











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Durante el siglo XIX se consolidó la idea que la biodiversidad que habita la Tierra muestra una distribución espacial no azarosa. Desde entonces, se distinguieron ciertos patrones espaciales (Juárez-Barrera et al., 2018): a) el gradiente latitudinal de riqueza y remplazo de especies, b) la variación latitudinal del fenotipo de grupos de cuadrúpedos (reglas zoogeográficas y de formas de vida vegetal), c) la sucesión de ensambles de formas de vida vegetal a lo largo de la latitud y de la elevación, d) la extensión y diferenciación de los linajes y e) la correspondencia entre las áreas de distribución de diferentes taxones, fueran éstas continuas (regiones biogeográficas) o disyuntas. De este último patrón derivó la hipótesis de múltiples centros de origen donde surgieron las especies nuevas, y con ello el debate sobre el origen único o múltiple de las especies, tanto en el espacio como en el tiempo. Desde una perspectiva metafísica, algunos autores supusieron que una misma especie podía aparecer en diferentes áreas y tiempos por una voluntad supranatural, como sostenía Louis Agassiz (1848). Desde otra perspectiva más secular, la premisa fue el origen único de las especies en un determinado espacio y tiempo, como sostenía Alphonse de Candolle (1855), quien interpretó a las regiones botánicas como la evidencia de la existencia de floras ancestrales que a través del tiempo habían dado lugar a las floras actuales.
El surgimiento de la hipótesis holarticista
Luego de la difusión y adopción de la teoría de la evolución por selección natural (Darwin, 1859), el estudio de los patrones biogeográficos, hasta entonces ampliamente aceptados, se fue relegando en la medida que se iba adoptando un nuevo enfoque para el estudio de la distribución espacial de los taxones. Las áreas de distribución de los distintos grupos se concibieron como entidades naturales sometidas a un cambio continuo. Bajo esta nueva perspectiva de una biogeografía dinámica, parecía carecer de sentido la búsqueda de sistemas de regionalización biogeográfica, los que representaban el viejo enfoque meramente descriptivo. Otro postulado relacionado con esta concepción dinámica, derivado de la teoría darwinista, fue que los grupos dominantes surgían originalmente en el hemisferio norte, para posteriormente invadir las áreas australes (Juárez-Barrera et al., 2018). En esas áreas de gran extensión, la variabilidad de los individuos de una misma especie y las presiones de selección se maximizaban. Por ello, el asumir que habían ocurrido oleadas sucesivas y continuas de invasión, hacía irrelevante el interés por encontrar una regionalización natural, como lo había propuesto anteriormente Sclater (1858). En las últimas décadas del siglo XIX, varios estudiosos de la distribución geográfica de la vida habían abandonado el interés por establecer clasificaciones biogeográficas. Allen (1878) estableció que todos los patrones latitudinales y graduales de la distribución de la vida en la Tierra podían ser razonablemente explicados por lo que él llamó “ley general de la adaptación”. Bajo esta perspectiva, las regiones postuladas por Sclater (1858) y Wallace (1876) eran un mero constructo que solo se justificaba por razones didácticas. El incluir en una misma región a organismos con formas de vida completamente diferentes, era no solo artificial, sino irrelevante.
“Afirmo sin vergüenza ... la distribución geográfica de la vida está por necesidad en armonía con una "ley de adaptación", es decir, de adaptación climática; que tal ley es legítima con esta relación, y que la referencia al "encubrimiento superficial" que adapta esencialmente a organismos de diferentes formas de vida a formas de vida similares es totalmente irrelevante para el punto en cuestión, una comparación de cosas que son en cierto sentido incomparable, además, que la "ley de distribución de vidas en zonas circumpolares" se aplica tanto en un sentido general como a los detalles: "a grupos de géneros y familias", así como a las especies” (Allen, 1878: 326).
Bajo el determinismo ecológico de Allen, las causas históricas de los patrones biogeográficos quedaron relegadas, en favor de la determinación de los factores ambientales presentes sobre la distribución de las especies.
El naturalista canadiense William Diller Matthew (1871-1930) siguió las ideas de Wallace, en particular la idea de sucesivas olas de migración provenientes de centros de evolución septentrional. Sin embargo, a diferencia de Wallace, Matthew (1915) sostuvo que el centro de origen de las especies no solo abarcaba el norte de Eurasia, sino que también incluía el norte de América. A la unión de estas 2 regiones norteñas la llamó región Holártica, retomando el nombre acuñado por Alfred Newton, en una correspondencia a Wallace, donde propuso la unión de las regiones Neártica y Paleártica (Ebach, 2015: 162).
De acuerdo con Matthew (1915), la flora y la fauna que habitaban el hemisferio meridional podían explicarse como remanentes de especies surgidas originalmente en el norte, temporalmente dominantes, que posteriormente se extinguieron en su centro de origen y fueron desplazadas hacia el sur por el surgimiento de nuevas especies competitivamente superiores. Así, algunos grupos, como los marsupiales, se habrían originado en el norte, luego migraron hacia el sur y posteriormente fueron extinguidos en su área de origen por los placentarios, con excepción de aquellos que quedaron confinados en áreas aisladas geográficamente durante largos periodos de tiempo, como Australia y la Patagonia.
“La no existencia de marsupiales en el registro de faunas en el Terciario Tardío de Europa occidental es un hecho significativo en la distribución de este orden. Su no existencia en el Terciario Temprano de Asia tiene poco peso, y hasta cualquier descubrimiento reciente éste era nulo. Esto se debe a que las faunas terciarias tardías de Europa occidental son extensas y variadas, representan varias facies y, en conjunto, probablemente la mayoría, aunque no todos, de los mamíferos que viven en esa región. Uno podría esperar que si algún marsupial hubiera sobrevivido allí hasta el Terciario Tardío, algunos restos de ellos habrían aparecido entre las faunas fósiles conocidas” (Matthew, 1915: 165).
México como área o zona de transición
Más tarde, los biogéografos líderes de la “Síntesis Moderna” (Darlington, 1957; Mayr, 1976; Simpson, 1965) ya no aceptaron la idea de un solo centro de origen norteño. Ante la innegable evidencia de la distribución congruente entre varios grupos de taxones, propusieron múltiples centros de origen. En cada uno de ellos, aislados durante largos periodos, evolucionaron faunas típicas. Cuando eventualmente estas faunas establecían contacto geográfico, se producía un intercambio faunístico. Sin embargo, se mantuvo la idea de la predominancia de la fauna norteña sobre la sureña. A partir de ahí se retomó la idea de Wallace, desarrollada por Matthew, según la cual la biota mexicana es el resultado de la mezcla entre las faunas neártica y neotropical que constituye un área de transición biótica ubicada en el sureste de México (Darlington, 1957). Al igual que Matthew, Ernst Mayr sostuvo la idea de la dominancia de los grupos norteños durante los periodos de intercambio:
“... Los elementos norteños han tenido mucho más éxito que los elementos de América del Sur al cruzar la brecha de agua terciaria entre América del Norte y América del Sur. Muchos de los supuestos elementos de origen norteño, las tangaras, por ejemplo, han tenido una radiación adaptativa extremadamente exitosa en América del Sur, por lo que ahora pueden considerarse elementos verdaderamente sudamericanos” (Mayr, 1976: 558-559).
A pesar de que Mayr admitía que podía haber excepciones a la regla y la invasión también podía ocurrir de sur a norte, había un sesgo marcado de migraciones norte - sur:
“Si bien solo dos familias de origen sureño, Trochilidae y Tyrannidae, han invadido con éxito América del Norte, prácticamente todas las familias de presunto origen norteamericano están ahora bien representadas en América del Sur” (Mayr, 1976: 558-559).
Mayr, además, terminó por desmantelar el propósito de hacer regionalización en biogeografía, que ya desde fines del siglo XIX se asociaba con una biogeografía obsoleta, esencialmente descriptiva (Ebach, 2015: 135): “Eventualmente se comprendió que el enfoque - Fragestellung [pregunta] - de esta zoogeografía esencialmente estática estaba equivocado. En lugar de pensar en regiones fijas, era necesario pensar en faunas fluidas” (Mayr 1946: 5).
Simpson (1965) sostuvo una posición menos rígida y aceptó un intercambio faunístico bidireccional más intenso entre América del Norte y del Sur, a través del puente de Panamá, aunque sostuvo también la idea de la predominancia de la mastofauna norteña sobre la del sur. Simpson mostró cómo las faunas de América Norte y del Sur se mantuvieron aisladas hasta antes del Pleistoceno, que fue cuando se dio un intercambio masivo de la mastofauna que elevó temporalmente el número de familias presentes en cada área, pero que finalmente se revirtió debido a la extinción diferencial de muchas familias (tabla 1, fig. 1).
Tabla 1 Mamíferos terrestres de América del Norte (AN) y América del Sur (AS), durante el Cenozoico tardío.
| Periodo | Época | AS Total | AS únicas | AS y AN | AN únicas | AN Total | Similitud (100C/N1) |
|---|---|---|---|---|---|---|---|
| Cuaternario | Reciente | 30 | 15 | 15 | 8 | 23 | 65 |
| Pleistoceno | 35 | 14 | 21 | 11 | 34 | 62 | |
| Plioceno | Tardío | 26 | 21 | 5 | 22 | 27 | 19 |
| Medio | 25 | 23 | 2 | 24 | 27 | 8 | |
| Temprano | 26 | 24 | 2 | 26 | 28 | 8 | |
| Mioceno | Tardío | 24 | 23 | 1 | 25 | 26 | 4 |
| Medio | 23 | 23 | 0 | 26 | 27 | 0 |
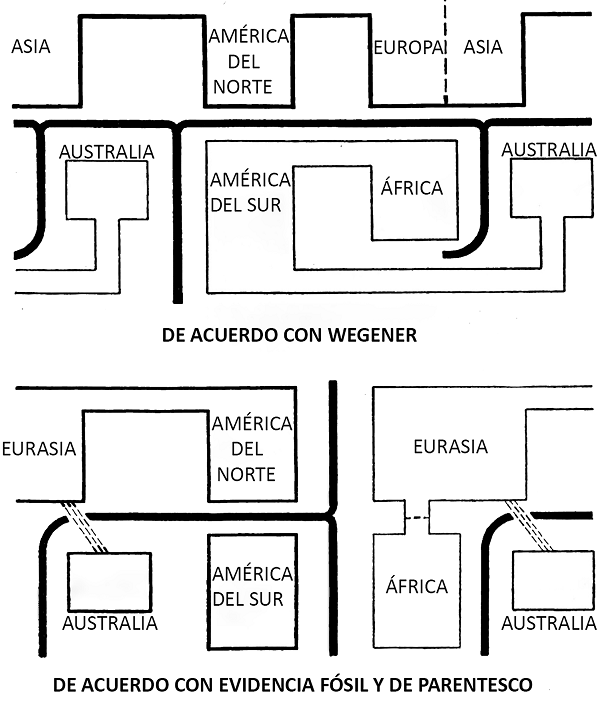
Figura 1 Relaciones entre las regiones zoogeográficas. a) Relación de los continentes de acuerdo con la teoría de Wegener (1920); b) conexiones y barreras entre los bloques continentales en el Eoceno diferentes a las que propuso Wegener.
Darlington (1957) propuso la existencia de un área de transición Centroamericano-Mexicana, cuyo límite indicativo es el Istmo de Tehuantepec (fig. 2). Según Darlington (1957), este límite es traspasado por la fauna del sur hacia el norte y por la de América del Norte en sentido inverso, que transitan a través del puente de Panamá.

Figura 2 Ubicación de la línea divisoria (círculos negros) entre las regiones Neártica y Neotropical, de acuerdo con Darlington (1957).
“Las faunas de vertebrados de las regiones Neártica y Neotropical son muy diferentes en muchos aspectos, pero también comparten muchas familias, géneros y algunas especies, y hay una transición especialmente en México y América Central, y algunas se extienden a áreas más amplias” (Darlington, 1957: 456).
“Esta zona de transición nunca ha sido tratada adecuadamente en detalle por los zoogeógrafos. No ha recibido la importancia prestada a la Línea de Wallace y la Wallacea, aunque es probablemente de igual importancia” (Darlington, 1957: 460).
Según Darlington (1957), cada taxón de la fauna neártica avanza hacia el sur más allá del Istmo de Tehuantepec de acuerdo con su propia capacidad de dispersión hasta Nicaragua e incluso hasta Chiriquí (Panamá). En sentido contrario, ocurre algo similar, donde los taxones del sur pueden alcanzar hasta el río Bravo o el suroeste de los Estados Unidos de América. Pero hay 2 rasgos que Darlington destaca particularmente: uno es el predominio de la fauna neártica sobre la neotropical, y otro, el carácter idiosincrásico de las historias de dispersión desde centros de origen particulares. Bajo la perspectiva de Darlington, resulta irrelevante el reconocimiento de patrones congruentes de distribución entre diferentes linajes.
Desde una perspectiva diferente, Léon Croizat distinguió 5 grandes portales (“gates”) biogeográficos, que hoy corresponden con la definición de zonas de transición biogeográfica por adición (Ferro y Morrone, 2014): áreas amplias donde 2 o más biotas se superponen diferencialmente. A diferencia de las historias idiosincrásicas de dispersión de Darlington (1957: 460-461) y Mayr (1976: 553), Croizat (1958) puso énfasis en buscar la congruencia entre las distribuciones de 2 o más especies para descubrir conjuntos de especies con historias evolutivas compartidas que ocurrieron en un espacio común y sufrieron conjuntamente los efectos de los mismos eventos geológicos históricos. El método de Croizat permite reconocer eventos tectónicos tanto de fragmentación como de convergencia. Estos últimos están representados por lo que Croizat denomina “nodos”, que son áreas donde 2 o más trazos generalizados convergen y representan el encuentro de biotas con distintas historias. Más aún, Croizat encontró algunas áreas particularmente complejas, donde se concentran muchos nodos y trazos, a las cuales denominó portales. Entre ellos destaca el portal Mesoamericano-Antillano, que incluye el sur y sureste de México, como una gran zona de convergencia (fig. 3). Croizat aceptó abiertamente una geografía cambiante que tiene por causa la translación de los continentes, aunque discrepó sustancialmente con la teoría de la deriva continental de Wegener (1920) en cuanto a la secuencia de las fragmentaciones. La panbiogeografía de Croizat representó un modelo de explicación que subvirtió radicalmente los principios de la biogeografía neodarwinista, pues abrió la posibilidad de explicar la complejidad de la biota mexicana desde perspectivas nuevas y claramente diferentes a las explicaciones dispersalistas-holarticistas.
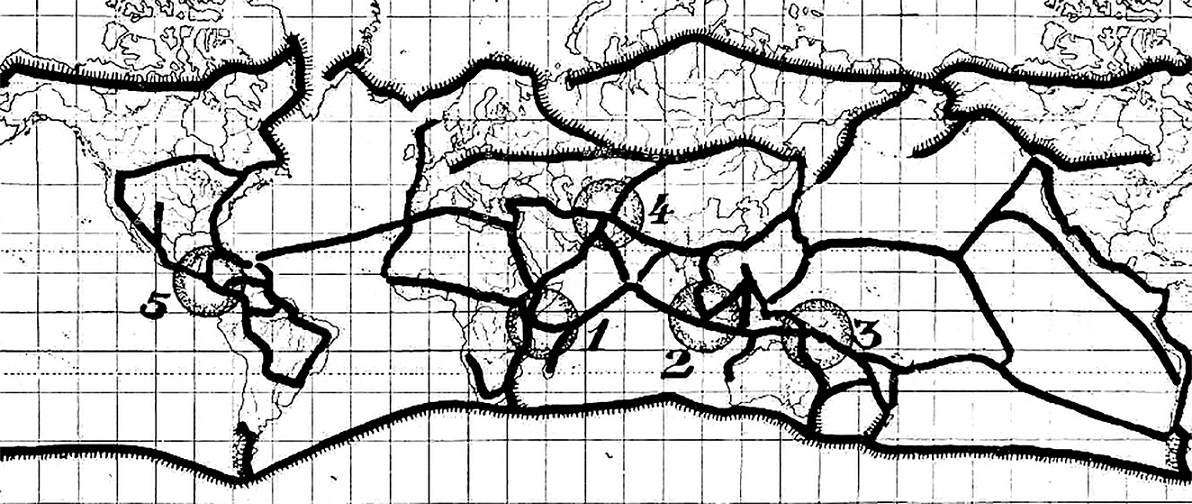
Figura 3 Resumen del análisis biogeográfico de Croizat (1958), donde se destaca el nodo 5 (México-Mesoamérica) como un área particularmente compleja.
Algunos científicos mexicanos (v. gr., Hoffman, 1933; Miranda, 1959) reconocieron claramente las relaciones complejas de la biota de México. Estos autores discutieron explícitamente las ideas del fijismo geográfico, el extensionismo (sensu Fichman, 1977) y el translacionismo, aunque no se comprometieron en particular con ninguna de ellas (Miranda, 1959).
El concepto de zona de transición desde la perspectiva de los estudiosos del territorio mexicano
A finales del siglo XIX e inicios del XX, varios botánicos y zoólogos extranjeros se establecieron en México de forma permanente e hicieron valiosas contribuciones al conocimiento de la biota mexicana. Podemos mencionar entre los pioneros a Karl Friedrich Reiche, Eizi Matuda, Cassiano Conzatti y Carlos Hoffmann. Ya desde los trabajos de Carlos Hoffmann se mantuvo la idea de transición como resultado de la mezcla de 2 faunas en México: “En términos generales puede decirse, que las formas del sur proviniendo de centros australes, se reparten hacia el norte tomando su camino preferentemente por las tierras calientes y templadas de las dos costas y que viceversa, las formas septentrionales se extienden al sur siguiendo en su mayoría el curso de las grandes cordilleras. De esta manera se efectúa un constante contacto entre el norte y el sur y un progreso lento de géneros y especies sobre la base de una aclimatación lenta y graduada durante el curso de una infinidad de años” (Hoffmann, 1933: 208).
Entre las crisis económicas, las 2 guerras mundiales y otras guerras y revoluciones internas ocurridas en varios países, México recibió también a personajes que consolidarían las nuevas instituciones científicas, como Faustino Miranda, Enrique Rioja Lo Bianco, Cándido Bolívar y Federico Bonet, entre otros. Sus ideas ejercerían una influencia importante sobre los estudios de generaciones posteriores de estudiantes e investigadores.
En México, Faustino Miranda tuvo una vida académica relativamente corta debido a su muerte temprana. No obstante, en sus trabajos sobre la vegetación de México encontró patrones de distribución geográfica que lo llevaron a proponer relaciones históricas complejas que podrían calificarse como vicariancistas. Miranda (1959), además de utilizar términos de connotación ecológica, también recurrió a causas históricas al elaborar sus explicaciones biogeográficas.
“Se pueden distinguir en la zona tropical húmeda americana dos grupos florísticos con arreglo a las relaciones de distribución de sus elementos bicontinentales: uno que podemos llamar ‘anfi-Atlántico’, con ligero predominio de elementos africanos-americanos, se encuentra en los lugares bajos, sobre todo en los pantanosos y tiene actualmente su mayor desarrollo hacia la zona ecuatorial; el otro, o ‘anfi-Pacífico’, con acentuado predominio de elementos asiático-americanos, caracteriza a las regiones montañosas sobre todo hacia el límite norte de la zona tropical” (Miranda, 1959: 129).
“[...] Pudiera pensar que lo que antecede habla a favor de la difusión de los géneros africano-americanos a través del Atlántico Central (Brasil-África), ya por emigración a la larga distancia, o bien mediante puente intercontinental, o por contigüidad de los continentes (translaciones continentales)” (Miranda, 1959: 130).
Miranda (1959) planteó así 3 explicaciones alternativas para entender los patrones de la vegetación de México: 1) una migración a larga distancia, como sería el intercambio entre el sur de América y África, 2) un puente continental hipotético a través del Atlántico, o 3) una conexión antigua entre ambos continentes, que permitiría la comunicación entre ambas áreas. Continúa diciendo: “Diversos matices de la vegetación no dependen sólo del clima y el suelo, sino también de la evolución propia de la vegetación misma o de los elementos que la forman, evolución que por lo general se halla relacionada con la evolución de los procesos geológicos” (Miranda y Hernández, 1963: 30).
Como se puede apreciar, Miranda consideró que los factores ecológicos son insuficientes para explicar los patrones biogeográficos; en sus razonamientos está implícita la idea de que vida y tierra comparten una misma historia. Recurre así, tanto a causas ecológicas como paleoclimáticas, en su intento por explicar los patrones de distribución de la flora mexicana.
Los elementos florísticos de Rzedowski
Jerzy Rzedowski (1965) también hizo énfasis en la complejidad histórica de la flora mexicana. Reconoció diferentes géneros que relacionan a ésta con otras floras tropicales como la antillana, centroamericana, sudamericana del Pacífico y del Caribe, la del Amazonas e incluso la africana. Las floras de los bosques templados de México comparten géneros con el resto de América del Norte y Eurasia. A diferencia de naturalistas del siglo XIX, como Fournier (1877), Rzedowski (1965) no invocó puentes como la mítica Atlántida, sino planteó posibles rutas y tiempos de migración que convergieron en México.
Rzedowski (1965, 1978, 1991) reconoció la existencia de 6 elementos florísticos principales de distinto origen en el territorio nacional: 1) elementos con afinidades meridionales, es decir, taxones neotropicales relacionados con formas centroamericanas y sudamericanas, los cuales constituían la mayor parte de la flora del país; 2) elementos con afinidades septentrionales, los cuales son taxones neárticos, relacionados con formas de regiones templadas y frías de EUA y Canadá; 3) elementos con afinidades antillanas, los cuales son taxones neotropicales con formas emparentadas con las de las Antillas; 4) elementos con afinidad a los del este de Asia, los cuales son más evidentes en formas de montaña, como el bosque mesófilo de montaña; 5) elementos con afinidades africanas, poco frecuentes y presentes en plantas de clima cálido; 6) finalmente, elementos endémicos, no tan abundantes como en Australia y África del sur, pero que le confieren un sello particular a la flora del país. Si bien algunos de estos elementos coinciden en parte con los que reconoce Halffter, Rzedowski no profundizó en las causas que han producido estas afinidades y se limitó a invocar la dispersión como causa general.
Las ideas sobre la zona de transición de Halffter
A diferencia de Rzedowski, Gonzalo Halffter intentó recurrentemente buscar explicaciones más concretas acerca de los patrones de distribución de las especies. Sus trabajos (Halffter, 1962, 1964, 1965, 1967, 1976) tuvieron un impacto mayor sobre los análisis biogeográficos posteriores e incluso hasta nuestros días, sobre el complejo mosaico geográfico de la biota mexicana (Morrone, 2015). Su acierto fue someter a prueba los patrones biogeográficos que propuso, derivados de sus amplios trabajos con escarabajos estercoleros y necrófagos (Morrone, 2015; Halffter y Morrone, 2017). A lo largo de sus trabajos, Halffter ha ido refinando su “teoría sobre la Zona de Transición Méxicana” (ZTM), como él la denomina, al comparar información taxonómica con información geológica (Halffter y Morrone, 2017: 82).
Como muchos autores, en sus primeros trabajos Halffter coincide en lo general con la idea de que no existe un límite definido entre las regiones Neártica y Neotropical, sino una superposición compleja, determinada por las particularidades fisiográficas de México, su historia geológica y las condiciones ecológicas. En todo caso, la línea divisoria entre ambas regiones estaría definida geográficamente por la Sierra Madre Occidental, la Faja Volcánica Transmexicana y la Sierra Madre Oriental, que en conjunto forman una gran barrera orográfica que separa el Altiplano Mexicano de las tierras bajas tropicales. La configuración fisiográfica particular de la ZTM reside en que ninguna otra zona de transición tiene tal profundidad y complejidad, ya que cuenta, por un lado, con 2 cordilleras que corren latitudinalmente y permiten la entrada de elementos templados norteños, y por otro, con 2 planicies costeras que permiten la entrada de elementos tropicales provenientes del sur. Ello explica la extensa superposición de faunas con historias distintas. Además, otro factor que ha contribuido a incrementar la complejidad de la ZTM es que ha habido diferentes eventos de dispersión (Halffter y Morrone, 2017). Halffter adicionalmente señaló que el límite de esta zona compleja no es el mismo para diferentes taxones (Halffter, 1965). Por ejemplo, en el caso de los vertebrados, este límite es más claro que en el de los insectos.
“En relación con los límites entre ambas regiones e incluso directamente con la existencia de las mismas, está el hecho, poco evidente, casi nunca mencionado, pero no por eso menos real, de que las dos regiones se han establecido utilizando casi exclusivamente la distribución de los vertebrados terrestres, y no la totalidad de la fauna” (Halffter, 1965).
“En el Altiplano dominan insectos de origen sudamericano [...] un contraste marcadísimo con la distribución de los vertebrados que en esta área son claramente neárticos” (Halffter, 1967: 109).
Previamente, el concepto de área o Zona de Transición Mexicana ya había sido abordado implícita o explícitamente por una serie de autores desde el siglo XIX, como Alexander von Humboldt, Alphonse de Candolle, Alfred R. Wallace, Hobart M. Smith, Philip Darlington y Léon Croizat, entre otros. El consenso general es que la ZTM tiene límites complejos, producto de la superposición de varias áreas de distribución geográfica. Algunos de estos autores propusieron que esta zona de transición se explicaba por la superposición continua y gradual de las áreas de distribución de varios taxones de origen distinto. Sin embargo, de acuerdo con Halffter (1962, 1965, 1966, 1967, 1987; Halffter y Matthews, 1966; Halffter y Morrone, 2017), dentro de esa aparente continuidad se pueden reconocer conjuntos de especies que muestran patrones de extensión-diferenciación congruentes. Al revisar los trabajos de Halffter, llama la atención que su idea acerca de tales patrones fue cambiando desde su primera propuesta en 1962 hasta sus trabajos más recientes (Morrone, 2015). En su primera etapa, propone a la dispersión como la causa principal de los patrones, pero posteriormente asigna un papel más importante a las causas históricas (Halffter, 1987). Inicialmente, Halffter se refirió a los patrones que encontró en la zona de transición como “patrones de dispersión” aunque después los llamó “patrones de distribución”:
“Aunque en algunos de sus primeros trabajos, Halffter usa el término “patrón de dispersión”, esto no es correcto ya que el patrón no está determinado exclusivamente por los fenómenos de dispersión, sino también por la evolución que ha ocurrido en la zona y, por lo tanto, por la vicarianza” (Halffter et al., 1995: 152).
Halffter (1965) reconoció patrones congruentes de extensión-diversificación que lo llevaron a retomar la idea de Smith (1941) de “horofauna”, entendida como un conjunto de linajes que muestran distribución y diferenciación congruente debido a que se expandieron simultáneamente por causa de los mismos eventos geológicos o paleo-climáticos. Este mismo concepto es nombrado años más tarde como “cenocrón” por Osvaldo Reig (1968), al analizar la transición entre las biotas patagónica y neotropical. Quizás el antecedente más antiguo sobre el reconocimiento y estudio de este tipo de patrones lo encontramos en las obras de Augustin y Alphonse de Candolle. En 1820, Augustin de Candolle propuso el concepto de “género endémico” en un sentido un poco diferente al utilizado en la actualidad (Juárez-Barrera et al., 2018). Lo distinguió del concepto de “género esporádico”. Con estos 2 conceptos distinguió patrones de extensión y diversificación. Un “género endémico” es un taxón altamente diversificado, cuyas especies no han traspasado las barreras que los confinan. Así, los géneros endémicos les dan identidad a las regiones. Por otra parte, los “géneros esporádicos” muestran una distribución amplia, más diferenciados en especies en una región que en las otras. Para Alphonse de Candolle (1855), este último patrón podría servir para relacionar históricamente a 2 o más regiones (Juárez-Barrera et al., 2018). Hoy en día, desde un enfoque biogeográfico histórico, un taxón endémico es simplemente un taxón propio o exclusivo de un área.
A lo largo de sus trabajos, el criterio de Halffter para asignar el origen y la edad de cada patrón de distribución se fundamenta en 4 aspectos: 1) la amplitud del área de distribución de cada linaje, 2) las afinidades filogenéticas de los linajes, 3) la diversidad diferencial de especies de un mismo linaje entre áreas o regiones, y 4) la amplitud ecológica como indicador de la edad del taxón.
“Estos grupos, originados en el Viejo Mundo, tienen una penetración muy antigua por la ruta del norte hacia el continente americano. Su riqueza y diversidad (tanto taxonómica como ecológica) son mucho mayores en el Viejo Mundo que en América. A partir de tal riqueza y diversidad (tanto a nivel específico como supra-específico) se puede deducir que el principal centro de evolución ha sido el Viejo Mundo. Aunque haya habido una expansión secundaria, ésta habría afectado a un número muy pequeño (a veces solo una) de especies ancestrales que derivaron claramente de las poblaciones del Viejo Mundo" (Halffter, 1976: 7).
“Los grupos que han dado lugar a elementos paleoamericanos, tanto en América como en el Viejo Mundo, revelan una amplia diversificación ecológica, aunque es obvio que predominan en climas cálidos y templados y en ningún caso son elementos típicos de climas fríos (excepto por consideraciones muy locales, las especies que se derivan claramente de troncos tropicales o subtropicales que logran colonizar las partes más altas y más frías de las montañas tropicales no pueden tomarse como elementos de climas fríos). Existe un marcado contraste dentro y fuera de América entre los elementos paleoamericanos y los que siguen en el patrón Neártico, que son grupos ecológicamente vinculados a climas fríos o templados fríos y bosques árticos y subárticos o fríos del norte. En la zona de transición mexicana, los insectos paleoamericanos no tienen sus límites necesariamente en las montañas; pueden extenderse a Sudamérica y las Antillas. Antes del Mioceno, estos grupos ocuparon el altiplano, que junto con las montañas sirvieron como centros de diversificación y evolución. Asimismo, estos grupos se extendieron incluso antes de las desertizaciones del oeste de América del Norte, que dieron lugar a formas especializadas.
Como un todo, el patrón paleoamericano incluye 2 tipos de linajes: 1) los ampliamente exitosos (generalmente también dominantes fuera del continente americano, especialmente en áreas tropicales y templadas cálidas). Las especies de estos grupos se encuentran en el este y oeste de los Estados Unidos, mientras que otras siguen en México y América Central y América del Sur, aunque en América del Sur el número de especies disminuye. 2) Formas con distribuciones relictuales altamente localizadas, que generalmente indican nichos ecológicos especializados o áreas geográficas que de alguna manera están protegidas; las especies del mismo género o grupo en el Viejo Mundo pueden tener una distribución euritópica” (Halffter, 1976: 8).
Así, un género cuya distribución abarca América del Sur y del Norte, más rico en especies en América del Norte, especialmente en el Altiplano mexicano, que ocupa una mayor diversidad de ecosistemas, y con una diversidad más pobre y con distribución relictual en América del Sur (generalmente con especies más primitivas), indica un origen que se remonta al Cenozoico temprano, al cual denomina Paleoamericano. En cambio, un género muy diversificado en América del Norte y, particularmente, en las montañas del territorio mexicano continental, con un fuerte sesgo hacia los ambientes templados y fríos, sería evidencia de un patrón septentrional más reciente, al que denomina Neártico. De esa forma, Halffter no solo pudo reconocer patrones septentrionales y meridionales, sino que además pudo distinguir entre patrones antiguos y recientes (Halffter, 1962, 1965, 1967, 1987).
Una vez reconocidos los patrones de distribución, Halffter explicó qué tipo de eventos los produjeron. En sus primeros trabajos, adoptó un enfoque dispersalista. Entre los geólogos, aún no había consenso sobre el fijismo geográfico y la deriva continental. Así que, igual que Miranda (1959), Halffter no toma partido en ese debate y simplemente explica sus patrones por migraciones sobre una geografía estable, adoptando implícitamente las ideas de Darlington (1957): “En cierto número de casos, es muy difícil determinar si los grupos se originaron en el Viejo Mundo o en América. Es posible que algunos de los grupos que consideramos que siguen el patrón paleoamericano se hayan originado en América del Norte, desde donde se han extendido hacia la zona de transición y a Eurasia, a través del estrecho de Behring. Sin embargo, toda evidencia parece sugerir que la mayor parte de los elementos paleoamericanos se han originado en el Viejo Mundo, especialmente en los trópicos” (Halffter, 1976: 7).
Hasta ese momento, lo relevante del método de Halffter es que puede descomponer los patrones imbricados de la zona de transición. Cuando se encuentran varios taxones con los mismos rasgos de distribución geográfica, ecológica y de diversificación, se puede separar como una unidad histórica. Ello permite deconstruir la compleja zona de transición en capas temporales, lo cual permite recuperar patrones biogeográficos históricos independientes (Halffter y Morrone, 2017).
Desde sus primeros trabajos, Halffter afirmó que los eventos geológicos generan diferencias ecológicas, lo que favorece y mantiene la separación faunística (Halffter y Matthews, 1966: 61). Parafraseando a Croizat (1958), se puede decir que tierra, ambiente y biota evolucionan juntos. Por ello, Halffter mencionó, en sus obras posteriores, que los ensambles de especies y todos los aspectos de la biodiversidad son de naturaleza eminentemente histórica (Halffter y Moreno, 2005).
A diferencia de autores holarticistas como William D. Matthew, o neodarwinistas posteriores como George G. Simpson y Philip J. Darlington, Halffter modificó sus explicaciones sobre la naturaleza y origen común de los patrones. Conforme la idea de Wegener de la deriva de los continentes iba evolucionando hacia la moderna teoría de la tectónica de placas, Halffter fue adaptando la explicación de las causas que dieron lugar a los patrones imbricados en la ZTM. A partir de 1978, sus trabajos ya se apoyan en la idea de una geografía dinámica, explicando los patrones de distribución geográfica como resultado de la expansión y diferenciación de linajes por razones geo-históricas, ya sean eventos tectónicos o paleoclimáticos. Esto puede observarse si comparamos los primeros trabajos de Halffter (1962), donde emplea el concepto de “patrones de dispersión”, con sus trabajos posteriores, donde considera que este término no es correcto, ya que dichos patrones no se originan sólo por migración, sino también por eventos de fragmentación de áreas (vicarianza) (Halffter et al., 1995) y lo sustituye por “patrones de distribución”. Estos patrones integran a un grupo de organismos que arribaron desde un área externa, en un periodo geológico dado, y que estuvieron sujetos a las mismas presiones macroecológicas durante un tiempo prolongado.
Una descripción de los diferentes patrones de distribución dentro de la ZTM fue realizada por Halffter y Morrone (2017). En la tabla 2 y la figura 4 se sintetiza la extensión geográfica, diversificación (especiación), amplitud ecológica y la edad de cada patrón, así como sus equivalencias a través de los trabajos publicados por Halffter y sus diferentes colaboradores desde 1962 hasta 2017.
Tabla 2 Patrones de distribución propuestos por Gonzalo Halffter en sus trabajos y su caracterización (con base en Halffter y Morrone, 2017).
| Publicación (Halffter) | Cenocrones (distribución) | Edad | Distribución geográfica y ecológica y diversificación | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1962-1966 | 1976 | 1995-2017 | Patrón mayor | Patrón menor | AM | SMW | SME | FVTM | SMS | NCA | PCM | ANT | AN | ASA | EA | AF | |
| X | X | X | Neotropical | Neotropical típico | Plio-Plei | md | md | hd | |||||||||
| X | X | X | Neártico | Neártico | Mio-Plio | pd | md | md | md | md | md | dga | |||||
| X | X | Mesoamericano de Montaña Altiplano mexicano | Oli-Mio | md | md | md | md | md | pd | pd | |||||||
| X | X | X | L-Cret-Paleo | md, de, dea | de | pd, re, se | ? | ||||||||||
| X | X | Paleoamericano | Tropical | Jur-Cret | pd | pd | pd | pd | pd | pd | ed | dga, dea | dga | ||||
| X | X | Mesoamericano de montaña | dee | dee | dee | dee | dga dea | ||||||||||
| X | X | Paleoamericano de montaña | md | md | md | md | dga dea | dga dea | |||||||||
| X | Paleomericano relictual | ed | ed | ed | ed | ed | dga dea | dga dea | |||||||||
Simbología: Plei (Pleistoceno), Plio (Plioceno), Mio (Mioceno), Oli (Oligoceno), Paleo (Paleoceno), Jur (Jurásico), L-Cret (Cretácico Tardío); AM (Altiplano Mexicano), SMW (Sierra Madre Occidental), SME (Sierra Madre Oriental), FVTM (Faja Volcánica Transmexicana), SMS (Sierra Madre del Sur), NCA (Núcleo Centroamericano), MCP (Planicie Costera Mexicana), ANT (Antillas), AN (América del Norte), AS (América del Sur), EA (Eurasia), AF (África); md (muy diversificado), pd (poco diversificado), dea (distribución ecológica amplia), de (distribución endémica), re (distribución relictual), dee (distribución ecológica estrecha), dga (distribución geográfica amplia).

Figura 4 Configuración de tierras y mares durante la evolución de la Zona Transición Mexicana (a) Jurásico Medio, (b) Cretácico Temprano (Berriasiano), (c) Cretácico Temprano, (d) Cretácico Temprano (Hauteriviano), (e) Cretácico Tardío (Cenomaniano), (f) Cretácico Tardío (Coniaciano), (g) Paleoceno (Selandiano), (h) Eoceno, (i) Oligoceno, (j) Mioceno, (k) Plioceno, (l) Presente
Conclusión
La revisión de las ideas de los naturalistas que han abordado la complejidad de la biota mexicana y la discusión de sus viejas polémicas suelen abrir perspectivas novedosas que ayudan a esclarecer la compleja ZTM. Las ideas biogeográficas de Halffter ilustran una aproximación progresiva a la resolución de la compleja composición de la ZTM, mediante la deconstrucción temporal del modo en que se ensamblaron los distintos taxones. El desarrollo de su obra biogeográfica a lo largo de varias décadas muestra un distanciamiento progresivo del paradigma dispersalista prevaleciente durante la mitad del siglo XX. La originalidad de sus ideas brinda una perspectiva más integral sobre la formación de la Faja Volcánica Transmexicana. Mediante el cotejo de su análisis taxonómico y sus inferencias paleogeográficas y paleoclimáticas, Gonzalo Halffter fue capaz de reconocer que la biota de la ZTM está constituida por patrones de distintos cenocrones.
De acuerdo con Halffter, todas las cordilleras continentales de México son parte de la zona de transición, por lo que no es tan marcada ni tan espectacular como la línea de Wallace. La notable extensión de la ZTM resulta congruente con su explicación sobre la evolución geológica de lo que actualmente es México. Es claro que, hasta el Cretácico, México no existía sino como una península del continente norteamericano. La emersión de la tierra que dio la configuración al actual territorio nacional jugó inicialmente un papel de corredor para el encuentro de las faunas laurásicas y gondwánicas. Solo hasta el Mioceno, pudieron formarse las barreras fisiográficas para la formación de islas virtuales que propiciaron la diferenciación y el origen in situ de especies propiamente mexicanas.
Antes de que se desarrollaran las técnicas de reconstrucción filogenética y filogeografía, Halffter tuvo la capacidad de revelar patrones que hasta ese entonces no habían sido descifrados. A partir de ellos reconstruyó la historia evolutiva de la entomofauna mexicana, con base en el análisis minucioso de la distribución geográfica y ecológica y su diversificación diferencial con respecto a otras regiones. Esto es, mediante una síntesis evolutiva de espacio, tiempo, forma y ambiente. Así, la idea de una zona de transición que resulta de la superposición de varios cenocrones o conjuntos de linajes que comparten un escenario espacial y temporal de evolución persiste hasta hoy. Se entiende así a la Zona de Transición como un conjunto de linajes de historias evolutivas diferentes que ocupan una misma área con diferentes patrones de diferenciación. Esto se debe a que estos cenocrones han ingresado al área en diferentes tiempos geológicos.
Lo más relevante de las aportaciones de Halffter es echar por tierra diferentes postulados implícitos en los análisis anteriores: 1) el predominio de los linajes del norte sobre los del sur; Halffter considera que, en el caso de los insectos, el predominio se da de sur a norte. 2) La reducción temporal de la mezcla de faunas en un momento geológico; de acuerdo con Halffter, la composición actual de la fauna de la ZTM es el resultado de distintos procesos geológicos temporalmente separados. 3) La idea que la ZTM es meramente resultado de la superposición de 2 biotas (neártica y neotropical); para Halffter, existen al menos 5 diferentes cenocrones que se integraron en la ZTM desde el Cretácico hasta el Pleistoceno.
Las ideas de Halffter han sufrido una marcada evolución que va desde una explicación dispersalista hasta una de múltiples historias de fragmentación geológica. Al revisar su obra, resulta claro cómo Halffter incorpora sucesivamente la evidencia geológica y evolutiva que otros especialistas han ido investigando. Las innovaciones en la datación geológica de fósiles en la calibración de filogenias y la inferencia del espacio ocupado por especies en el pasado, así como la integración de todas estas herramientas, siguen teniendo como punto de partida los estudios de la distribución geográfica de la biodiversidad, la discriminación de patrones de distribución, horofaunas y cenocrones.
A lo largo de varias décadas, Halffter elaboró una explicación sobre los patrones biogeográficos de la ZTM que toma en cuenta tanto procesos de dispersión como de vicarianza. En ese sentido, su enfoque coincide con el de autores que han promovido el modelo de dispersión-vicarianza (Brooks, 2004; Crisci y Katinas, 2009; Lieberman, 2004; Morrone, 2003, 2009; Reig, 1981; Riddle et al., 2008; Sanmartín y Ronquist, 2004; Savage, 1982).
El estudio pionero de Halffter (1964) contradijo la tesis preponderante del enfoque holarticista: si bien los vertebrados americanos habían tenido una dispersión norte-sur a fines del Cenozoico, los insectos, de manera sincrónica, habían tenido principalmente una dispersión inversa, es decir, de sur a norte. La dominancia de las formas norteñas no se podía tomar como una premisa básica, al menos no en todos los taxones.
Halffter consideró la diferente temporalidad de diversos procesos paleoecológicos y paleogeográficos para explicar los diferentes patrones que mostraban los insectos. Pudo así distinguir 2 eventos de dispersión, uno fue el de algunos grupos de escarabajos que alcanzaron la actual Altiplanicie Mexicana, pero que ocurrieron desde el Mioceno, antes de la formación de este Altiplano, y otro posterior, que no pudo superar su elevación y se dispersó dicotómicamente por las costas del Pacífico y del golfo de México.
La explicación de Halffter sobre la ZTM desde mediados del siglo XX significó un impulso para el estudio de los patrones biogeográficos de México. Desde entonces, se han realizado una serie de estudios desde diferentes enfoques, que incluyen el dispersalismo, el análisis de trazos, el análisis biogeográfico cladístico, el análisis de parsimonia de endemismos y la filogeografía (Morrone, 2015). La distinción que ha hecho Halffter entre distintos cenocrones coincidentes en la ZTM y su reconocimiento de eventos tanto de dispersión como de vicarianza, ha permitido avizorar de manera más fina la compleja historia de la ZTM y abre la posibilidad de contrastarlos con estudios filogenéticos y biogeográficos que permitan identificar otros cenocrones.

