-cells (rows) was generated. For the analysis using hydrological sub-basins a matrix of 148 species (columns) by 27 sub-basins (rows) was generated. Once the matrices were prepared, they were introduced into the TNT (Goloboff & Catalano, 2016) program to generate the PAE, instructing the program to carry out the analysis through a Wagner network, with a value of 10 in the random seed and to perform 100 replicas. The rest of the parameters were assigned by default. The cladograms obtained with the TNT program were saved and a strict consensus visualized with Winclada 1.00.08 (Nixon, 2002). Results were represented as maps using ArcMap 10.1 (ESRI, 2010).
Results
Areas of endemism identified with the EA by grid-cells
The analysis based on grid-cells identified 8 areas of endemism and 7 consensus areas located along the TVB and towards the central and east of the BB (Table 3, Fig. 2). The endemicity index (E) values vary from 1.8 to 4.63 and 25 species (16.5% from total) showed high e values, characterizing such areas of endemism (Table 3). Figure 2 shows the placement of the consensus areas, which are described as follows:
Western TVB. In the western portion of the TVB coincided 2 consensus areas (2 and 3), including 9 grid-cells supported by 7 TVB strict endemic species (Ageratina jocotepecana B.L. Turner, Ageratum platypodum B.L. Rob., Cosmos ramirezianus Art. Castro, Harker & Aarón Rodr., C. deficiens (Sherff) Melchert, C. longipetiolatus Melchert, Grindelia sublanuginosa Steyerm., and Stevia mascotensis Soejima & Yahara).
Central-Eastern TVB. In this area coincided 3 consensus areas (4, 5 and 7), which included 17 grid-cells located at central and eastern TVB. This area is supported by 13 species, 7 of them TVB strict endemics, 1 BB strict endemic, and 5 shared by both provinces (Table 3).
Central BB. This area comprises 6 consensus areas including 5 grid-cells situated at the central part of BB and is supported by 2 strict endemic species to this province (Chrysanthellum involutum Paul G. Wilson and Eryngiophyllum pinnatisectum Paul G. Wilson).
Eastern BB. This area includes one consensus area comprising 5 grid-cells located in the eastern part of BB and is supported by 3 strict endemic species of this province: Dendroviguiera guerrerana (Panero & E.E. Schill.) E.E. Schill. & Panero, Lasianthaea squarrosa (Greenm.) K.M. Becker, and Stevia mitopoda B.L. Rob.
Table 3 Consensus areas identified by the endemicity analysis using the species of Asteraceae endemic to the BB and the TVB. First the results using the grids-cells are indicated and later those obtained using the hydrological sub-basins.
| Grid-cells of the TVB | ||||
|---|---|---|---|---|
| Consensus area | Number of species defining the area | Maximum score | Endemic species and endemicity value (e) | |
| Western | 2 | 3 | 2.5 |
|
| 3 | 4 | 3.05 |
|
|
| Central-Eastern | 4 | 4 | 3.38 |
|
| 5 | 7 | 4.63 |
|
|
| 7 | 2 | 1.8 |
|
|
| Grid-cells of the BB | ||||
| Central | 6 | 2 | 1.95 |
|
| Eastern | 1 | 3 | 2.42 |
|
| Hydrological subbasins | ||||
| Eastern-Central | 1 | 5 | 3.63 |
|
| 3 | 3 | 2.55 |
|
|
| 4 | 3 | 2.23 |
|
|
| Western-Central | 2 | 3 | 2.32 |
|
| 5 | 4 | 3.30 |
|
|
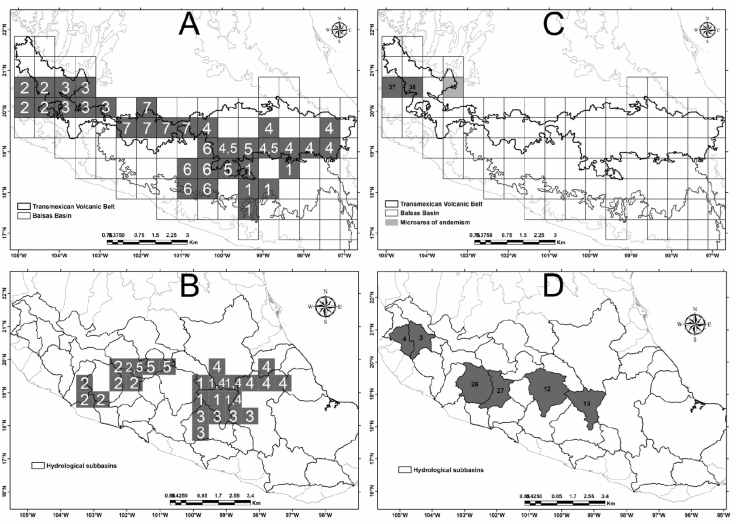
Figure 2 Areas of endemism identified in the biogeographic provinces BB and TVB. A and B show the results obtained with the endemicity analysis (EA) and C and D those obtained with the parsimony analysis of endemicity (PAE). In some cases, there is spatial coincidence between the areas of endemism, for which the overlapping areas are indicated separated by a “comma”.
Areas of endemism identified with the EA by hydrological sub-basins
The analysis based on hydrological sub-basins revealed the occurrence of 5 areas of endemism and 5 consensus areas (Table 3, Fig. 2B). The Endemicity index values vary from 2.23 to 3.63 and 18 species characterizing such areas of endemism showed high e values (Fig. 2B). Figure 2 displays the placement of the consensus areas, which are described as follows:
Eastern-Central area. This area includes 16 grid-cells located in the eastern-central part of the study area and comprises 3 consensus areas (1, 3 and 4). Such grids comprise part of 11 hydrological sub-basins (Río Atoyac-A, Río Balsas-Mezcala, Río Grande de Amacuzac, Río Moctezuma, as well as small parts of the Río Balsas-Zirándalo, Río Cutzamala, Río Jamapa, Río Lerma-Toluca, Río Papagayo, Río Tecolutla and Río Tlapaneco [Fig. 2B]). Eleven species defining this consensus area of which 4 are strict endemics of the TVB, 3 are strict endemics of BBand 4 are shared by both provinces (Table 3).
Western-Central area. This area includes 9 cell grids located at the western-central portion of the study area and comprises 2 consensus areas (2 and 5). These grid cells occupy parts of 10 hydrological sub-basins (Lago de Pátzcuaro-Cuitzeo and Lago de Yuriria, Río Coahuayana, Río Lerma-Chapala, Río Tepalcatepec, Río Tepalcatepec-Infiernillo, as well as small parts of the Río Cutzamala, Río Lerma-Salamanca, Río Lerma-Toluca, Río Nexpe, and Río Tacámbaro). The area is supported by 7 species, 4 of them TVB strict endemics (Astranthium laetificum De Jong, Brickellia leonis Rzed. & Calderón, Eremosis solorzanoana (Rzed. & Calderón) Pruski, and Viguiera moreliana B.L. Turner), 2 BB strict endemics (Pectis leavenworthii Standl. and Roldana glinophylla H. Rob. & Brettell) and 1 distributed both in BB and TVB (Acourtia michoacana (B.L. Rob.) Reveal & R.M. King).
Areas of endemism identified with PAE by grid-cells
A number of 50 equally parsimonious trees based on 151 species and 87 grid-cells were obtained, all them with a TBR score of 194 (Fig. 3A). The strict consensus of these trees recovered a single area of endemism with 4 exclusive species (Fig. 2C). This area records 4 TBV strict endemic species (Cosmos deficiens, C. jaliscensis, C. longipetiolatus, and Stevia mascotensis). In addition, the analysis showed 1 candidate grid-cell to be considered a “micro-area of endemism” because it contains at least 2 TBV strict endemic species (Cosmos ramirezianus and Grindelia sublanuginosa) (Table 4, Fig. 2C). This micro-area of endemism is located at the western part of the province, close to the previous area of endemism (Fig. 2C).

Figure 3 Cladograms obtained with the parsimony analysis of endemicity (PAE) rooted on Operative Geographic Units (OGU´s). A) Strict consensus from 50 equally parsimonious trees using 87 grid-cells. A close-up of the branches of interest is shown on the right. B) The tree obtained using the 30 hydrological sub-basins. A close-up of the branches of interest is shown on the right. In bold the species restricted to the grid-cells are indicated; the white circles represent homoplasy. Above the branches the numerical identifier of the species as cited in Table 1 are shown.
Table 4 Areas of endemism and micro-endemism (*) identified with a parsimony analysis of endemicity (PAE) of the BB and the TVB. The species supporting the areas of endemism are also indicated.
| Areas of endemism by grids | ||
|---|---|---|
| Grid cell | Number of species | Endemic species |
| 37, 38 | 4 | Cosmos deficiens, C. jaliscensis, C. longipetiolatus, Stevia mascotensis |
| 40* | 2 | Cosmos ramirezianus, Grindelia sublanuginosa |
| Areas of endemism by hydrological subbasins | ||
| Endemism area | Number of species | Endemic species |
| Río Ameca-Atenguillo and Río Ameca-Ixtapa | 2 | Cosmos jaliscensis, C. longipetiolatus |
| Río Cutzamala And Río Grande de Amacuzac | 2 | Acourtia lepidopoda, Porophyllum warnockii |
| Río Tepalcatepec and Río Tepalcatepec-Infiernillo | 3 | Pectis leavenworthii, Piqueria glandulosa, Simsia tenuis |
Areas of endemism identified with PAE by hydrological sub-basins
The PAE analysis using the 27 sub-basins resulted in 100 parsimonious trees, recovering 3 areas of endemism supported by 7 endemic species (Table 4, Fig. 3B). One area of endemism is located in the western section of the studied region, corresponding to the Río Ameca-Atenguillo and Río Ameca-Ixtapa sub-basins (Fig. 2D). Both basins share 2 TBV strict endemic species (Cosmos jaliscensis and C. longipetiolatus). A second area of endemism comprises 2 sub-basins placed at the western-central portion, corresponding to the Río Tepalcatepec and Río Tepalcatepec-Infiernillo sub-basins (Fig. 2D); this area is supported by 3 species, 2 of them BB strict endemics (Pectis leavenworthii and Simsia tenuis) and 1 shared by both biogeographic provinces (Piqueria glandulosa) (Table 4). Finally, at the middle of the analyzed sub-basins, is located the third area of endemism, forming part of the Río Cutzamala and Río Grande de Amacuzac sub-basins, supported by 2 endemic species distributed both in BB and TVB provinces (Table 4; Fig. 2D).
Discussion
The BB and TVB biogeographic provinces have been recognized as rich in Asteraceae endemic species restricted to them. Rodríguez-Jiménez et al. (2005) reported 48 endemic species to BB, while Rzedowski (2020) reported 116 endemic species to TVB. A critical review of the endemism in both provinces updated the figures to 39 for the former and 79 for the latter (Table 2). Several discrepancies among the species included in this paper and those former publications will be evident; taxonomic changes, but especially as result of recent fieldwork and species formerly considered strict endemics to one province, are now recorded as part of a wider area. An example of changes in taxonomic circumscription is Wedelia cronquistii B.L. Turner, a species accepted by Rzedowski (2020) but now considered a synonym of W. greenmanii B.L. Turner by Strother (1991), a species reaching northern Mexico in its distribution. One example of endemic species of wider distribution is Coreopsis rhyacophila Greenm., cited by Rodríguez-Jiménez et al. (2005) as endemic to BB, but known to occur at the Sierra Madre Oriental and the xerophytic regions of Hidalgo and Querétaro.
TVB is a mountain chain covered mostly by temperate forests (Pinus, Pinus-Quercus, and Quercus forests) and in some protected ravines the occurrence of humid mountain forests is also recorded (Flores & Gerez, 1994; Rzedowski, 1978). Species of Asteraceae are favored by the presence of such biomes, concentrating the greatest proportion of endemic species. In contrast, only 39 species distribute exclusively in the Balsas Basin, which is predominantly covered by seasonally dry tropical forests (Fernández-Nava et al., 1998; Pineda-García et al., 2007; Rodríguez-Jiménez et al., 2005). A non-negligible number of endemic species (30) is shared between both provinces, without a clearly defined ecological preference (Table 2).
Relations between areas of endemism by AE and PAE
Even using the same data of the Asteraceae species endemic to the biogeographic provinces under study, differences were obtained both in the EA and in the PAE, in the number of areas of endemism identified as well as in the number of species that support them (Morrone, 2014b). However, both methods identified areas of endemism in the western portion of the TVB, when carrying out the EA by cells (Fig. 2A), and when analyzing the data with the PAE by cells and hydrological sub-basins (Fig. 2C, D). It should be noted that Estrada-Márquez et al. (2020) mentioned that the volcanic activity of the TVB has occurred asynchronously along its surface, beginning in its western portion and continuing more recently to the east, highlighting it as an older area in the TVB, a region in which both methods identified the areas of endemism previously indicated.
Particularly, endemicity analysis with both grid-cells and hydrological sub-basins, revealed important sites as areas of endemism. Although they are not characterized by a large number of species (Table 3), they show coincidences in specific areas at the center and west of the study area, specifically in the portion corresponding to the TVB (Fig. 2A, B). PAE identified a single area of endemism based on grid-cells division; however, the analysis carried out allowed to identify one so-called micro-area of endemism, which contains 2 endemic species not shared with contiguous grid-cells. This micro-area is located at the western portion of the study area, near to the only identified area of endemism through the PAE (Fig. 2C).
Areas of endemism in the Transmexican Volcanic Belt province
The placement of most of the areas of endemism towards the eastern portion of the TVB, coincides with other results, for example those obtained by Gámez et al. (2012) studying the mastofauna of the region. Similar results were reported by Villaseñor et al. (2006) with mosses and Senecios (Asteraceae). The high richness localities identified by these authors show important geographic congruence with the areas of endemism identified in this study. The areas of endemism here found are included within the East biogeographic district proposed by Escalante et al. (2007) based on mammal data.
At the western and central portions of the TVB, the areas of endemism identified form part of those found by Estrada-Márquez et al. (2020) with a set of Magnoliophyta species. These authors identified such areas of endemism towards the center of the country, particularly in the BB and TVB provinces, and highlighted the relevance of Asteraceae species in identifying them. Likewise, Rodríguez et al. (2018) pointed out the existence of areas of endemism and centers of high species richness for several groups of angiosperms in the western portion of TVB, coinciding several of them with the areas of endemism found in this study. In the same way, the regions of high vascular plant species richness found by Rzedowski (2020) in Jalisco and Michoacán, coincide with the areas of endemism here identified. In summary, the Transmexican Volcanic Belt has been recognized as an area rich in endemic mammals and other groups (Ceballos & Navarro, 1991), as well as a center of diversification, endemism and biogeographic transition (Gámez et al., 2012). Similarly, Villaseñor and Ortiz (2007), studying the family Asteraceae, coincide that the TVB is a center of diversity and endemism for the family.
Areas of endemism in the Balsas Basin province
Our results support previous findings, pointing out the central-eastern portion of the BB as an important area of endemism. In a similar way, the studies of Castro-Torreblanca et al. (2014) with birds reported the eastern region as of the 3 highest species concentration centers. Rodríguez-Jiménez et al. (2005) pointed out that states that concentrate more than 50% of BB´s endemic vascular plant species are Guerrero, Estado de México and Michoacán, which coincide in part with the area of endemism identified in this region. Montaño-Arias et al. (2018) discussed the high endemism values found in the tropical dry forests of the western portion of the BB. The BB areas of endemism found in this study agree with such statement, placing the western part of the BB as the richest in endemic species. Finally, the areas of endemism found towards the central portion of BB coincide with the results of Sosa and De-Nova (2012), who identified one area of endemism towards the center of the BB province based on a set of Mexican angiosperm species.
Natural Protected Areas and areas of endemism
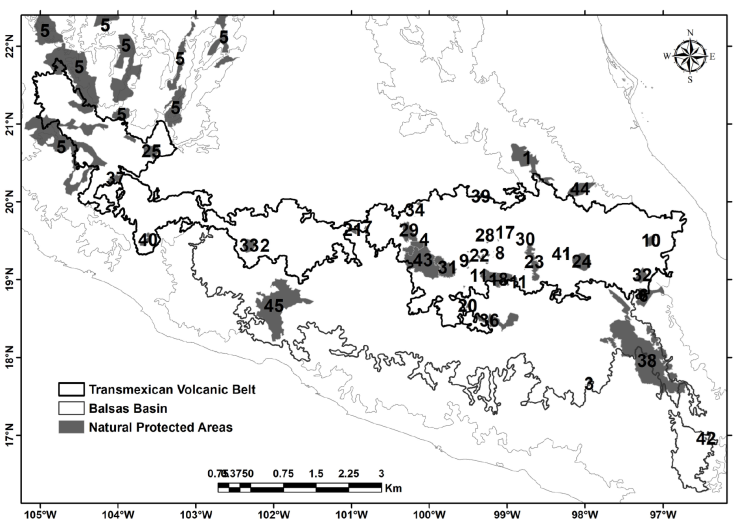
Within the study area, 45 Natural Protected Areas (NPAs) include all or part of their territory, 35 in the TVB, 5 to the BB and 5 sharing surfaces between both provinces (Table 5, Fig. 4). Most of the NPAs are located towards the center and east of the TVB, while in BB a single area is located towards its west part and the other concentrated towards its east portion (Fig. 4). The areas of endemism identified with both EA and PAE indicate that 38 NPAs include portions of the endemism areas (Table 5).
Table 5 Natural Protected Areas (NPAs) including part of the areas of endemism identified with the endemicity analysis (EA) and the parsimony analysis of endemicity (PAE). The asterisks indicate if it is an NPA of the TVB (*), the BB (**) or share part of both (***).
| Natural protected areas | Areas of endemism by EA | Areas of endemism by PAE | Total areas of endemism | |||
|---|---|---|---|---|---|---|
| Grid-cells | Hydrological sub-basins | Grid-Cells | Hydrological sub-basins | |||
| 1 | *Barranca de Metztitlán | 0 | ||||
| 2 | *Barranca del Cupatitzio | 7 | 2 | 26, 27 | 3 | |
| 3 | **Boquerón de Tonalá | 0 | ||||
| 4 | *Bosencheve | 4 | 12,13 | 2 | ||
| 5 | *C.A.D.N.R. 043 Estado de Nayarit | 2, 3 | 37, 38 | 4,3 | 3 | |
| 6 | *Cañón del Río Blanco | 4 | 1 | |||
| 7 | *Cerro de Garnica | 7 | 5 | 12,13 | 3 | |
| 8 | *Cerro de La Estrella | 4, 5 | 1, 4 | 2 | ||
| 9 | *Ciénegas del Lerma | 5 | 1, 4 | 2 | ||
| 10 | *Cofre de Perote o Nauhcampatépetl | 4 | 4 | 2 | ||
| 11 | ***Corredor Biológico Chichinautzin | 4, 5 | 1, 4 | 12,13 | 3 | |
| 12 | *Cumbres del Ajusco | 1,4 | 12,13 | 2 | ||
| 13 | *Desierto de los Leones | 5 | 1, 4 | 2 | ||
| 14 | *Desierto del Carmen o de Nixcongo | 5 | 1 | 12,13 | 3 | |
| 15 | *El Histórico Coyoacán | 1, 4 | 1 | |||
| 16 | *El Jabalí | 0 | ||||
| 17 | *El Tepeyac | 1, 4 | 1 | |||
| 18 | ***El Tepozteco | 4, 5 | 1, 4 | 12, 13 | 3 | |
| 19 | *Fuentes Brotantes de Tlalpan | 5 | 1, 4 | 2 | ||
| 20 | **Grutas de Cacahuamilpa | 1 | 1 | 12, 13 | 3 | |
| 21 | ***Insurgente José María Morelos | 7 | 5 | 12, 13 | 3 | |
| 22 | *Insurgente Miguel Hidalgo y Costilla | 5 | 1, 4 | 2 | ||
| 23 | *Iztaccíhuatl-Popocatépetl | 4, 5 | 1, 4 | 12, 13 | 3 | |
| 24 | *La Montaña Malinche o Matlalcuéyatl | 4 | 4 | 2 | ||
| 25 | *La Primavera | 3 | 40 | 2 | ||
| 26 | *Lagunas de Zempoala | 5 | 1, 4 | 12, 13 | 3 | |
| 27 | *Lomas de Padierna | 5 | 1, 4 | 2 | ||
| 28 | *Los Remedios | 1, 4 | 1 | |||
| 29 | *Mariposa Monarca | 4 | 12, 13 | 2 | ||
| 30 | *Molino de Flores Netzahualcóyotl | 4 | 1, 4 | 2 | ||
| 31 | *Nevado de Toluca | 4, 5 | 1 | 12, 13 | 3 | |
| 32 | *Pico de Orizaba | 4 | 4 | 2 | ||
| 33 | *Pico de Tancítaro | 7 | 2 | 26, 27 | 3 | |
| 34 | *Rayón | 4 | 1 | |||
| 35 | *Sacromonte | 4, 5 | 1, 4 | 2 | ||
| 36 | ***Sierra de Huautla | 1 | 1, 3 | 12, 13 | 3 | |
| 37 | *Sierra de Quila | 2, 3 | 4, 3 | 2 | ||
| 38 | **Tehuacán-Cuicatlán | 4 | 1 | |||
| 39 | *Tula | 0 | ||||
| 40 | *Volcán Nevado de Colima | 0 | ||||
| 41 | *Xicoténcatl | 4 | 4 | 2 | ||
| 42 | **Yagul | 0 | ||||
| 43 | ***Z.P.F.T.C.C. de los ríos Valle de Bravo, Malacatepec, Tilostoc y Temascaltepec | 4, 6 | 1 | 12, 13 | 3 | |
| 44 | *Z.P.F.V. la Cuenca Hidrográfica del Río Necaxa | 0 | ||||
| 45 | **Zicuirán-Infiernillo | 26, 27 | 1 | |||
| Total NPAs | 33 | 29 | 2 | 19 | 38 | |
Based on the EA by grid-cells, 33 NPAs matched with them, while for the analysis using hydrological subbasins, 29 NPAs matched with their surface (Table 5). In contrast, the area of endemism identified with PAE using grid cells, as well as the micro-area of endemism matched with 2 NPAs; the same analysis based on hydrological subbasins matched with 19 NPAs (Table 5). A figure of 14 NPAs coincided in 3 out of 4 analyses carried out to identify the areas of endemism (Table 5); among them stand out the Corredor Biológico Chichinautzin, El Tepozteco, Insurgente José María Morelos, Sierra de Huautla and Z.P.F.T.C.C. de los ríos Valle de Bravo, Malacatepec, Tilostoc y Temascaltepec (Table 5).
Implications for the conservation of Asteraceae
Endemism has been of particular relevance for the design and delimitation of protected areas. Due to their geographic restriction, endemic species are sensitive to the modification of their environment, particularly if it is due to anthropogenic agents (Alcántara & Paniagua, 2007). The Asteraceae family does not only stand out for its endemism, but because it is well known taxonomically, it is easy to sample, it has representatives of all types of growth forms, and it is distributed over a wide range of habitats. It has also turned out to be a good surrogate for country’s biodiversity (Villaseñor et al., 1998, 2007). Thus, conservation of its species as well as the ecosystems in which they are found would also contribute to conserve much of the Mexican plant diversity, especially that threatened by several anthropogenic and natural factors.
Despite the fact that 84% of the NPAs already established inside the study region matched with several of the areas of endemism identified in this study, there are still important regions to be considered due to their endemism and actually not protected by any NPA. Such is the case of the central and southeast portion of the BB, as well as some regions of its western portion; the same situation can be observed within the TVB in its western portion, where there are not enough NPAs (Fig. 4). For this reason, we propose that those areas of endemism whose surface does not have any official protection, have some strategies for their conservation especially due to their coincidence with regions identified as important for different biotic groups (plants, birds and mammals), highlighting them as potentially irreplaceable areas, one of the utmost criteria for the definition of priority areas for conservation (Méndez-Larios et al., 2005).

